
Abstract
Influenza has been recognized as a respiratory disease in swine since its first appearance concurrent with the 1918 “Spanish flu” human pandemic. All influenza viruses of significance in swine are type A, subtype H1N1, H1N2, or H3N2 viruses. Influenza viruses infect epithelial cells lining the surface of the respiratory tract, inducing prominent necrotizing bronchitis and bronchiolitis and variable interstitial pneumonia. Cell death is due to direct virus infection and to insult directed by leukocytes and cytokines of the innate immune system. The most virulent viruses consistently express the following characteristics of infection: (1) higher or more prolonged virus replication, (2) excessive cytokine induction, and (3) replication in the lower respiratory tract. Nearly all the viral proteins contribute to virulence. Pigs are susceptible to infection with both human and avian viruses, which often results in gene reassortment between these viruses and endemic swine viruses. The receptors on the epithelial cells lining the respiratory tract are major determinants of infection by influenza viruses from other hosts. The polymerases, especially PB2, also influence cross-species infection. Methods of diagnosis and characterization of influenza viruses that infect swine have improved over the years, driven both by the availability of new technologies and by the necessity of keeping up with changes in the virus. Testing of oral fluids from pigs for virus and antibody is a recent development that allows efficient sampling of large numbers of animals.
Keywords
The evolution and epidemiology of influenza A virus (IAV) infections in swine have been integrally tied to the segmented nature of the genome of the virus, which facilitates gene reassortment, and to the adaptability of these genes through mutation, which sometimes facilitates replication in a new host. 8,19,77,85,89 Influenza was first recognized as a respiratory disease entity in swine during the 1918 human Spanish flu pandemic. 67 The virus was not isolated until 1931, when nasal discharge from pigs was inoculated into ferrets and subsequently into embryonated chicken eggs. 129 In 1933, a similar virus was recovered from humans with this technique. 131 The viruses were essentially identical and served as the prototype for type A subtype H1N1 influenza viruses.
All influenza viruses of significance that infect swine are type A viruses, defined by the highly conserved internal nucleoproteins and matrix proteins. Influenza A virus subtypes are based on the external hemagglutinin (HA) and neuraminidase (NA) proteins. The predominant viruses in swine populations in different parts of the world may have genes of different origin, even though the subtypes may be similar. 10,73,171 The original classic H1N1 swine influenza virus, which remained the predominant influenza virus in North American swine populations for nearly 70 years, has been almost completely displaced by subsequent reassortant viruses. 157
Pathogenesis of Infection
Although the genetic makeup of influenza viruses in swine varies, the clinical disease and lesions induced by the classic H1N1 swine influenza virus, 52,53,101,139,158,166 avian-like H1N1 viruses adapted to swine, 9,24,29 and various H1N1, 21,84,134,155,158,159 H1N2, 29,56,155,158 H3N2, 76,118 and H2N3 87 reassortants (including the recent 2009 human pandemic H1N1 78,156 and subsequent reassortants 15,63 ) are essentially the same. The gross and microscopic lesions, clinical signs, virus distribution in tissues, and virus shedding patterns have been described with variable emphasis in the above studies and in other reports on isolates of interest, in comparison studies, and in vaccine trials. Two of the earliest studies, one employing histopathology and immunofluorescence, 101 the other histopathology and ultrastructural characterization, 166 still provide some of the most detailed histopathologic descriptions.
In experimentally infected pigs inoculated with a sufficient dose of challenge virus (2 ml of 106 egg infectious dose 50 [EID50] or tissue culture infectious dose 50 [TCID50] intratracheally) and in the individual pig with natural infection, the course of infection is very short (5–7 days) with minimal virus detected after this time. In the immunofluorescent study, 101 the first evidence of virus infection was a pale fluorescence in the nucleus of bronchial epithelial cells by 2 hours postinoculation (PI). By 4 hours PI, virus antigen was abundant in both nucleus and cytoplasm of infected cells. In the ultrastructural study, 166 virus was observed budding from the surface of type II pneumocytes as early as 5 hours PI. In a more recent ultrastructural study on a differentiated human airway epithelial cell line, virions in high numbers were budding from cell surfaces at 24 hours PI, predominantly from the tips of microvilli of both ciliated and secretory cell types. 69 Although virus has been detected in mediastinal lymph nodes, blood, and other tissues such as brain in a few studies in swine, 9,24,77 productive influenza virus replication with consequent tissue damage is limited in this species to the respiratory tract.
Because the virus enters the lung through the airways, the most consistent macroscopic manifestation of influenza virus infection is cranioventral bronchopneumonia that affects a variable percentage of the lung. In milder infections, small clusters of medium to dark red lobules are located in the cranial and middle lung lobes and occasionally in cranioventral portions of the caudal lobes and in accessory lobes (Fig. 1). In some pigs, these foci of consolidation are somewhat linear and confluent and situated in the hilar region at the junction of the lung lobes. In more severe or extensive infections, most or all of the cranial and middle lung lobes and cranioventral portions of the caudal lobes may be consolidated. Less frequently, severe acute infection presents as a diffusely congested lung with prominent interlobular edema and extensive foam in the bronchi and trachea. Such cases likely represent an exaggerated cytokine response, and in such lungs, the characteristic cranioventral lobular consolidation is obscured by the more diffuse interstitial pneumonia and edema.
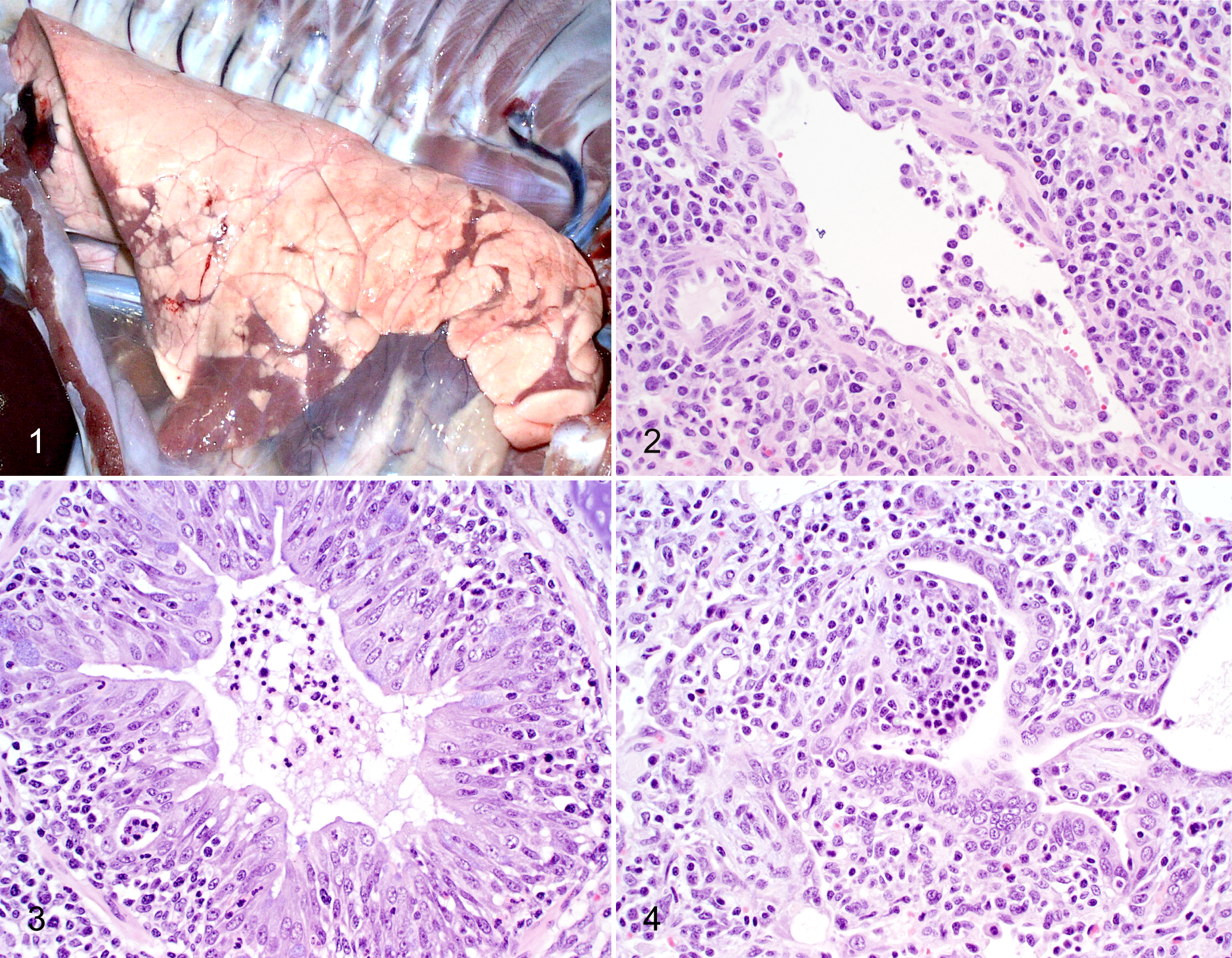
In swine, influenza viruses primarily infect the epithelial cells lining the surface of the respiratory tract, from nasal mucosa to alveoli, but virus also has been detected in the glandular epithelial cells associated with the larger airways. The hallmark microscopic lesion is necrotizing bronchitis and bronchiolitis. During the first 24 hours PI, neutrophils begin to accumulate in the vasculature adjacent to bronchioles, and light loose lymphocytic cuffs appear around the airways, sometimes before epithelial damage is evident. Necrosis and sloughing of airway epithelial cells and neutrophil transmigration into airway lumens occur at 24 to 48 hours PI (Fig. 2). By 48 hours PI, affected airways are lined by flattened epithelial layers, and neutrophils in the luminal debris have been replaced or are accompanied by macrophages. Lymphocyte accumulation around airways and adjacent vessels becomes more prominent. Continuing in repair, the epithelial cells lining the airways proliferate and mature into tightly packed tall columnar cells, resulting in an irregular hyperplastic appearance, often by 72 hours PI (Fig. 3). In severely damaged airways, proliferation of fibroblasts from exposed subepithelial lamina propria may result in formation of endobronchial polyps or incomplete splitting of repairing airways (bronchiolitis obliterans) (Fig. 4). Such damage is most likely to occur in the smaller airways.
In both field cases and experimental challenge studies, the sizes of airways that are affected may vary. In the author’s experience, the largest noncartilaginous airways (primary bronchioles) are most consistently and most severely affected, likely due to the dynamics of airflow that affect droplet suspension and deposition or possibly due to receptor distribution. There is some variation between cases, and the larger lobar or segmental bronchi may be spared or only minimally affected, or conversely, only these larger airways may be damaged. Likewise, the smallest terminal or respiratory bronchioles also are frequently spared but in severe extensive infections may exhibit the characteristic epithelial necrosis. An early immunofluorescent study on sequentially sampled pigs described infection as progressive, beginning in the small bronchi followed by distal spread to terminal bronchioles and alveoli as well as proximally to bronchi and the trachea, 101 which may suggest that some of the variation in lesion distribution is a function of time after infection or of the position of the lobule in question along the infected branch of the respiratory tree.
Simultaneous to the onset and progression of bronchial and bronchiolar damage and inflammation, degenerative and inflammatory lesions develop in the alveoli. 101,166 During the first 24 hours PI, alveolar walls, primarily peribronchiolar in location, are widened by vascular congestion and interstitial edema. Endothelial cells lining the capillaries are swollen, and neutrophils accumulate in capillary lumens and marginate against vessel walls. Type II pneumocytes become swollen. Neutrophils, macrophages, and lymphocytes lightly infiltrate the alveolar interstitium and migrate into alveolar lumens. As in the airways, neutrophils predominate early in infection, but subsequently (by 48 hours PI), the alveolar luminal exudate becomes predominantly mononuclear. These cells appear to be a mixture of sloughed swollen pneumocytes and macrophages. By close examination, necrosis of constituent cells may be identified in alveolar walls, but the septa usually remain intact. In severe infections, necrotic cellular debris accumulates in alveolar lumens, or rarely, the necrotic debris and exudative proteins form hyaline membranes. In such cases, alveolar and interlobular edema is usually evident. Swollen alveolar epithelial cells lining the lumen add to the thickness of the alveolar septa. In some infections, in contrast to the alveolar lesions described above, alveoli may be affected only minimally.
Consistent with the multifocal macroscopic lesions in the lung, lobules within the same histopathologic section of lung may differ in the degree of involvement, and unaffected lobules often sit adjacent to severely affected lobules. This likely reflects irregular distribution of virus through the branches of the respiratory tract via inhalation of aerosolized virus. Experimentally, the virus titer in the inoculum and the route of inoculation can affect lesion development. 24,76 Inducing clinical disease and significant lung lesions can be difficult with intranasal inoculation as pigs may swallow much of the inoculum, reducing the volume of inoculum that reaches the lung in contrast to intratracheal inoculation. If the virus dose is too light, lesions may be minimal but multifocally asynchronous in development. The latter issue can interfere with studies that evaluate lesion progression in sequential necropsies.
Subsequently, the epithelial hyperplasia resolves and all that remain are peribronchiolar lymphocytic cuffing and partially atelectatic alveoli with variable leukocyte populations. Although somewhat dependent on the extent of damage, lungs return to normal by 2 weeks PI. However, in pigs that survive severe infection, airways disrupted by bronchiolitis obliterans may provide evidence of earlier infection.
Mechanisms of Cell Injury
Research into the molecular mechanisms of influenza-induced cell injury has been conducted primarily in vitro in cell lines of nonswine origin with viruses not isolated from swine. Although in vitro studies have their limitations because of the artificial nature of the medium, control of variables is more readily accomplished than in the in vivo studies. The segmented genome of influenza viruses and the ability to isolate the effect of specific proteins through generation of virus with the desired genes by reverse genetics facilitate these studies. Commonalities in mechanisms of infection likely exist regardless of the source of virus or host involved. Thus, in attempting to understand pathogenesis of influenza virus infection at the cellular level and the resulting disease in swine (or in other species), the results of such studies warrant consideration. Findings suggest necrosis, apoptosis, and cytokines of the innate immune response all contribute to the destruction of influenza virus–infected cells.
Necrosis
Much of the damage to the respiratory tract by influenza virus can be attributed to direct viral destruction of infected cells. 55,126 Influenza virus infection induces cytolysis of infected cells through suppression of host cell gene expression and protein synthesis. Viral polymerases bind to host cell polymerase transcripts and cleave host messenger RNAs (mRNAs) (5′ cap removal), which then degrade. Viral NS1 protein inhibits export of host mRNAs from the nucleus, which greatly reduces translation in the cytoplasm. Blockage of host cell protein synthesis, in concert with initiation of viral protein synthesis and virus particle budding from cytoplasmic membranes, results in cytolytic death within 20 to 40 hours of infection. Necrosis is characterized by disruption of cell membranes and swelling of cytoplasm and mitochondria, followed by disintegration of organelles and total cell lysis. Cell death by necrosis stimulates a strong inflammatory response through cytokine induction.
Apoptosis
Cell death induced by influenza virus infection also occurs through apoptosis by both extrinsic and intrinsic pathways. 46 Apoptosis has been most thoroughly studied in established cell lines of both respiratory and nonrespiratory origin 74,75,126 but has also been detected in respiratory tract cells collected following in vivo infection. 98,145 Viral components that have been most strongly implicated in the induction and regulation of apoptosis include nonstructural proteins NS1 and PB1-F2 and the ion channel structural protein M2. 46,74,75 NS1 appears capable of exerting either proapoptotic or antiapoptotic influences, depending on intracellular conditions. 46,74,75 Some studies suggest viral neuraminidase 99,124 and M1 proteins 176 and single- or double-stranded RNA intermediates 3 also influence apoptosis. Inconsistent results between different studies with regard to the induction of necrosis vs apoptosis appear to be related to the cell types studied and the viruses used.
Although apoptosis has been proposed as a host defense mechanism that reduces virus spread through rapid death of virus-infected cells and dampens inflammation through phagocytosis of apoptotic cell debris, recent studies suggest viruses can use the process to their advantage. Viruses may enhance their infectivity by inhibiting caspase activity until sufficient replication of viral proteins has been completed. 46,79,94,168 Contrarily, in a case of Highly Pathogenic Avian Influenza (HPAI) zoonosis, an H5N1 virus isolated from a human fatality induced more cellular apoptosis in vitro than less virulent viruses of other subtypes. 75
Immunopathology
Numerous studies have described the cellular and cytokine response to influenza virus infection as measured in vivo in various animal models and in vitro in cell cultures. 11,14,54,55,59,126,148 Contradictory results from various studies on the interactions of components of the inflammatory response suggest that the severity of tissue damage is determined by the balance between proinflammatory and anti-inflammatory cytokine activity. An in-depth discussion of the complex multitude of molecules involved in the innate immune response to influenza virus infection is beyond the scope of this review, but an excellent review article on immunopathology has recently been published. 23 A brief summary of the mechanisms of destruction of influenza virus–infected cells by the innate immune response is given below.
Type 1 interferon (IFN), predominantly INF-α, is generated within a few hours by infected respiratory epithelial cells to inhibit viral replication and infection of adjacent cells 31 and to initiate proinflammatory cytokine/chemokine responses. 107 Surface proteins expressed by the infected cells serve as signals for their destruction by cells of the innate immune system (killer cells, neutrophils, macrophages) resident in nearby tissues. Neutrophils destroy virus-infected cells through generation of oxidases and engulf debris from cells killed by virus but also may damage tissue by release of other enzymes (eg, elastase, myeloperoxidase). 51,61
The leukocytic cells also effect destruction of virus-infected cells during the early innate immune response through generation of a variety of cytokines, 36,51,61,62,109,116,144 which also recruit additional leukocytes to the site of injury. The primary proinflammatory cytokines are tumor necrosis factor (TNF)–α, interleukin (IL)–1, and IL-6. TNF-α and IL-1 are strong upregulators of leukocyte adhesion molecules on endothelial cells. TNF-α also increases microvascular permeability in the lung. IFN-α, TNF-α, and IL-1, IL-6, and IL-8 can be detected in bronchoalveolar lavage fluid (BALF) of influenza virus-infected pigs, reaching peak levels within 18 to 24 hours PI, followed by a drop to insignificant levels by 48 to 72 hours PI. This wave corresponds to a similar rise and fall in neutrophil numbers in BALF, virus titers in lung, lesions of acute necrotizing bronchiolitis, and clinical illness. By 48 hours, macrophages have replaced neutrophils as the predominant inflammatory cell in the lung. 149
Cytokines released by innate immune cells also have effects that reach beyond the localized cellular environment. TNF-α has other well-described actions in the lung, including increased bronchoconstriction and bronchial hyperreactivity, as well as changes in mucus and surfactant production. IFN-α and IL-6 also have systemic effects such as induction of fever and production of acute phase proteins by the liver. 4,116
Although endothelial cells have not been considered targets of influenza virus infection, recent studies suggest that endothelial cells in the lung are significant modulators of both intracellular and cytokine aspects of the acute response. 34,107,138 These studies have been conducted primarily in mouse tissues and human endothelial cell lines using human viruses considered highly virulent but not necessarily pandemic viruses. Various cytokines and cell surface signals play significant roles in the endothelial damage. Endothelial cells variably exhibit necrosis or apoptosis, but the predominant change is loss of intercellular junctions, which results in vascular leakage. 2 When prominent, this leakage corresponds to extremely high cytokine levels in the tissues (cytokine storm). 162 Although some studies have described receptors for influenza virus on the surface of human endothelial cells 170 and virus infection of avian 28,65 and human endothelial cells, 175 these studies suggest the endothelial cell damage is due to cytokines. Influenza virus infection of endothelial cells in swine lungs has not been documented.
In summary, injury and destruction of the cells lining the respiratory tract by influenza virus is effected by both direct virus infection and by the cytokines of the innate immune response. The most severe infections are characterized by high cytokine levels in the tissues. In humans, therapy directed toward important components of the innate immune response may be as effective as antiviral products in alleviating severe lung damage. 107
Immune-Mediated Enhancement of Disease
In an experimental trial evaluating the effect of an inactivated H1N1 vaccine on subsequent challenge with a heterologous virus, several pigs developed lung lesions dramatically more severe than that induced in nonvaccinated control pigs. The enhanced lesions were produced in a second trial with the same vaccine and challenge virus and in additional trials with different vaccines and challenge viruses. 32,33,64,157
The enhanced lesions were characterized macroscopically by involvement of a greater percentage of lung and microscopically by more severe necrotizing and proliferative bronchiolitis, more prominent peribronchiolar lymphocytic cuffing, and severe lymphohistiocytic interstitial pneumonia. There was more intense lymphocyte infiltration into the epithelial layer and subepithelial lamina propria of the large- to medium-sized airways and suppurative bronchitis and bronchiolitis of greater severity than that observed in the nonvaccinated, challenged pigs. The epithelial lining of the trachea was also more frequently disrupted and disorganized, and the subepithelial inflammation was more intense in the vaccinated pigs.
The antigenic relationship between vaccine virus and challenge virus or other factors that might contribute to this enhanced inflammatory response have not been determined. Such overzealous reactions likely occur in field situations, but the frequency of concurrent multipathogen infections that result in more severe pneumonia makes identification of such cases of immune-augmented disease difficult. The researchers speculate that the potential may exist for similar vaccine-associated disease enhancement in humans. 33
Virulence
In discussions on the pathogenesis of disease induced by influenza virus infections, the question soon arises whether viruses vary in inherent virulence capabilities. The clinical signs associated with natural influenza virus infections in both swine and humans can vary considerably, from mild respiratory illness to severe dyspnea followed shortly by death. Secondary bacterial infection can contribute to more severe pneumonia and higher mortality rates in both species. Rapidly spreading epidemics, sometimes accompanied by higher mortality rates, usually signal introduction of a new virus strain against which the population has little immune protection. However, some influenza viral infections without bacterial augmentation and some epidemics with excessively high morbidity and/or mortality rates suggest the capacity for increased virulence may reside within some viruses. In swine, only very occasional influenza field outbreaks present with high mortality that appears to be effected by virus alone. 86 In humans, viruses responsible for pandemics usually elicit higher morbidity rates (likely reflective of greater transmissibility) and/or greater severity of clinical disease (likely reflective of higher virus replication rates) than that observed with most seasonal influenzas, but overall mortality rates may or may not be higher. Depending on the pandemic, the incidence of infection may be higher in certain age segments of the population.
Pathologic studies on tissues from natural infections reveal some differences. In lungs of pigs submitted from high mortality field cases, diffuse noncollapse and prominent interlobular edema may mask the characteristic cranioventral consolidation. Microscopically, the extent of airway involvement (the number of affected airways, both larger and smaller, within each lobule) and the severity of alveolar inflammation (widespread prominence of pneumocyte swelling, increased numbers of infiltrating cells in alveolar walls and lumens, and abundance of proteinaceous fluid in alveolar lumens) are the most remarkable differences from routine infections. In humans, microscopic lesions induced by the most virulent influenza viruses, the 1918 H1N1 and HPAI H5N1 viruses, have been described in some studies as not differing markedly from nonpandemic infections but with the most significant differences involving the degree of inflammatory changes at the alveolar level. 137 The alveolar inflammation described in these studies resembles that observed in severe swine cases. 86 Others have described the lesions induced by the 1918 and 2009 pandemic H1N1 viruses and the HPAI H5N1 virus as involving both the upper and lower portions of the respiratory tract, whereas a seasonal H3N2 virus infection induced lesions primarily limited to the upper tract. 42 Thus, results of pathologic examination of lungs of both naturally infected swine and humans suggest viruses that induce inflammation in the lower respiratory tract cause the most severe disease.
In vivo challenge studies also have been used in the search for more virulent strains. With swine-origin viruses, comparative challenge studies conducted in pigs have not provided definitive results. Attempts to experimentally reproduce severe disease with isolates from high mortality field cases have not been successful. 86 Pigs challenged with viruses adapted to or endemic in swine usually develop only mild clinical disease and similar lesions of mild to moderate severity. Although some studies describe more extensive lung involvement, comparison of results from different researchers must consider the impact of factors that may vary between studies, such as virus titer in inoculum, method of restraint and route of inoculation, age of pigs, and so on. 24,76,118 In most comparative studies with endemic viruses run concurrently under the same protocols, lesion differences have not tended to be dramatic. However, measurement of quantifiable parameters such as fever, cytokine levels in lung or BALF, and virus titers in nasal secretions, lung tissue, or BALF can supply additional data that often correlate well with gross and microscopic lung scores. Such studies provide further evidence of possible virulence differences between viruses. Some studies suggest that the triple reassortant viruses may be more virulent than the classic H1N1 viruses, 158 especially those reassortants that have been recovered from circumstances in which both pigs and humans were infected. 159
Pigs also appear to be relatively resistant to infection with viruses that cause disease in other host species. Infection of pigs with low-pathogenicity avian influenza (LPAI) viruses or seasonal human influenza viruses usually results in limited viral replication with mild clinical disease and lesions, unless the virus adapts through genetic mutation. 77 Even inoculation with viruses such as the 1918 human pandemic H1N1 virus and HPAI H5N1 that induce severe disease in other mammals (humans, primates, ferrets, and mice) only results in disease of similar or less severity than that induced by endemic swine viruses. 82,164
With viruses associated with human disease, challenge studies seeking to identify or characterize virulent strains have of necessity been conducted in animal models, primarily mice, ferrets, and macaques, and in the in vitro studies in influenza-susceptible cell lines. Mice have been used extensively to study influenza viruses but are not a natural host for these viruses; human viruses do not cause disease in mice unless first adapted through serial passage. 6 Ferrets are a better model for human influenza infections. 5,7,153 Pigs probably would be an even closer model, 59 but the logistics and cost of maintaining swine for experimental studies precludes their use in many research institutions. In mouse and ferret models, clear differences in lesion severity between the HPAI H5N1 or 1918 human pandemic H1N1viruses and the seasonal viruses are evident. The severity of disease induced in ferrets by the 2009 human pandemic H1N1 virus is intermediate. 42,146
In summary from these studies, influenza viruses that are more virulent in humans consistently express 2 characteristics of infection: (1) higher or more prolonged levels of virus replication and (2) induction of a strong proinflammatory response (cytokine storm), a significant result of which is increased permeability of alveolar vasculature. 70,81,113,162 Dysregulation of the early innate immune response appears to be responsible for both of these characteristics. 70
Numerous studies, using both in vivo animal models and in vitro cell culture systems, have investigated the viral proteins as the basis for these differences in virulence. Parameters evaluated in animal studies include severity of clinical disease or lesions and virus titers or cytokine levels resulting from infection. Although evaluation of virulence in the in vitro systems is not technically plausible, increased production of virus or cytokines (if the cell type is capable of such production) is interpreted in these studies to be evidence for possible contribution to virulence. These studies often use viruses generated by reverse genetics to evaluate the effect of gene replacement or gene mutation from a more or less virulent strain on an otherwise identical virus.
Many such studies report findings that run counter to earlier work or reach opposite or different conclusions when initial studies are extended to other viruses, hosts, or experimental protocols. Nearly all viral proteins have been implicated as virulence factors, either individually or in concert with other proteins. 17,88,123 Such studies emphasize the polygenic nature of virulence attributes in influenza viruses, the complexity of the interactions between viral proteins, and the importance of differences between hosts (and tissues within hosts) in the disease process. Several recent articles have summarized current knowledge of how viral proteins affect virulence. 6,70,143 The reported roles for various specific viral proteins in virulence are described below.
Hemagglutinin
The hemagglutinin (HA) is a protein protruding from the surface of the virus that is responsible for attachment of virus to host cells, internalization of virus, and intracellular fusion of viral capsid with endosomal membranes. In avian species, the cleavage site in the HA plays an important role in virulence, which is manifested as systemic infection. The single basic amino acid adjacent to the cleavage site in LPAI viruses is susceptible to proteases that are limited to the gastrointestinal and respiratory tracts. The H5 (and H7) in HPAI viruses has a sequence of multiple basic amino acids that allows cleavage by furin-like proteases that are present in many tissues, thus permitting lethal systemic infection. 135
Influenza viruses that naturally infect mammalian hosts do not have a multibasic cleavage site, and systemic infection is not a normal characteristic of influenza virus infection in mammals. Artificially inserting a multiple basic amino acid sequence by reverse genetics into the HA of mildly virulent viruses does not result in increased virulence or systemic infection in ferrets. 122 However, the HPAI H5N1 virus, which does have a multibasic cleavage site, can be lethal for mice and ferrets in which it induces increased cytokine levels and virus titers and results in systemic infection. 136 This does not occur in pigs, 82 which may reflect a difference in tissue proteases in this species. The HA from the 1918 human pandemic H1N1virus, which does not have a multibasic cleavage site but still appears to be highly cleavable, also confers increased virulence to reassortants as reflected in increased virus titers and inflammation in mice and human airway cells. 66,111 Thus, no consistent association has been established between a sequence of multiple basic amino acids in the HA and virulence of viruses possessing such a sequence for mammalian species.
Neuraminidase
Neuraminidase (NA) is an external protein that prevents aggregation of viral progeny upon release from infected cells. Balanced function of HA and NA results in the most efficient virus replication, and alterations in the NA protein can interfere with this balance, resulting in decreased virulence. In studies of HPAI H5N1 in mice, changes in the length and glycosylation of NA influenced virulence. 93 Some NA proteins inherently appear to contribute to greater virulence. In a study with reassortant 1918 human pandemic virus and contemporary H1N1 viruses, NA from the 1918 human pandemic H1N1 virus contributed to high virus replication rates in both mouse lungs and human airway cells. 111 In studies of reassortants between pandemic 2009 H1N1 and seasonal human viruses inoculated into ferrets, reassortants containing the H3N2 NA were more virulent, whereas other reassortants were attenuated. 123 Some studies suggest the NA protein also contributes to apoptosis. 99,124
Polymerases
The polymerases direct cell processes toward virus replication over cell maintenance functions. Within the virus, a trimeric polymerase gene complex is bound with the nucleoprotein attached to each genome segment. After virus entry into the cell and release from the endosome, the viral ribonucleoproteins and template RNA are transported to the nucleus, where viral mRNA is transcribed by the polymerases (RNA-dependent RNA polymerase). PB1 is the catalyst, and PB2 removes 5′ caps from host pre-mRNA molecules (cap-snatching), assisted by the endonuclease PA, for use as primers for viral mRNA production. These viral mRNAs are moved to the cytoplasm for translation of viral proteins and for additional rounds of transcription. 6,12 It is to the advantage of the virus to maintain the function of cellular mRNA synthesis. The polymerases control the steps of production of viral components, and increased rates of activity could result in increased viral replication. 6
Reverse genetics techniques have been used extensively to study the individual polymerases and to compare the virulence of different gene reassortments 6,17,45,47,84,88,123,133,143 and point mutations within specific polymerases. 12,83,177,178 All 3 polymerases have some effect on virulence in various studies, including PA 47,133,178 and PB1, 133 but PB2 protein seems to most significantly alter pathogenicity. 88,133,177 The amino acid at position 627 of the PB2 protein influences virulence and host preference. A glutamic acid at this site (627E) favors replication in avian hosts, possibly through temperature sensitivity (ie, increased replication rates at the warmer body temperatures of birds). Lysine at this site (627K) favors adaptation to and increased virulence in mammalian hosts, which have cooler temperatures in the upper respiratory tract. 90 Another mutation in PB2 that favors mammalian host infection is D701N (aspartic acid to asparagine at position 701), as manifested by increased virus replication. 30 In larger studies using single and multiple gene reassortants, it became obvious that interaction between the polymerases was as important as the individual polymerases. Compatibility of the polymerases with each other as well as with the other viral proteins appeared to be critical. 17,88,133
Surprisingly, the pandemic 2009 H1N1 virus, which was more virulent than most seasonal influenza viruses for humans and in experimental mammalian hosts, did not have the reputed virulence marker mutations in its PB2 protein. 45 When the molecular mutations in PB2 suggested to be virulence markers for avian viruses were introduced individually into the avian-origin PB2 of the pandemic 2009 H1N1 virus, virulence was not altered. 45 In summary, the polymerases appear to influence virulence through control of protein synthesis processes that favor virus production over cell maintenance functions, but sequence variations between polymerases will likely preclude identification of definitive virulence markers for all viruses.
PB1-F2
The PB1-F2 protein is a nonstructural protein that is expressed during infection as an alternative open reading frame of the PB1 gene. 72,130,151 Within host cells, the protein associates most extensively with mitochondria but has also been detected in both cytoplasm and nucleus. 18,94 The results of multiple in vitro and in vivo studies in various cell lines and experimental animals suggest the protein contributes to virulence through several functions: (1) apoptosis induction, primarily in cells of the innate immune system; (2) suppression of early interferon response by infected cells; (3) increased viral replication rates or delayed viral clearance; and (4) increased inflammation in host tissues. Nearly all of these functions require cooperative interaction of the PB1-F2 with other cellular or virus-induced proteins. 41,80,169
Some effects of the protein involve dampening of the host response to infection. Localization of the protein to mitochondria in macrophages results in mitochondrial membrane depolarization and permeabilization, initiating death of these cells via the intrinsic apoptosis pathway. 18,172 Studies also have demonstrated suppression of interferon induction by PB1-F2 in both epithelial and immune cells in the in vitro and in vivo models. 22,151
Other studies have focused on the effect of the protein on virus multiplication and host tissue response. Co-localization of the protein with PB1 in the nucleus leads to increased polymerase activity and likely consequent increased viral replication. 94 In a mouse model, insertion of PB1-F2 from the 1918 H1N1 pandemic virus into a laboratory-adapted virus resulted in a higher viral production rate per cell and a higher infected cell rate. 130 Contrarily, in another mouse model study, virus replication rates were not increased, but virus clearance was delayed. 173 Studies in mice have associated PB1-F2 with increased cytokine concentrations in BALF, infiltration of alveolar walls and lumens with neutrophils and macrophages, and deposition of fibrin and necrotic cell debris in alveoli. 95
Although the PB1-F2 protein appears to contribute to virulence in many studies, the length (full vs truncated) of the protein and association with virulence are not consistent for all influenza viruses. 44,96,121 The protein is expressed at full length (87–90 aa) by most avian influenza viruses, including H5N1 HPAI viruses, and by the 1918 (H1N1), 1957 (H2N2), and 1968 (H3N2) human pandemic viruses, but in truncated form in many viruses from mammalian hosts, including the pandemic 2009 H1N1 virus. Exceptions to these generalizations abound. 72,143,174 Insertion of genes for the full-length protein into viruses with truncated forms does not necessarily enhance virulence, calling into question the protein’s role as a virulence factor. Similarly in swine, most but not all swine triple reassortant H3N2 viruses express full-length PB1-F2 proteins, but in vitro and in vivo studies have not consistently associated the presence of these proteins with increased virulence. 114,115
NS1
The NS1 protein is not a structural part of the virion and is present only within the cytosol of infected cells. The protein contributes to virulence by interference with the host antiviral response, including IFN-α/β production and expression of other cytokines such as TNF-α and IL-6. 6,132 Both proapoptotic and antiapoptotic activities have been documented for NS1. 46,74,75 Inhibiting apoptosis of cells that produce viral progeny and promoting apoptosis of cells of the innate immune system that attack infected cells would be beneficial for the virus. 152
In conclusion, influenza viruses evaluated under a variety of experimental protocols do appear to express a range of virulence or pathogenic capabilities. Identification of specific virulence determinants has proven to be difficult because of polygenic contributions to pathogenicity. The polymerase proteins effect alterations in virulence through misdirection of host cell protein synthetic processes to viral protein production. Viral proteins NS1 and PB1-F2 contribute to virulence through inhibition in innate immune responses of the host. The roles of HA and NA proteins as virulence attributes in the infection of mammalian species are less clear, but balanced function of these 2 proteins is necessary for efficient virus multiplication. Thus, not all HA-NA pairings that emerge from reassortment will likely result in viruses of equivalent virulence.
Cross-Species Infections
Although only tangentially related to pathogenesis of infection or virulence, infection of swine with viruses originating from other species has been a major factor in the evolution of endemic strains within swine populations. In any discussion of influenza in swine, the question of what to expect from cross-species infection receives consideration.
Wild waterfowl serve as the reservoir of nearly all known subtypes of influenza A viruses (16 HA, 9 NA), and the few subtypes that have successfully adapted to mammalian host species originated from this virus gene pool. 163 Recently, an influenza virus expressing a novel 17thHA was detected in bats. 141 The other genes in this virus also were widely divergent from other type A viruses. Until this discovery, all influenza viruses endemic in mammalian host species, if traced to their source, appeared to be the result of crossover by avian viruses. The genes in influenza viruses that currently circulate in swine populations originated from avian viruses that directly infected pigs or that adapted first to humans before contributing genes to swine viruses. 39,77,87 Domestic avian species are infected by a larger subset of the subtypes that infect wild birds, but routine infection of humans and swine is limited to only a few subtypes. Subtypes H1N1, H3N2, and H1N2 routinely infect pigs. Humans are infected with these same subtypes but historically also were infected with an H2 subtype, the 1957 H2N2 pandemic virus (Asian flu). No infection of swine with an H2 subtype virus had been documented until the report of the one localized H2N3 outbreak in the Midwestern United States in 2006. 87
A point of frequent discussion is whether swine must serve as the intermediate host for infection of humans with avian viruses, particularly the pandemic viruses, a question that goes back to the origin of the classic swine influenza virus and the 1918 Spanish flu pandemic. Early studies that reported experimental infection of pigs with nearly all avian HA subtypes 60 and identified receptors for both avian and human viruses in the trachea of swine led to speculation that pigs likely serve as the “mixing vessel” from which pandemic human influenza viruses arise. 50 However, there is no evidence that pigs were previously infected with the subtypes responsible for the 1918, 1957, and 1968 pandemics but rather that, with the exception of the 1918 virus, the pandemic viruses were reassortants of avian viruses and the preexisting viruses circulating in human populations. Recent receptor studies on the respiratory tracts of humans and swine have revealed very similar distribution patterns, suggesting that avian viruses could just as readily infect the human respiratory tract as the swine respiratory tract, negating the need of the pig as an intermediate host. 142,147
Avian and human viruses do not readily infect swine, 77 but infection of pigs with intact avian and human viruses has been reported. 77,85 The presence of avian- and human-origin genes in the triple reassortants that comprise most of the influenza viruses currently circulating in swine also provides evidence for such infections. 73,157,171 These triple reassortant viruses with the cassette of internal protein genes from human-, avian-, and swine-origin viruses appear to have an increased capability of adding genes from viruses currently circulating in human populations. Since 1998, when the triple reassortants were first identified, the incidence of new reassortant viruses that contain genes from human viruses appearing in swine populations has increased. These new reassortant viruses cause significant problems for the swine industry by becoming endemic, adding to the antigenic diversity of influenza viruses against which pigs need protection. 103
Infection of humans with swine viruses also has been documented, 1,63,71,77,100,127,159,167 including more than 70 cases since 1958, with 41 cases in the United States alone from 1990 to 2009. Viruses that apparently reassorted in pigs and infected humans have been documented. 13 The pandemic 2009 H1N1 virus, the initial source of which is still unknown, is the most recent well-known example. 37 Interestingly, in different locations in the world, this virus first caused widespread clinical disease in the human population and only subsequently was detected in swine populations (in which it promptly reassorted with endemic swine viruses 154 ). This pandemic virus highlights the possibility of “reverse zoonosis” that exists where humans and swine closely interact. 125 Serologic surveys have been conducted that suggest people who work with swine are more frequently infected with swine viruses than people who do not. 40,108 Results of these surveys should be interpreted with caution because of possible partial cross-reactivity in the serologic assays between human and swine viruses of the same subtype. Thus, the incidence of human infection with swine viruses is unknown.
Studies have been conducted to determine the factors that favor movement of intact virus across species and reassortment. 77,87 Two factors influencing cross-species infection have received the most focus in swine studies: (1) the relationship between viral HA and the host cell receptors to which the HA attaches and (2) the polymerase gene complex, the viral proteins responsible for usurping cellular processes for virus replication.
Hemagglutinin and Host Cell Receptors
The presence or absence of specific cell receptors dictates which hosts can be successfully infected by a given virus, and the distribution of those receptors in the respiratory tract may influence extent of infection and thus secondarily the severity of disease. Gene reassortment that results in emergence of a virus with a new combination of genes can occur only in a host species with receptors for the viruses that contributed genes to the new virus.
The molecular structure of the HA selects the type of cells within the host that are infected and thus strongly influences the host range. The HA occurs as a trimer molecule projecting from the surface of the influenza virion with a receptor pocket at the distal end that binds to sialic acids linked in either α2,3 or α2,6 conformation to the terminal galactose of oligosaccharide chains on glycoproteins or glycolipids that project from the host cell surface. 112 Avian viruses attach preferentially to α2,3 sialic acids, whereas mammalian-adapted viruses prefer to attach to α2,6 sialic acids.
The relative distribution of these receptors in avian and mammalian tissues and the binding of viruses recovered from different hosts to these receptors have been studied by a variety of techniques: lectin immunohistochemistry on frozen or formalin-fixed tissue sections, ex vivo biopsy tissues, live cells derived from respiratory tracts or respiratory cell lines, 48,50,92,105,128,140 and fluorescein isothiocyanate–labeled inactivated viruses on sections of formalin-fixed, paraffin-embedded tissues. 26,150 Information derived through use of the various methods has tended to be in agreement, but conflicting or confusing results are sometimes reported, and initial “facts” may actually be more complicated. 91,106 Early studies tried to define the association of receptors with specific cell types (ciliated, nonciliated, goblet, type I pneumocyte, type II pneumocyte), with inconsistent results. Later studies revealed that viruses infect more than one type of cell, that some cells express more than one type of receptor, 48,68,140 and that after virus entry, cells may cease to express receptors. 142 In the in vitro studies, removal of sialic acid residues with neuraminidase did not prevent infection, 38,43,140 and virus infection of cells with few or no recognized receptors has been reported. 106 In 2 studies, virus attachment did not always correlate with efficient virus replication. 20,119 These studies may suggest a coreceptor is necessary for virus entry, or at least caution is warranted against overinterpretation of binding patterns to sialic acids. 35 Studies that include discussion of problems with the use of lectins 48 and some of the discrepancies in these studies 106 have been published.
Results of more recent studies suggest that other factors beyond the type of sialic acid linkage also influence attachment: conformation of the glycan, the structure and length of the oligosaccharide chain underlying the sialic acid, the density of receptors on the surface of the cell, the avidity of the binding between HA and receptor, the balance between HA binding avidity and neuraminidase activity, and temperature sensitivity. 49,161 Mutations at HA amino acids 226, 228, and 190 influence host receptor specificity. 104
Several recent studies have provided excellent detail on receptor distribution in the pig respiratory tract. 102,147,150 Although early studies identified both α2,3 and α2,6 receptors on tracheal epithelial cells, setting the stage for establishing the pig’s reputation as “a mixing vessel,” the more recent studies indicate that very few swine tracheal cells express α2,3 receptors. Receptor distribution in human and swine respiratory tracts is very similar, suggesting that swine are no more likely than humans to be successfully infected by avian viruses and thus are not a necessary intermediate host for avian-to-human infections. The ferret respiratory tract is also very similar to that of human and swine, supporting the use of this species as a laboratory model. Contrarily, receptor distribution in the mouse, which is not a natural host for influenza virus, is quite dissimilar. Very few α2,6 receptors are present at any level of the mouse respiratory tract.
In the pig, epithelial cells lining the nasal passages, trachea, and bronchi almost exclusively express α2,6 receptors; only a very few scattered cells with α2,3 receptors were identified in the upper airways in these studies. At the level of the terminal bronchioles, the number of cells expressing α2,3 receptors begins to increase. At the level of respiratory bronchioles and alveoli, both α2,6 and α2,3 receptors can be identified, although the percentages of each and the types of cells expressing these receptors vary with the study. In summary, α2,6 receptors are present in abundance throughout the porcine respiratory tract, but α2,3 receptors are limited to the small bronchioles and alveoli and are of similar or lesser abundance than α2,6 receptors in this portion of the tract. Interestingly, many cells other than the airway epithelial cells also appear to express sialic acid receptors, including peribronchial mucosal gland epithelial cells, subepithelial connective tissue cells, and endothelial cells. 170
The placement of these receptors in various portions of the respiratory tract (and other tissues) dictates to a large degree the distribution of infection and severity of disease in the individual animal of a given host as well as the likelihood of transmission between individuals of the same species and between species. Thus, a preponderance of receptors for a virus in the smaller bronchioles and alveoli will predispose a host to developing severe disease (pneumonia) rather than bronchitis, but transmission of virus between individuals may be limited. Conversely, if most receptors for a particular virus are in the upper respiratory tract and larger airways, virus shedding and thus transmission between individuals may be facilitated, but the disease induced will be relatively milder (bronchitis).
Polymerases and Other Viral Proteins
After hemagglutinins and receptors, the polymerases have received the most study directed toward defining determinants of host range. These studies have been conducted in the in vitro systems and in various experimental hosts (mice, primates, ferrets) but infrequently in pigs. As with much of the basic research on virulence factors, inconsistent results in studies on host range conducted in different experimental hosts precludes definitive extrapolation of observations to other host species until supported by additional evidence.
The techniques that employ viruses generated by reverse genetics to investigate virulence also have been used to provide information about host range, and results should be interpreted with caution. A gene substitution or mutation in a given virus that results in reduced replication in the same host or in vitro system could be interpreted to demonstrate attenuation of the virulence capability of that virus. However, a similar gene change (in the same virus or in another virus) that results in reduced replication when inoculated into a different host could be due to attenuation of the virulence of the virus or be a reflection of host range, that is, a lesser ability to replicate in the second host, regardless of the gene change. Particularly in the investigation of the effect of a specific gene change in additional viruses or in other hosts, sufficient control groups must be included to support interpretation of results and conclusions. Herein lies the problem with studies seeking to clarify viral virulence factors that affect severity of disease in humans since inoculation of humans with potentially virulent viruses is not morally or ethically justifiable. The use of cell culture systems derived from humans is as close to the actual host as such studies can get even though results may not accurately reflect human infection. Likewise, results of studies on swine viruses that are conducted in mice or other laboratory animals or in nonswine cell culture systems may not reflect what happens in the pig.
In investigation of host range, reduced virus replication in a second host may not necessarily be the result of differences in cell tropism but could involve differing ability to efficiently use the protein synthetic machinery or an incompatibility with other viral proteins. As an example, in a study that compared cell tropism, replication, and cytokine induction by HPAI H5N1, pandemic 2009 H1N1, human seasonal H1N1 and H3N2, LPAI H12N5, and classic swine H1N1 viruses in human lung explants, all viruses replicated only in type II pneumocytes, but only the HPAI H5N1 and seasonal viruses replicated to high titer. The avian viruses (HPAI and LPAI) stimulated high cytokine responses, whereas the seasonal viruses and the pandemic 2009 H1N1 stimulated only weak responses. The classic swine virus replicated poorly and stimulated only a weak cytokine response. 165
Evidence is strongest for a major role of the PB2 polymerase protein in host range. 129 One of the most consistent results of studies indicates a glutamic acid (E) at amino acid 627 favors infectivity of avian species, whereas lysine at this position (627K) favors infectivity of mammalian species. 129 Passage of avian viruses in mammalian hosts will result in selection of E627K mutants. Another potential determinant of host range occurs at amino acid 701 of PB2, with aspartic acid favoring avian hosts and asparagine favoring mammalian hosts. In other studies, most of the viral proteins have been shown to have some influence on host range. 16,58,77,97,104
Diagnosis of Influenza Virus Infections in Swine
Methods of diagnosis of influenza virus infections have improved over the years with change being driven by both the availability of new technologies and the necessity of keeping up with changes in the virus. A recent well-referenced review article describes the various tests used in the diagnosis of influenza virus infection in swine and addresses attributes and rationale behind influenza surveillance systems. 25
Pig Selection
Diagnosis of influenza virus infection in swine is most effectively accomplished by sampling multiple pigs rather than individual animals. This approach may be critical because, within a group of affected pigs at any given point in time, not all pigs will be at the same point in the course of infection. Sampling multiple animals increases the chances of catching some pigs at the peak of virus shedding or with maximal viral titers in lung tissues. Given the short course of infection by this virus, selection of the right pig for individual sampling cannot be emphasized enough. Such pigs have high fevers (105°F) and clear nasal discharge and exhibit lethargy, reluctance to move, and dyspnea or hyperpnea. Although cough is a clinical hallmark of influenza virus infection, pigs at this stage of disease will already have passed the point of highest virus load in the lungs.
Samples of Choice
Although samples to be used for diagnostic efforts may involve collection from individual animals, these samples often are analyzed in pools. Antemortem sampling involves collection of nasal secretions on synthetic swabs and suspension of the fluid in saline or similar physiologic solution. Postmortem samples include collection of bronchial swabs or bronchoalveolar lavage fluid flushed from the intact respiratory tract and fresh and formalin-fixed portions of lung with characteristic lesions.
Histopathology
Although a variety of assays are available, histopathologic examination of lung tissue remains a valuable tool in the diagnosis of influenza infection in swine: (1) the acute lesions are nearly pathognomonic for this virus, and (2) lesions induced by the virus will remain after the virus is gone, although resolving lesions become less definitive. No other commonly encountered respiratory pathogen in swine induces widespread necrotizing bronchitis and bronchiolitis. Even severe purulent bronchopneumonia will not cause similar damage to the airways.
Severe porcine circovirus type 2 (PCV2) infections may result in necrosis and fibrosis of medium to small airways that can cause confusion with influenza infection; however, other features of that virus infection differ. In PCV2-infected lungs, the damage to airways may be “one-sided” and not involve the entire circumference of the cross section, and the fibrosis may extend irregularly into the surrounding interstitium. Peribronchiolar inflammation in PCV2 infection also tends to be more histiocytic than lymphocytic.
Resolving influenza infections have lesions that resemble Mycoplasma hyopneumoniae infection, and additional testing may be required to differentiate these possibilities. Airways are lined by tightly packed, somewhat irregular, hyperplastic columnar epithelial cells and surrounded by prominent lymphocytic cuffs. A subtle difference sometimes evident is early nodular hyperplasia of bronchial-associated lymphoid tissue associated with the medium to larger airways (in addition to the lymphocytic cuffing) in the M. hyopneumoniae–infected lungs, whereas the lymphocytic infiltration is relatively evenly distributed around airways in resolving influenza infections.
Methods to Detect Influenza Virus
The direct or indirect fluorescent antibody test using hyperimmune serum (polyclonal antibody) to detect virus antigen in frozen lung sections was the primary assay employed in veterinary diagnostic laboratories for many years. The staining techniques could also be used on cell culture monolayers used for virus isolation.
Immunohistochemistry applied to sections of formalin-fixed tissue is now used more frequently to detect virus in tissue sections. The preferred antibody is a monoclonal antibody against the conserved type A nucleoprotein because it will detect influenza virus of all subtypes. 160 Virus antigen is visualized in both cytoplasm and nucleus, and in many sections, the nuclear staining predominates. This probably reflects the fact that during viral morphogenesis, nucleoprotein is formed within the nucleus. 55
In situ hybridization methods applied to tissue sections also have been described. 57 This technique is not routinely used in US veterinary diagnostic laboratories.
Commercially available antigen-detection enzyme-linked immunosorbent assay (ELISA) kits developed for diagnosis of human and avian influenza virus infections are used in some laboratories to detect the virus in nasal or bronchial swabs or bronchoalveolar lavage fluids. These tests can be used to detect endemic swine influenza viruses because the antibodies in the assays are directed against conserved type A antigens in matrix proteins or nucleoproteins.
Virus isolation can be conducted in embryonated chicken eggs or in a number of established cell lines. Before the development of polymerase chain reaction (PCR) methodologies, virus growth in allantoic fluids was monitored by HA tests. Currently, PCR techniques are used to detect and characterize influenza viruses isolated in eggs or cell cultures. The isolation procedures are more frequently used for recovery of virus for autogenous vaccine production or for amplification of virus titer to facilitate subtype determination than for routine diagnosis.
PCR techniques are currently used routinely for diagnosis because large numbers of samples can be handled efficiently and because the method is least affected by sample quality. Either in separate assays or as a multiplex PCR, genes for the conserved nucleoproteins or matrix proteins are detected as a screening test followed by assays for the HA and NA subtypes.
Serology
The hemagglutination inhibition (HI) test has served as the standard serologic assay for the detection of antibody against influenza viruses in swine. ELISAs have been developed, some of which are available as commercial kits, to remove the subjectivity of the HI test and to facilitate testing of large numbers of samples. However, because of the variety of reassortant influenza viruses circulating in swine populations and the antigenic variation within subtypes, and because of the widespread use of commercial and autogenous vaccines, diagnosis of influenza infection through the use of serologic assays has become much more difficult. The ultimate result is a much reduced ability to detect antibody against many reassortants.
To circumvent this problem due to antigenic variability, a blocking ELISA has been developed that will detect antibody against the nucleoprotein (NP), a highly conserved internal protein in all type A influenza viruses. If results of the blocking ELISA suggest recent influenza infection, additional HI serologic assays can be conducted with a panel of standard virus strains. An ELISA also has been developed that will detect antibody against the NS1 protein. This protein is not present in intact virions in killed vaccines but is produced only in infected cells. Thus, antibody against NS1 will be stimulated in infected pigs but not in vaccinated pigs, an example of a naturally occurring DIVA (Differentiating Infected from Vaccinated Animals) vaccine.
More Than Detection—Virus Characterization
Because of the propensity of influenza viruses for reassortment, mere detection of influenza virus may not provide sufficient information for full evaluation of the field situation and management decisions. Determination of H and N subtypes is routine in many veterinary diagnostic laboratories, but a subtype description by itself provides little information about the origin of the genes that make up an influenza virus or the possible source of infection. Determining the full or partial nucleic acid sequence of the HA1 portion of the hemagglutinin protein allows preparation of dendograms, which compare the current virus isolate with well-characterized viruses endemic in swine populations, as well as prior isolates from the same operation. If this historic information includes prior isolates from the same site or production unit as well as different segments of the pig flow in the same operation, this information may suggest whether the currently detected infection is due to reappearance of the same virus from that site or elsewhere in the operation, or the introduction of a new virus from the outside. The first possibilities may suggest a need for changes in management and biosecurity measures within the operation. The latter possibility may be even more critical if pigs will now be exposed to an antigenically different virus and additional reassortant viruses appear.
Although methodologies exist for characterization/comparison of all genes, characterization efforts on routine cases usually are limited to the HA gene. An exception would be the development of PCR assays for the M gene that were used for surveillance purposes to identify the potential entrance of the 2009 H1N1 virus into swine populations. 140 Research studies in many veterinary diagnostic laboratories focus on characterization of other genes in field isolates with the objective of identifying new reassortants or reassortment trends.
Oral Fluids
A relatively new approach to diagnosis involves collection of oral fluids from groups or pens of pigs to detect virus shedding in oral secretions, a method particularly valuable in detecting a virus with such a short duration of infection in the individual pig. 27,110,117,120 More pigs can be sampled easily and with less expense, without the need to identify individual pigs early in infection. This method has a very high probability of detecting virus when only 10% to 20% of the pigs in the group are shedding virus, and thus this approach can be used to suggest the cause of current illness. Alternatively, through repeated sampling of a single group of pigs as they move through various growth stages or through sampling of sequential groups of pigs at each stage (or a particular stage), the behavior of a virus within a large operation may be better defined.
Oral fluids are collected by hanging absorbent thick cotton ropes on the sides of pens. As naturally curious creatures, pigs will quickly investigate the new object in their environment by chewing on the rope. As social creatures, pigs will join their penmates in examining the rope. Within a short period, the rope will be soaked with saliva. Videotaping has revealed that a large percentage of the pigs in a pen will take a turn at the rope (S. T. Millman, personal communication, 2012). After approximately 10 to 20 minutes, the rope can be removed and the saliva wrung out into a container. Contaminating feed and fecal debris are removed by centrifugation and the supernatant processed for virus detection.
Virus is detected by PCR techniques, although determination of subtype may be less successful in some samples. Virus isolation has proven to be more difficult. Substances are present in oral fluids that may interfere with detection by PCR, neutralize live viruses, or are toxic to cell lines. Studies are being conducted to remove or counteract these substances and improve the efficiency of assays performed on oral fluids.
Oral fluids are not currently used for influenza serology, but testing will likely be available soon. Research studies indicate that the commercial avian influenza multispecies ELISA (FlockChek; IDEXX Laboratories, Westbrook, ME) will detect antibody against a variety of influenza viruses infecting pigs, including the 2009 pandemic H1N1. 21,110 Antibody can be detected in saliva as early as 5 days PI. Antibody induced by vaccination also can be detected. Because antibody titers in oral secretions are not as high as in serum, some alterations in methodology may be necessary for optimum results.
Swine influenza has been a recognized respiratory disease in pigs for more than 90 years, and over those years, the etiology and pathogenesis of infection have been determined and stepwise facilitated by advances in research technologies that allow us to probe further into the details of infection. The virus has continued to change, most significantly by the introduction of genes from influenza viruses from other hosts, to the extent that “influenza A virus infection of swine” better describes this disease than “swine influenza.” These changes in the virus, as well as significant changes in swine production operations, have compelled veterinary diagnostic laboratories to develop improved and more flexible diagnostic methods.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.