
Abstract
Two Boxer dogs developed progressive ataxia in association with a neoplastic infiltration of the spinal leptomeninges. In the first dog, the leptomeningeal neoplasm encompassed the entire cord and the ventral aspect of the brainstem and extended bilaterally into the piriform lobes. In the second, the neoplasm surrounded the C1–C3 segments of the spinal cord and the brainstem without involvement of the brain or spinal cord parenchyma. In both dogs, the neoplastic cells had variably distinct cell borders, clear to eosinophilic cytoplasm, and a round to ovoid hyperchromatic nucleus. Neoplastic cells were immunopositive for Olig2 and doublecortin in both dogs and for vimentin in one dog but were immunonegative for glial fibrillary acidic protein, S-100, CD34, E-cadherin, cytokeratin, CD3, and CD20. The morphological and immunohistochemical features of the neoplastic cells were consistent with an oligodendrocyte lineage. This hitherto poorly recognized neoplasm in dogs is analogous to human leptomeningeal oligodendrogliomatosis.
Oligodendrogliomas are the third most common primary brain tumor in dogs after meningiomas and astrocytomas. Canine oligodendroglioma tends to develop in the periventricular cerebral white matter as an expansile mass that may extend into ventricular or subarachnoid spaces, but with limited tendency to spread through the cerebrospinal fluid pathways. Oligodendroglioma of the spinal cord is rare in dogs and has growth characteristics like those of oligodendroglioma of the brain. 5 Brachycephalic breeds such as Boxer dogs and Bulldogs are predisposed to develop glial tumors including oligodendrogliomas. 10 Most human oligodendrogliomas have growth patterns like those in the dog, but a rare diffuse spinal leptomeningeal neoplasm has been classified as diffuse leptomeningeal oligodendrogliomatosis (DLO). 6 In DLO, neoplastic cells grow around spinal cord segments or the entire spinal cord in the subarachnoid space and remain extramedullary. This report describes clinical, histological, and immunohistochemical findings of oligodendrogliomatosis in 2 Boxer dogs with this unique leptomeningeal growth pattern.
Case Histories and Necropsy Findings
A 9-year-old intact male Boxer dog (dog No. 1) was presented with a 3-month history of hind limb ataxia. Further clinical investigation was declined by the owner. Dog No. 2, a 14-year-old spayed female Boxer dog, presented with a 7-month history of progressive tetraparesis and ataxia. Magnetic resonance imaging (MRI) of the head and neck revealed a mild T2-weighted hyperintensity in the left piriform lobe that was interpreted as edema. In addition, a circumferential contrast-enhancing lesion surrounded the cervical spinal cord from C1 to C3 segments with moderate compression of the cord. Both dogs were euthanatized due to poor prognosis.
At postmortem examination of dog No. 1, an intradural-extramedullary gray mass encircled the entire length of the spinal cord (Fig. 1). In dog No. 2, the spinal cord had multifocal thickening between the first and third cervical (C1–C3) segments. Slicing the spinal cord after formalin fixation revealed a mass that encircled the spinal cord from C1 through C3 and extended rostrally along the ventral meninges of the brainstem.
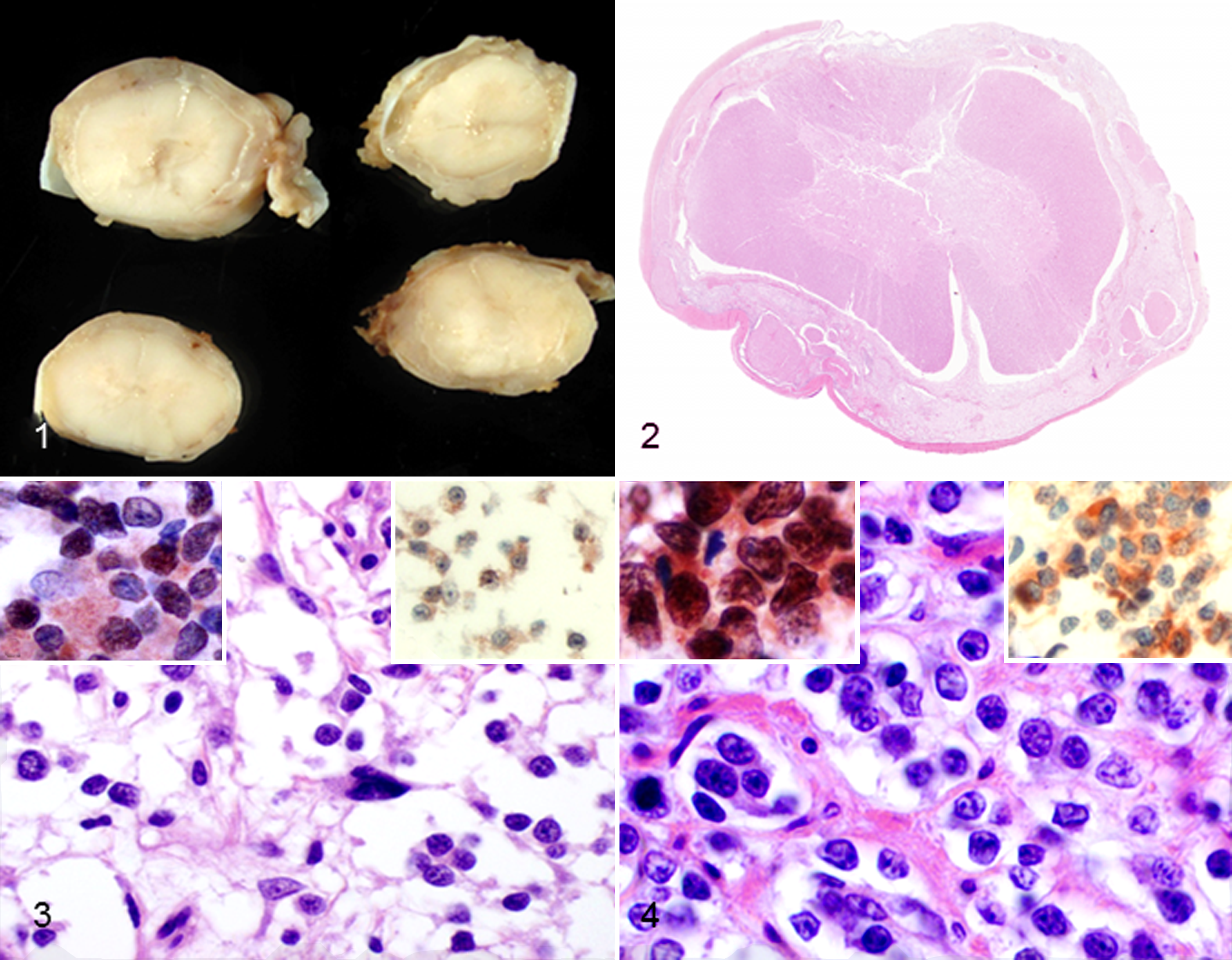
Spinal cord, formalin-fixed cross-sections; dog No. 1. The leptomeninges are diffusely thickened by pale gray to tan neoplastic tissue.
Histopathological Examination
Tissue samples were fixed in 10% neutral buffered formalin, paraffin embedded, sectioned at 5 μm, and stained with hematoxylin and eosin (HE). Selected sections of the mass in both dogs were treated with periodic acid–Schiff (PAS) reaction. A peroxidase-based polymer system (Envision TM-HRP, Dako North America Inc., Carpinteria, CA) was used for immunohistochemistry. Primary antibodies included glial fibrillary acidic protein (GFAP, 1:800, polyclonal, Dako), vimentin (1:500, V9 clone, Dako), S100 (1:800, polyclonal, Dako), cytokeratin (1:25, monoclonal Lu5, Labvision, Fremont, CA), CD20 (1:500, polyclonal, Labvision), CD34 (1:75, polyclonal, Santa Cruz Biotech Inc., Santa Cruz, CA), E-cadherin (1:200, monoclonal, BD Transduction Lab, Lexington, KY), Olig2 (1:250, polyclonal, Millipore, Billerica, MA), and doublecortin (1:800, polyclonal, Santa Cruz Biotech Inc.). Four-micrometer-thick sections were mounted on glass slides, deparaffinized, and rehydrated in a graded alcohol series. The antigen was unmasked using pressure cooker heat-induced epitope retrieval with Diva Decloaker Solution in decloaking chamber (Biocare, Concord, CA) at 125°C for 30 seconds. The sections were incubated with 3% H2O2 for 15 minutes to block endogenous peroxidase activity. Sections were rinsed in Tris-buffered saline with 0.05% Tween 20 (TBST 0.05 M, pH 7.6). To block unspecific binding sites, the sections were incubated with Background Punisher (Biocare) for 10 minutes. After excess blocking solution was drained, the rabbit anti-Olig2 (dilution 1:300; Millipore) polyclonal antiserum was applied for 45 minutes at room temperature. The sections were rinsed with TBST and then incubated with the secondary antibody for 45 minutes at room temperature (EnVision+/ horseradish peroxidase–labeled goat anti-rabbit immunoglobulin G). After rinsing in TBST, positive antigen–antibody reactions were visualized by incubating the slides with 3-amino-9-ethylcarbazole for 15 minutes; sections were counterstained with Mayer’s hematoxylin counterstain. Controls included substitution of the primary antiserum with diluted normal serum from nonimmunized rabbits and inclusion of brain tissue with oligodendrocytes.
In dog No. 1, the spinal subarachnoid space was expanded by round to polygonal neoplastic cells loosely arranged in sheets and packets and separated by clear spaces and fibrovascular stroma (Figs. 2, 3). The neoplastic cells had variably distinct cell borders, moderate amounts of finely granular eosinophilic cytoplasm, and a round to ovoid hyperchromatic nucleus with 1:1 nucleus to cytoplasm ratio. Bilaterally, the piriform lobes of the brain were effaced and moderately expanded by a poorly demarcated neoplasm with similar histological features. Mitotic figures were not detected in any of the brain or spinal cord sections. Immunohistochemically, the neoplastic cells had moderate nuclear reactivity for Olig2 (Fig. 3, left inset) and moderate cytoplasmic reactivity for doublecortin (Fig. 3, right inset). Individual GFAP-positive cells throughout the neoplasm were interpreted as reactive astrocytes. The neoplastic cells were immunonegative for vimentin, cytokeratin, S100, CD34, E-cadherin, CD3, CD20, and cytokeratin.
In dog No. 2, the mass surrounding the spinal cord was composed of neoplastic round to polygonal cells like those in dog No. 1. The neoplastic cells were organized into nests and clusters and separated by a moderate amount of fibrovascular stroma (Fig. 4). The fibrovascular stroma was especially dense in the periphery of the mass subjacent to the dura mater. The neoplastic cells had mild anisocytosis and anisokaryosis, but mitotic figures were not detected. The neoplastic cells were PAS-negative. Neoplastic cells from dog No. 2 had strong nuclear immunopositivity for Olig2 (Fig. 4, left inset) and strong cytoplasmic immunopositivity for doublecortin (Fig. 4, right inset) and vimentin. Rare GFAP-positive cells within the mass were interpreted as reactive astrocytes. The neoplastic cells were immunonegative for CD34 and E-cadherin; however, the stromal cells and nonneoplastic leptomeningeal cells were multifocally, moderately immunopositive for CD34 and E-cadherin. The neoplastic cells were negative for S100, CD3, CD20, and cytokeratin.
Summary and Conclusions
This report describes a leptomeningeal neoplasm in the spinal cord of 2 Boxer dogs. The tumor was intradural but extramedullary in both dogs. Histological and immunohistochemical characteristics of the neoplastic cells are consistent with an oligodendroglial origin. The few GFAP-positive cells within the neoplasm were interpreted as reactive astrocytes, which can occur in the leptomeninges (eg, due to tumor-induced activation of the residential meningeo-glial network). 1 Olig2 is a basic helix-loop-helix transcription factor involved in the specification and initial phase of differentiation of cells in the oligodendroglial lineage. 7 Expression of Olig2 was restricted to glial tumors and nontumoral oligodendrocytes. The proportion of Olig2-positive cells was significantly higher in oligodendroglioma compared with other glial tumors including astrocytoma and oligoastrocytoma. 7 Doublecortin is a microtubule-associated protein that is expressed in migrating neurons during development and is highly expressed in invasive brain tumors in humans. 2 Ide et al 3,4 reported the overexpression of doublecortin in canine oligodendroglioma and anaplastic meningiomas. 3 Neoplastic cells in both dogs of this report expressed doublecortin and Olig2, suggesting that the neoplastic cells had a low degree of differentiation and were of oligodendroglial origin. The neoplastic cells were positive for vimentin in one dog. Although uncommon, immunoreactivity for vimentin has been reported in a canine brain oligodendroglioma. 8 The lack of immunoreactivity for CD3 and CD20 helped to eliminate meningeal lymphoma from the differential diagnosis. The lack of PAS reactivity in the neoplastic cells and their lack of immunohistochemical expression of CD34 and E-cadherin helped rule out meningioma. Canine meningiomas of various histological patterns have been reported to be immunopositive for CD34 and E-cadherin in 92% and 79% of dogs, respectively. 9
The growth pattern of the oligodendroglial tumor in these 2 dogs is unusual. However, widespread dissemination of oligodendroglial tumors into the leptomeninges is recognized in humans. 6 The leptomeningeal infiltration can occur without evidence of an intraparenchymal neoplasm (primary DLO) or can be associated with a parenchymal oligodendroglioma. In dog No. 2, a neoplastic process was not detected in the parenchyma of the brain or spinal cord despite thorough diagnostic imaging (MRI) and macroscopic and microscopic postmortem examination. Therefore, dog No. 2 is consistent with primary DLO. In dog No. 1, the brain involvement was not obvious at necropsy and was recognized only at histological examination. The involvement of the left and right piriform lobes and lack of mass effect are in stark contrast to the usual growth pattern of brain oligodendrogliomas and could indicate that the brain involvement was secondary rather than primary. It has been hypothesized that widespread dissemination of a primary oligodendroglioma within the central nervous system occurs by circulation of exfoliated neoplastic cells in the cerebrospinal fluid within the ventricular system, central canal, and subarachnoid spaces and rarely by direct invasion of leptomeninges. Canalicular spreading of central nervous system tumors along the central canal or leptomeninges may be underreported in domestic mammals because the spinal cord may only be examined when clinical evidence of spinal cord disease is reported to the pathologist. Alternatively, and rarely, glial tumors may arise from heterotopic nests of glial tissue in the subarachnoid space. 1
In conclusion, these 2 cases illustrate a growth pattern of oligodendroglial neoplasia that is poorly recognized in the veterinary literature. Meningeal oligodendrogliomatosis should be considered in the differential diagnosis for segmental or diffuse circumferential leptomeningeal masses in the spinal cord of dogs.
Footnotes
Acknowledgements
Portions of this study were presented at the 2010 annual meeting of the American College of Veterinary Pathologists.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.