
Abstract
Tenascin-C (Tn-C) is an extracellular matrix glycoprotein implicated in the progression of several human cancers. In canine mammary carcinomas, accumulation of Tn-C has been recognized in 3 different areas: regions of proliferating myoepithelial cells in complex carcinoma, basement membrane zone in low-grade simple carcinoma, and reactive stroma in high-grade simple carcinoma. To identify the Tn-C synthesizing cells in these areas, we utilized double-labeling immunohistochemistry, branched DNA in situ hybridization, and in situ hybridization–immunohistochemistry double-labeling techniques. In complex carcinomas, Tn-C was generated by proliferating myoepithelial cells. Tn-C in low-grade simple carcinomas was also derived from myoepithelial cells existing as a basal monolayer. However, stromal Tn-C in high-grade carcinomas was mainly synthesized by fibroblasts/myofibroblasts, similar to human breast cancer. Thus, the origin of Tn-C in canine mammary carcinomas differs between low- and high-grade malignancies. The role of myoepithelial cell-generated Tn-C is not yet understood.
Keywords
Tenascin-C (Tn-C) is one of the extracellular matrix glycoproteins that is transiently expressed in organogenesis and reexpressed in pathologic conditions including inflammation and cancer. 3 Tn-C is considered to promote progression of tumors by forming a matrix favorable to neoplastic cell growth, migration, and tumor angiogenesis. 3,6 In several types of human cancers (eg, breast cancer), Tn-C is mainly synthesized by stromal cells, and a high accumulation of Tn-C in cancer stroma correlates with poor prognosis. 3,6 However, in dogs, Tn-C is present in all mammary tumors regardless of their benign or malignant nature. 2,4 Our group and others recognized 3 localization patterns of Tn-C protein in canine mammary carcinomas: areas of proliferating myoepithelial cells of complex carcinoma, basement membrane zone of low-grade simple carcinoma, and reactive stroma of high-grade simple carcinoma. 4,12 These findings suggest that Tn-C in these areas could be synthesized by different cells. As Tn-C has been suggested to have a number of splice variants, 1,3 the Tn-C proteins from different sources in each area may differ in splice variants and play different roles. However, the immunohistochemistry (IHC) performed in the previous studies was not applicable in determining the cellular source of this extracellular matrix protein. 2,4,12
During the last decade, signal amplification strategies for in situ hybridization (ISH) have developed rapidly. Branched DNA (bDNA) technology has recently been adapted to ISH for the detection of DNA or RNA on formalin-fixed paraffin-embedded (FFPE) tissues. 9 Based on signal amplification by a sequential hybridization of multiple probes synthesized by bDNA technology, this ISH method is highly sensitive and specific. 9 In this study, we utilized bDNA ISH to detect Tn-C mRNA in canine mammary carcinomas. Furthermore, to identify the types of cells with Tn-C mRNA, a double-labeling method for IHC and ISH was applied.
Complex carcinoma (n = 3), low-grade simple carcinoma (grade 1 of Peña grading, 8 n = 4), and high-grade simple carcinoma (grade 3 of Peña grading, 8 n = 4) specimens were obtained from the archive of the Department of Veterinary Pathology, Nippon Veterinary and Life Science University. FFPE sections were cut serially and subjected to hematoxylin and eosin staining and IHC with a labeled streptavidin-biotin method with a mouse monoclonal antibody (mAb) against Tn-C (clone 4F10TT, IBL, Takasaki, Japan), α-smooth muscle actin (α-SMA; myofibroblast and myoepithelial cell marker, clone 1A4, DAKO, Glostrup, Denmark), and p63 (basal/myoepithelial cell marker, clone 4A4, Neomarkers, Fremont, CA), as previously described. 12 Additional serial tissue sections were also used for the following experiments.
The protocol of double-enzyme IHC with anti-Tn-C and anti-α-SMA mAbs was as follows. Deparaffinized sections were treated in 0.3% H2O2/methanol for 30 minutes, followed by incubation for 5 minutes at 37°C with 0.1% trypsin. The sections were incubated for 30 minutes with Block Ace (DS Pharma Biomedical Co., Osaka, Japan) and then incubated overnight (4°C) with anti-Tn-C mouse mAb (1:20). After washing, they were incubated with Histofine Simple Stain MAX-PO (NICHIREI, Osaka, Japan) for 30 minutes, followed by visualization in DAB substrate. Next, the slides were heated in a microwave oven in a citrate buffer (pH 6.0) for 10 minutes at 95°C to 100°C to inactivate the anti-Tn-C mouse mAb used in the first staining process. Then, Block Ace treatment was performed again and anti-α-SMA (1:400) mouse mAb applied overnight (4°C). After washing, the sections were incubated with Histofine Simple Stain AP (NICHIREI) for 30 minutes. New fuchsin was used as a chromogen with hematoxylin as a counterstain.
bDNA ISH was performed according to the protocol of QuantiGene ViewRNA for FFPE Samples (Panomics, Fremont, CA). Two dog Tn-C mRNA probe sets (A and B, catalog Nos. VF1-11809-01 and VF1-11500-06, respectively), a dog ubiquitin-C mRNA probe set (catalog No. VF1-11162-06), and an E. coli K12 dapB mRNA probe set (catalog No. VF1-10272-06) were designed by Panomics. The Tn-C probe set A contained 47 types of selected probes covering the 587–1707 region of the mRNA sequence, and the Tn-C probe set B contained 41 types of selected probes covering the 2424–3277 region (Supplemental Table 1, available at http://vet.sagepub.com/supplemental). The tissue sections were refixed in 10% formaldehyde/phosphate-buffered saline, deparaffinized, boiled in Pretreatment Solution (1:100, Panomics) for 5 to 20 minutes at 95°C to 100°C, and digested with Protease (1:100, Panomics) for 10 to 15 minutes at 40°C. Four serial tissue sections were hybridized with the Tn-C probe set A, Tn-C probe set B, ubiquitin-C probe set (positive control), or E. coli K12 dapB probe set (negative control) diluted 1:100 for 4 hours at 40°C. After washing, the sections were hybridized with PreAmplifier Probe (1:100, Panomics) for 25 minutes at 40°C, washed, hybridized with Amplifier Probe (1:100, Panomics) for 15 minutes at 40°C, washed, and finally hybridized with Label Probe conjugated to alkaline phosphatase (1:1000, Panomics) for 15 minutes at 40°C. After washing, the sections were incubated with fast red substrate and counterstained with hematoxylin. Using these hybridization conditions, we found that hybridization signals with the positive control probe set were present in all cells within the sections, while tissue sections with the negative control probe set showed no signal in any cells. Signals with the Tn-C probe set A and signals with the Tn-C probe set B were similar in staining intensity and localization patterns. The specificity of the probes was confirmed in muscular blood vessels as endogenous positive controls and normal mammary glands as negative controls in each slide.
Double labeling with bDNA ISH for Tn-C mRNA and IHC for α-SMA protein was also employed as follows: after being labeled by ISH and visualized by fast red substrate as mentioned above, the tissue sections were incubated with anti-α-SMA mouse mAb (1:400) overnight (4°C). After washing, they were incubated with Alexa Fluor 488 goat anti-mouse IgG (1:500, Molecular Probes, Eugene, OR) for 30 minutes and mounted with a medium containing DAPI (Vector Laboratories, Burlingame, CA). Images were acquired with a Zeiss Axiovert200M fluorescence microscope with an ApoTome system and AxioVision software (Carl Zeiss MicroImaging, Jena, Germany).
In complex carcinomas, extracellular matrices close to proliferating myoepithelial cells showed positive immunoreactivity for Tn-C (Fig. 1a, 1b). In sections with ISH, Tn-C mRNA signals were found in the cytoplasm of myoepithelial cells but not in luminal epithelial cells (Fig. 1c).
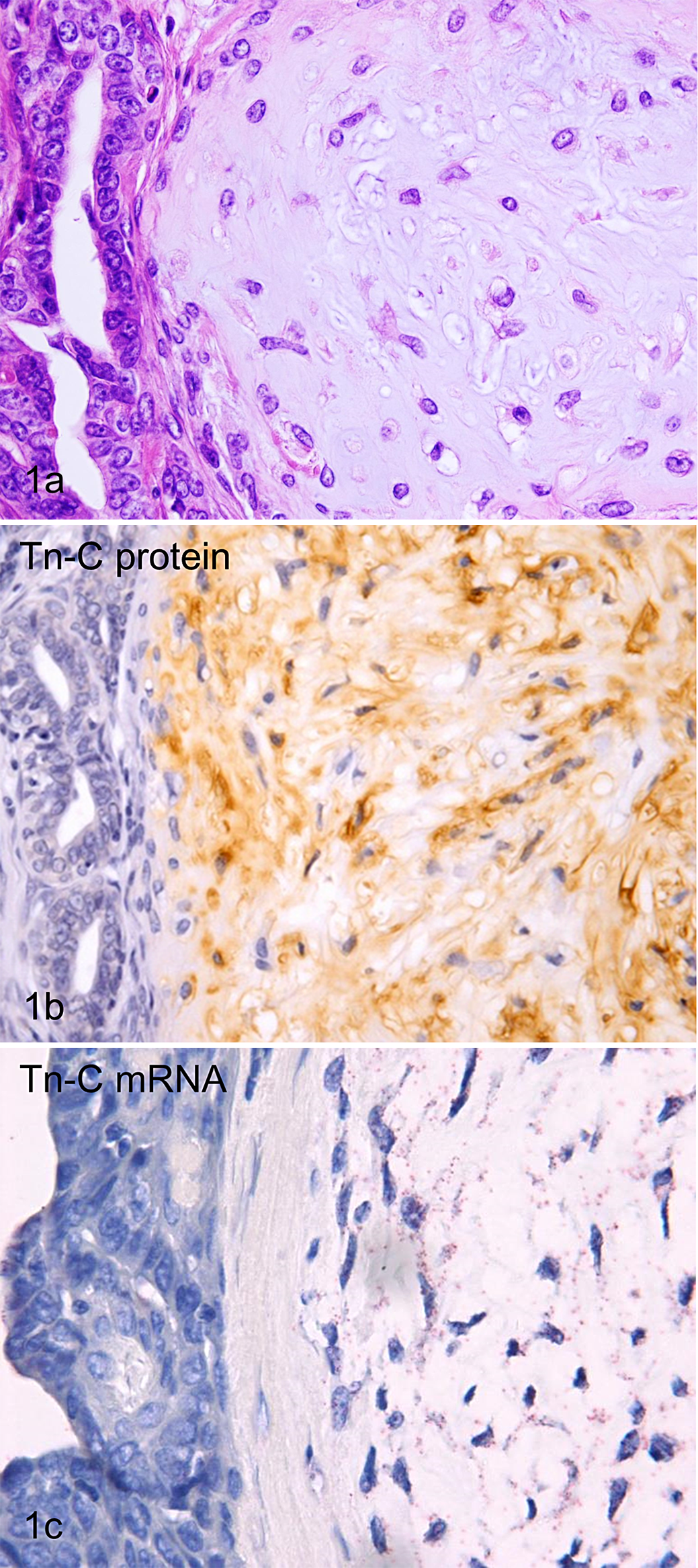
(a) Mammary gland, dog, complex carcinoma. Neoplastic tubules and proliferating myoepithelial cells with abundant extracellular matrices are present. HE. (b) Mammary gland, dog, complex carcinoma. Extracellular matrices around proliferating myoepithelial cells are positive for tenascin-C (Tn-C). Tn-C immunolabeling, DAB chromogen, hematoxylin counterstain. (c) Mammary gland, dog, complex carcinoma. Signal dots representing hybridized branched DNA probes for Tn-C mRNA were localized in proliferating myoepithelial cells. Branched DNA in situ hybridization for Tn-C mRNA (probe set B), fast red chromogen, hematoxylin counterstain.
Low-grade simple carcinomas were composed of tubular and papillary structures of tumor cells (Fig. 2a). Tn-C immunoreactivity was mainly observed in the basement membrane zone adjacent to the tubulopapillary structures. Double-labeling IHC for Tn-C/α-SMA revealed that α-SMA-positive myoepithelium often existed as a discontinuous monolayer beneath the luminal epithelium, and the Tn-C immunoreactivity in the basement membrane zone was almost consistent with the existence of myoepithelium (Fig. 2b). In ISH sections, Tn-C mRNA signals were detected in the cytoplasm of cells located at the basal layer of the tubulopapillary structures (Fig. 2c). However, there were no signals in luminal epithelial tumor cells. Double labeling with ISH and IHC clearly demonstrated that the basal layer cells with Tn-C mRNA signals were α-SMA-positive myoepithelial cells (Fig. 2d).
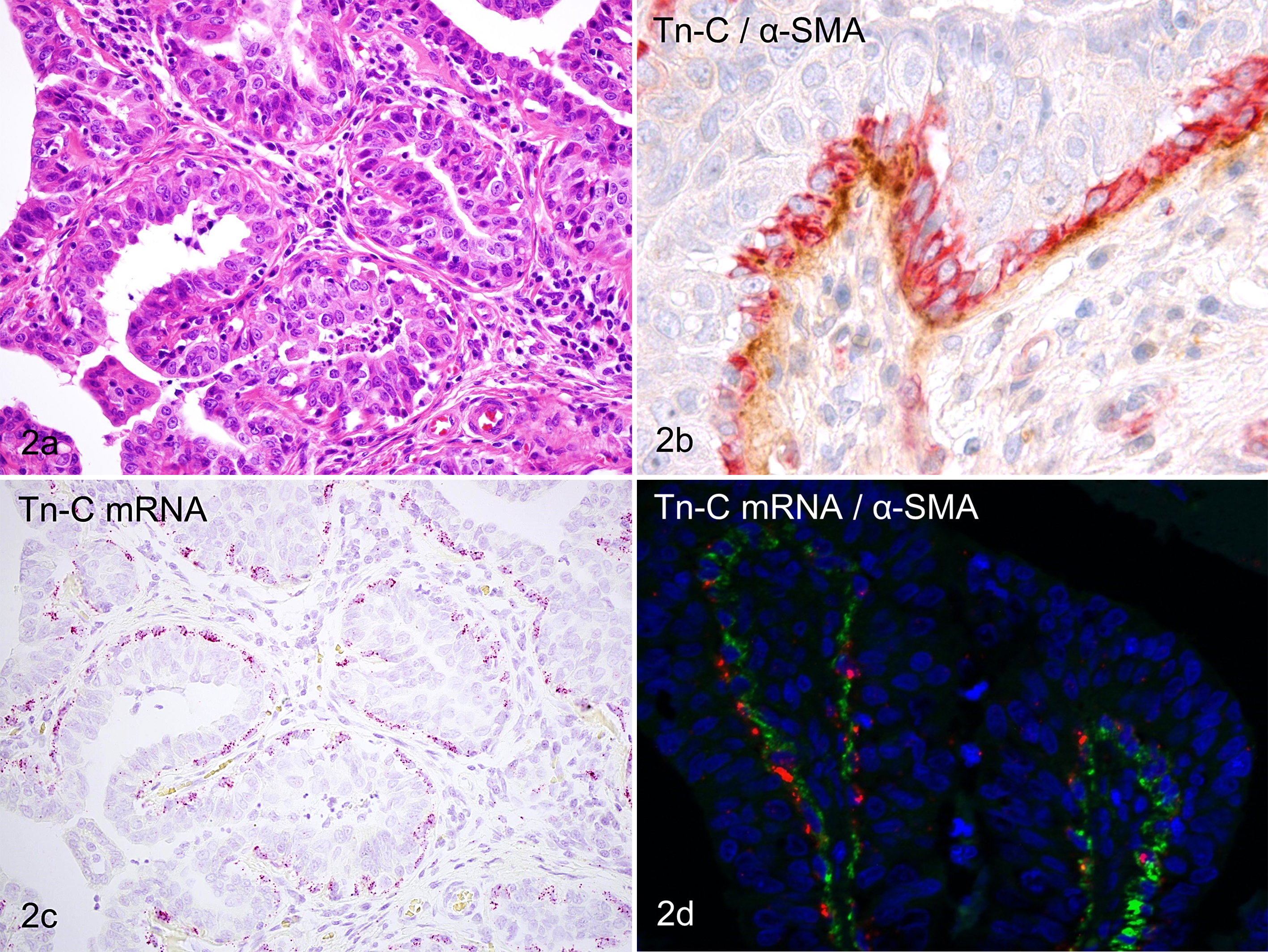
(a) Mammary gland, dog, low-grade simple carcinoma. Luminal epithelial tumor cells with moderate atypia are arranged in tubular and papillary structures. HE. (b) Mammary gland, dog, low-grade simple carcinoma. α-Smooth muscle actin (α-SMA)–positive myoepithelium (red) exists as a single layer at the basal compartment beneath the luminal epithelium. The basement membrane zone adjacent to the myoepithelium is immunolabeled for Tn-C (brown). Tn-C and α-SMA double immunolabeling, DAB and new fuchsin chromogen, hematoxylin counterstain. (c) Mammary gland, dog, low-grade simple carcinoma. Tn-C mRNA signals are localized to basal layer cells but not luminal cells in the tubulopapillary structures. bDNA in situ hybridization for Tn-C mRNA (probe set B), fast red chromogen, hematoxylin counterstain. (d) Mammary gland, dog, low-grade simple carcinoma. Tn-C mRNA signals (red) are present in α-SMA-positive myoepithelium (green). Double labeling with in situ hybridization for Tn-C mRNA (probe set B) and immunohistochemistry for α-SMA protein, fast red chromogen and Alexa 488, DAPI counterstain.
High-grade simple carcinomas were characterized by highly infiltrative growth and prominent reactive stroma with extracellular matrices intensely immunopositive with Tn-C (Fig. 3a). Tubulopapillary structures in these tumors lacked a myoepithelial layer and basement membrane pattern of Tn-C immunoreactivity. Double-labeling IHC for Tn-C/α-SMA revealed that the desmoplastic stroma was rich in α-SMA-positive myofibroblasts and intercellular deposition of Tn-C (Fig. 3b). Signals of Tn-C mRNA were detected in stromal cells but not in epithelial tumor cells (Fig. 3c). In sections double labeled with ISH–IHC, Tn-C mRNA signals were often present in α-SMA-positive myofibroblasts (Fig. 3d) and less frequently detected in α-SMA-negative fibroblasts. A small number of α-SMA-positive myofibroblasts without Tn-C mRNA signals also existed.
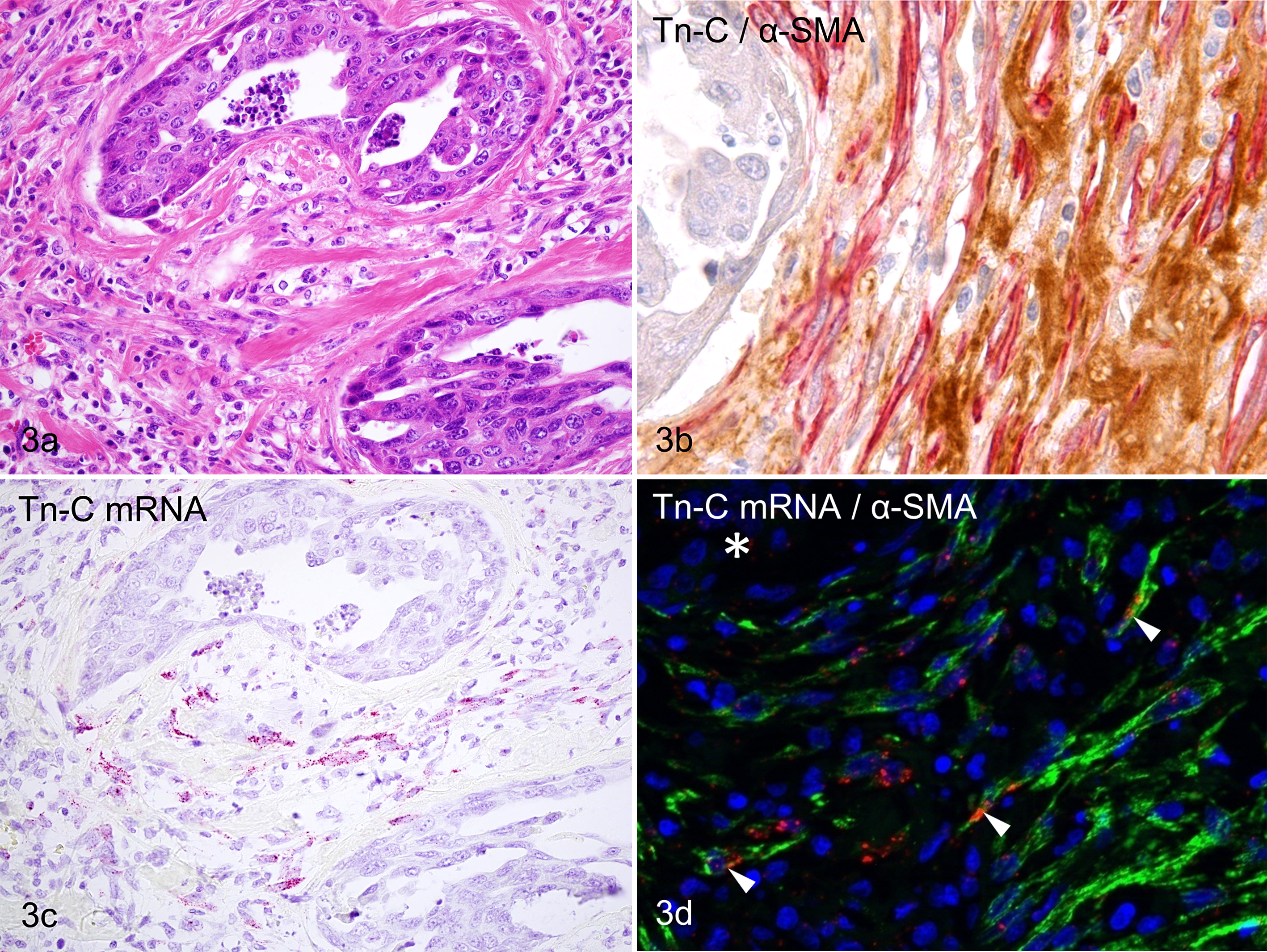
(a) Mammary gland, dog, high-grade simple carcinoma. Marked reactive stroma formation surrounds the tumor nests, which are composed of luminal epithelial cells with a high degree of atypia. HE. (b) Mammary gland, dog, high-grade simple carcinoma. Reactive stroma is rich in α-SMA-positive myofibroblasts (red) and Tn-C-positive extracellular matrices (brown). Tn-C and α-SMA double immunolabeling, DAB and new fuchsin chromogen, hematoxylin counterstain. (c) Mammary gland, dog, high-grade simple carcinoma. Some stromal cells possess Tn-C mRNA signals, but carcinoma cells do not. bDNA in situ hybridization for Tn-C mRNA (probe set B), fast red chromogen, hematoxylin counterstain. (d) Mammary gland, dog, high-grade simple carcinoma. Tn-C mRNA signals (red) are present in some α-SMA-positive myofibroblasts (green, arrowheads). The nest of neoplastic cells does not show any signals (asterisk). Double labeling with in situ hybridization for Tn-C mRNA (probe set B) and immunohistochemistry for α-SMA protein, fast red chromogen and Alexa 488, DAPI counterstain.
To solve the long-standing question regarding the localization pattern of Tn-C in canine mammary tumors, we utilized double-labeling IHC, bDNA ISH, and ISH–IHC double-labeling techniques. As expected, in complex carcinomas with a high accumulation of Tn-C in areas of myoepithelial cell proliferation, this extracellular matrix protein is produced by the myoepithelial cells themselves. In low-grade simple carcinomas, double-labeling IHC revealed that α-SMA-positive myoepithelial layers were present adjacent to the Tn-C immunolabeled-basement membrane zone, and ISH–IHC double labeling clearly indicated that the myoepithelium was associated with Tn-C. Canine simple carcinomas, especially low-grade ones, frequently possess a myoepithelial cell layer adjacent to the basal membrane. 10 In our previous study, 12 there was hardly any basement membrane pattern of Tn-C immunoreactivity in feline mammary carcinomas. The likely reason was that most feline mammary carcinomas lack any myoepithelial layer, unlike canine mammary carcinomas. Tn-C production from myoepithelial cells was also noted in ductal carcinoma in situ of the human breast. 1
Contrary to the relatively low-grade malignancies mentioned above, Tn-C in high-grade simple carcinomas was mainly produced by stromal cells within their reactive stroma. In particular, α-SMA-positive myofibroblasts were identified as the main cellular source of Tn-C, though they were not exclusively nor consistently the only origin. The appearance of stromal myofibroblasts is known to correlate with grades of malignancy in canine mammary tumors. 12 In addition, recent studies in human cancers have suggested the presence of tumor-stromal interaction as follows: secretions such as TGF-β from cancer cells induce transdifferentiation of stromal fibroblasts into myofibroblasts, and secretions such as Tn-C from myofibroblasts modulate cancer cell proliferation and metastasis. 3,5 Recent studies in human breast cancer have also demonstrated that carcinoma cells themselves can express Tn-C, which plays an important role in metastasis 7 ; however, there were no significant Tn-C mRNA signals localized to the luminal epithelial tumor cells in this study.
In conclusion, we found the difference of cellular origin of Tn-C in carcinomas of the canine mammary glands: it was produced by myoepithelial cells of low-grade carcinomas and by stromal fibroblasts/myofibroblasts in high-grade carcinomas. Until now, the Tn-C protein derived from myoepithelial cells has received less attention. Further research is required to clarify the significance and role of Tn-C generated by myoepithelial cells.
Footnotes
Acknowledgement
The authors thank Ms Yoko Kawamoto for her excellent technical assistance. This work was supported in part by a grant-in-aid for fellows of the Japan Society for the Promotion of Science (No. 25-11084 to H.Y.).
This work was supported in part by a grant-in-aid for fellows of the Japan Society for the Promotion of Science (No. 25-11084 to H.Y.).
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported in part by a grant-in-aid for fellows of the Japan Society for the Promotion of Science (No. 25-11084 to H.Y.).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.