
Abstract
β-Catenin, encoded by the ctnnb1 gene, plays a critical role in intercellular adhesion, and its altered expression has been implicated in tumor progression in humans and animals. The aims of this study were to examine the alterations in β-catenin expression in canine melanoma as well as the causes of these changes (eg, E-cadherin or exon 3 mutations) and to compare identified changes between skin and oral melanomas. Forty-two primary canine skin and oral melanoma tissue samples were used in the study. The expression levels of ctnnb1 and the levels of E-cadherin/β-catenin complex in the tissues were determined by semiquantitative RT-PCR and immunohistochemistry, respectively. The mutational status of β-catenin exon 3 was examined by DNA sequencing. RT-PCR revealed higher levels of ctnnb1 expression in oral melanoma tissues compared with normal melanocytes, irrespective of sex or histopathological appearance of the tissue (ie, amelanotic vs melanotic). Immunohistochemistry revealed simultaneous loss of membrane E-cadherin/β-catenin complex and cytoplasmic accumulation of both proteins in 37 cases (84%). Intranuclear β-catenin was also detected in all tissues with reduced membrane β-catenin expression. In mutational analyses, one amelanotic oral melanoma showed 13 single nucleotide polymorphisms (SNPs); however, after protein translation, all the SNPs were silent mutations. The present study demonstrates that dysregulation of E-cadherin/β-catenin complexes is involved in both types of canine melanotic tumors and that the disruption of E-cadherin/β-catenin complexes and increased β-catenin may induce tumor progression and malignancy.
Melanotic tumors originate from the transformation of melanocytes and represent a common neoplastic disease. 9 The transformed melanocytes often lose normal contact with the surrounding keratinocytes and proliferate into the surrounding tissues. 25 In dogs, melanotic tumors account for 9%–20% of skin tumors, and they are the most common malignant tumors of the oral cavity and digits of dogs.3,17,27 In the oral cavity, these tumors primarily arise from the gingiva, but they also come from the lingual, buccal, pharyngeal, tonsillar, and palatine epithelium. 25 The tumors usually grow rapidly and are invasive, often recurring after surgical removal. 25 Metastasis is common (∼70%–90%), irrespective of the location of the tumor in the oral cavity, and usually carries a grave prognosis.4,5,25,27 Unlike oral or digit melanoma, however, melanotic tumors arising from hairy skin are usually benign and surgically curable. 25 The pathogenesis of each type of tumor is largely unknown.
β-Catenin, which is encoded by the ctnnb1 gene, has 2 functions: regulation of cell-to-cell adhesion and involvement in Wnt/β-catenin signaling. 19 In the Wnt/β-catenin signaling pathway, β-catenin acts as an activator of the T-cell factor (Tcf)/lymphoid enhancer factor (Lef) family of transcription factors in the nucleus, resulting in increased cell division and proliferation. In the absence of Wnt signals, the cellular concentration of free β-catenin is kept low through phosphorylation and subsequent degradation by a protein complex of glycogen synthase kinase (GSK)-3β, adenomatous polyposis coli (APC), and axin. GSK-3β recognizes and phosphorylates specific serine/threonine residues in exon 3 of β-catenin, leading to β-catenin ubiquitination and degradation.1,18,23 In the absence of the signals, β-catenin becomes a component in cell-to-cell adhesion through cadherin–catenin complexes. E-cadherin is known as a cadherin type of the epidermis in dogs. Therefore, in the absence of Wnt signal, an abnormal increase of β-catenin in the cytoplasm is associated with dysregulated degradation (eg, dysfunction of the GSK-3β/APC/axin complex or failure of the complex to recognize β-catenin), increased inflow of β-catenin (eg, release from disruption of adherens junction), or increased β-catenin synthesis (eg, from enhanced Wnt signal through receptor tyrosine kinases).2,6,7,18,23
In our previous study, we showed that dysregulated β-catenin expression is involved in canine skin melanotic tumors.10,11 We demonstrated a substantially increased expression of ctnnb1 mRNA by RT-PCR and the migration of unbound β-catenin into the cytoplasm (in both benign and malignant tumors) or the nucleus (in malignant tumors only).
The aims of this study were (1) to examine the dysregulation of β-catenin in canine oral melanoma, (2) to examine correlations of E-cadherin/β-catenin complex levels with the mutational status of β-catenin exon 3, and (3) to compare any correlation between skin and oral melanotic tumors. We propose that the dysregulation of β-catenin also occurs in canine oral melanoma.
Materials and Methods
Samples
Archived tissue blocks of canine skin and oral melanotic tumors referred to the Veterinary Teaching Hospitals of South Korea and the Department of Veterinary Pathology of North Carolina State University were selected by a pathology record search from 2008 to 2010. The tumor tissues had been collected at the time of surgical removal or necropsy and histologically diagnosed as melanotic tumors. A designation of benign or malignant was assigned to each neoplasm based on the following physical and histological features: tumor location and size, ulceration (especially for cutaneous melanotic tumors), nuclear atypia, mitotic index (total number of mitoses in 10 hpf), degree of pigmentation, vascular invasiveness, and intralesional necrosis.24,26 From the database, the following information was collected: breed, sex, age, diagnosis, tumor location, tumor size (diameter), and mitotic index.
Detection of ctnnb1 Expression in Canine Oral Melanomas
Because overexpression of ctnnb1 in skin melanotic tumors was demonstrated in our previous study, we further examined its expression in oral melanomas. Total RNA extraction and cDNA synthesis were performed as described previously. 10 Briefly, total RNA was extracted from normal melanocytes (n = 7) and oral melanoma tissues after resecting the melanocytes or the area of the tumor by using a laser capture microdissection (LCM) system (MDS Analytical Technologies, Toronto, Canada). For normal dog skin, we made 3 continuous skin sections. The first section was stained with hematoxylin and eosin (HE) stain, and the third section was stained with DOPA stain and then restained with HE stain. By examining the first and third section, we confirmed the location of melanocytes, and then we resected melanocytes from the second section.
Characterization of E-Cadherin/β-Catenin Expression
To identify the preservation of membrane E-cadherin/β-catenin complexes in the tumors, cellular localization of E-cadherin and β-catenin was examined by immunohistochemistry (IHC) as described previously. 10 For skin melanotic tumors, the characterization of β-catenin had already been performed in our previous study; here, we additionally evaluated E-cadherin expression in the tumors. The skin tissues of normal Beagle dogs (n = 3) were used as controls. For E-cadherin, a mouse monoclonal E-cadherin antibody (610181; BD Transduction Laboratories, BD Biosciences, San Jose, California) was used at a dilution of 1:100. For heavily pigmented tumors, we additionally made a bleached section to distinguish between the melanin pigment and the chromogen signal, and we confirmed that the labeling pattern of IHC we examined is due to the altered target protein, not melanin pigment.
The immunoreactivities of E-cadherin and β-catenin were scored according to their respective patterns of expression. If the immunoreactivity was localized to the cell membrane and appeared stronger than that in the cytoplasm, it was scored as “membranous.” However, if the immunoreactivity was uniformly distributed throughout the cytoplasm, with no distinction between the cell membrane and the cytoplasm, it was scored as “cytoplasmic.” Next, for both proteins, the cases were scored as “preserved” when there was positive membrane labeling in more than 75% of tumor cells and as “reduced” in all the remaining samples, including negative immunoreactivity cases. In addition, E-cadherin/β-catenin complexes were scored as “preserved” when both proteins had membranous expression and as “reduced” when the expression of at least one protein was low. Last, the immunoreactivity of nuclear β-catenin was recorded as “positive” when the immunolabeling appeared throughout the nucleus. The mean number of tumor cells with nuclear β-catenin positivity per hpf was counted.
Determination of β-Catenin Exon 3 Mutational Status
To determine whether mutations in exon 3 of β-catenin are involved in the pathogenesis of canine melanotic tumors showing cytoplasmic labeling of β-catenin, genomic DNA was isolated from formalin-fixed, paraffin-embedded tissues. The formalin-fixed, paraffin-embedded tissue sections (4 μm) were deparaffinized in xylene and washed with 100% ethanol at room temperature for 5 minutes. After drying the sample for 30 minutes at room temperature, we added 200 μl of tissue lysis buffer containing 100 mM Tris-Cl (pH 8.0), 200 mM NaCl, 5 mM EDTA, and 100 μg/ml proteinase K, and the mixtures were incubated at 56°C until complete lysis. After 1 minutes of centrifugation at 12,000 g, the upper aqueous layer was transferred to a clean tube containing an equal volume of isopropanol at room temperature. After 2 minutes of centrifugation at 16,000 g, the aqueous layer was discarded, and the pellet was washed with 70% ethanol at room temperature. The resulting DNA pellet was air-dried for 5 minutes and resuspended in 30 μl of distilled water. The rehydrated DNA pellet was incubated at 60°C for 1 hour. The concentration and quality of extracted DNA were determined by measuring the absorbance at 260 and 280 nm.
To amplify exon 3, two primer pairs were designed to cover the intron/exon junctions around exon 3 by using the sequence of canine ctnnb1 available from GenBank databases (accession number NM_001137652.1) (Table 1). Each primer pair was designed to contain a 128-bp-long overlapping sequence with each other. The conditions of the designed primers were verified by DNAMAN software, version 4.16 (Lynnon Corp, Pointe-Claire, Quebec, Canada). The template DNA (5 μl) was amplified by PCR in a reaction volume of 50 μl containing 10 mM Tris-HCl (pH 8.5), 50 mM KCl, 2 mM MgCl2, dNTPs (0.2 mM each), and 0.5 units of Taq polymerase (Thermo Fisher Scientific, Waltham, Massachusetts). PCR amplifications were performed as follows: 1 cycle at 94°C for 5 minutes; 35 cycles at 94°C for 30 seconds, 60°C for 30 seconds for annealing, and then 72°C for 30 seconds; and a final cycle at 72°C for 3 minutes. The sizes of the resultant PCR products were analyzed by 2% agarose gel electrophoresis.
Oligonucleotide Sequences for Cloning β-Catenin Exon 3.

All PCR products were sequenced using an ABI PRISM Bigdye Terminator Cycle Sequencing Ready Reaction Kit, version 5.1 (PE Applied Biosystems, Foster City, California). Finally, all identified sequences were merged (with the overlapped sequences excluded) and compared with the normal canine sequence (NM_001137652.1) by using CLC Sequence Viewer, version 4.6.2 (CLC Bio, Aarhus, Denmark).
Statistical Analysis
Associations of ctnnb1 expression levels among normal melanocytes, oral melanoma, and skin melanotic tumors were analyzed by Kruskal-Wallis test, and then the expression levels between normal melanocytes and oral melanomas or between skin and oral melanotic tumors were reanalyzed by Student’s t-test. The interaction between E-cadherin and β-catenin was graded as yes (preserved)/no (reduced) and was tested for significance by using the Fisher’s exact test. The relationships between E-cadherin, β-catenin, E-cadherin/β-catenin, and mitotic index were investigated separately by Student’s t-test. All statistical analyses were performed using SPSS for Macintosh edition (IBM Corp., Armonk, New York). P < .05 was considered statistically significant.
Results
Canine Melanotic Tumor Cases Selected for Evaluation
Forty-two canine melanotic tumors were used in this study. Eighteen were skin melanotic tumors (16 melanocytomas and 2 melanomas), and 24, oral melanomas (Fig. 2). If a benign or malignant designation could not reliably be applied to a particular neoplasm, that neoplasm was excluded from the study. The number and breed of dogs from which the tumor tissues were collected included 6 Labrador Retrievers, 4 Pekingese, 3 Golden Retrievers, 3 American Cocker Spaniels, 2 Akitas, 2 mongrels, 1 Newfoundland, 1 English Shepherd, 1 Staffordshire Bull Terrier, and 19 dogs with unknown breed. The skin melanotic tumors were from 2 neutered males, 9 intact males, and 7 intact females with a median age of 9.4 years (range, 2–16 years). The oral melanoma tissues were from 8 neutered males, 2 intact males, and 4 female spayed dogs with a median age of 9.5 years (range, 2–13 years). Four cases of oral melanoma were amelanotic melanomas. Two oral melanoma cases, 1 amelanotic and 1 melanotic melanoma, revealed metastasis to the lung/brain/lymph node and lymph node/tonsil, respectively. The level of mitotic index of the oral melanomas (n = 24) was 25.50 ± 17.67 (range, 6–85/10 hpf). Follow-up survival data could not be collected because of either a change in the patient identification number or an incomplete medical record.
Characterization of ctnnb1 and β-Catenin Expression
When normalized to GAPDH expression levels, normal melanocytes, oral melanoma, and skin melanotic tumor tissues showed significantly different levels of ctnnb1 mRNA expression (Kruskal-Wallis test, P < .0001) (Fig. 1). All the oral melanoma tissues examined exhibited significantly higher levels of expression (mean ± SD of ctnnb1/GAPDH, 1.07 ± 0.36) than normal melanocytes (0.51 ± 0.18) (Student’s t-test, P < .0001); however, they showed much lower levels of expression than the skin melanotic tumors (2.95 ± 1.17, data from the previous study), although most of the skin melanotic tumors were benign (16/18, 88.9%) (Student’s t-test, P < .0001). The expression levels did not differ between males and females or between amelanotic and melanotic melanomas.
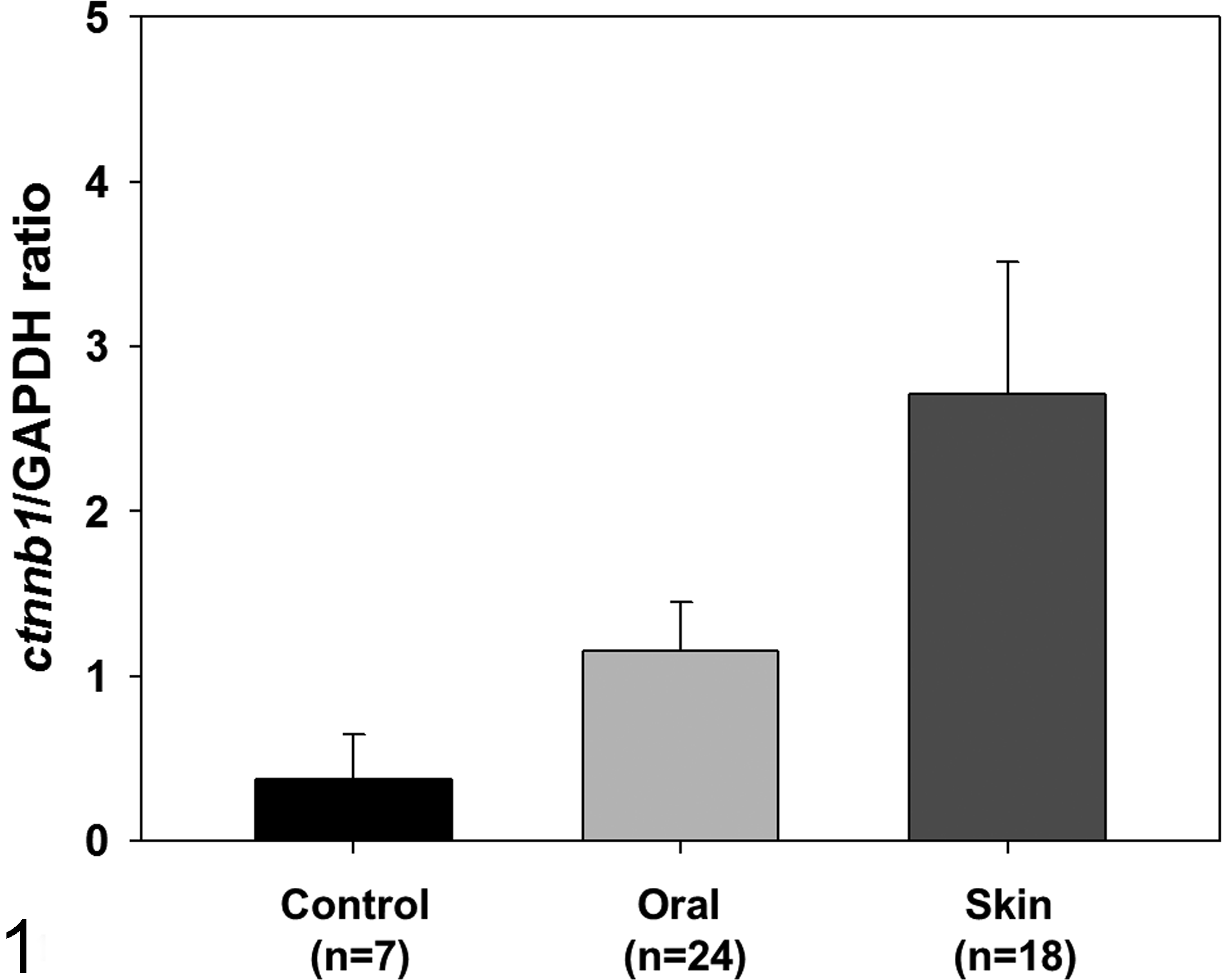
Statistical analysis of the expression levels of ctnnb1 in normal melanocytes (n = 7), oral melanoma tissues (n = 24), and skin melanotic tumor tissues (n = 18) indicating higher levels of expression in both oral and skin melanotic tumor tissues (Kruskal-Wallis test, P < .0001). The skin melanotic tumor tissues revealed higher expression of ctnnb1 than did those of oral melanoma. The bar graphs summarize the experimental analysis (mean ± SD). Control, normal melanocytes; oral, oral melanoma; skin, skin melanotic tumors.
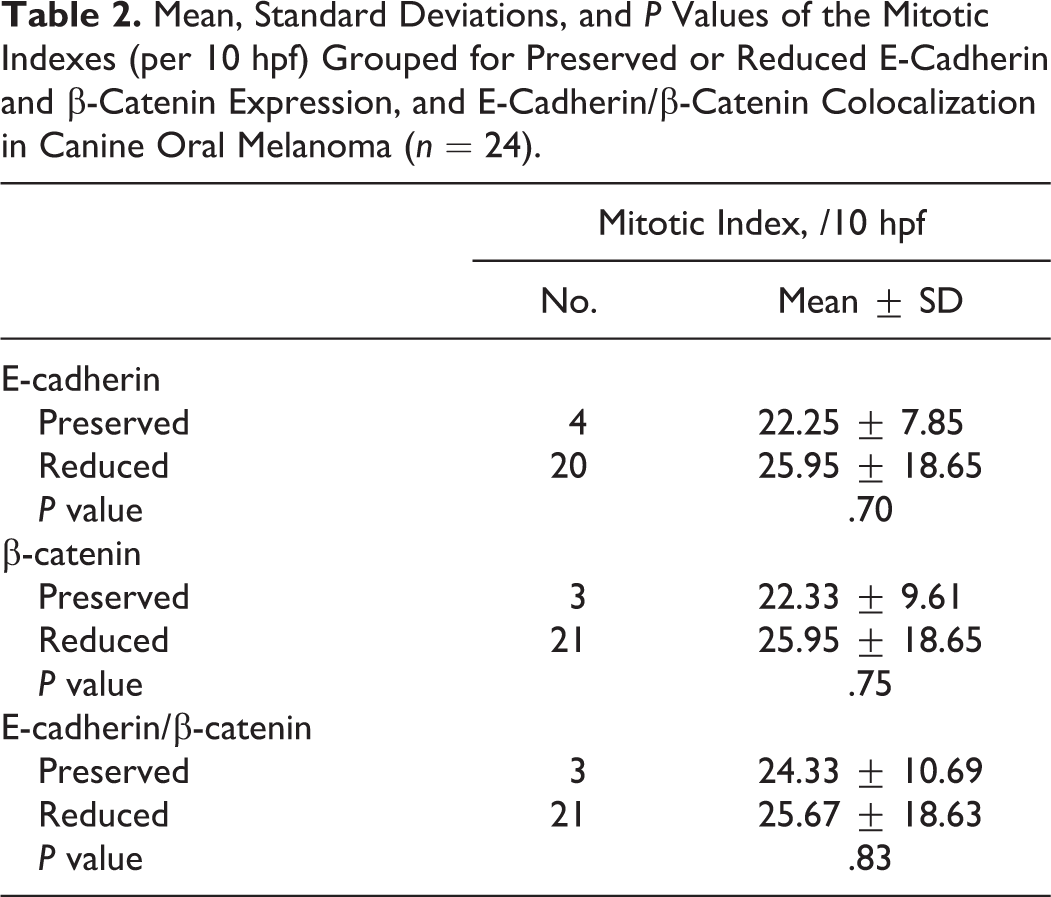
For IHC, immunolabeling for both E-cadherin and β-catenin was classified as membranous, cytoplasmic, or negative, and only for β-catenin, nuclear positive. The membrane labeling patterns of E-cadherin and β-catenin appeared as red labeling, which was stronger in the membrane than in the cytoplasm (Fig. 3). The cytoplasmic labeling of both proteins appeared stronger than membranous labeling, or no distinction between membrane and cytoplasm was noted (Fig. 4). In 4 of 24 oral melanoma cases (17%), E-cadherin expression was preserved, whereas in the remaining 20 (83%), E-cadherin expression was reduced (20 reduced and 0 negative). For β-catenin in oral melanomas, 21 (87%) cases had reduced expression (21 with cytoplasmic labeling and 0, negative). In skin melanomas, E-cadherin expression was preserved in 5 of 18 cases (28%). For β-catenin, we had confirmed in the previous study that all cases showed reduced expression. 10 According to our IHC analysis, expression of the 2 proteins in the same tumors was statistically associated (data for skin melanotic tumors from the previous study) (Fisher’s exact test, P = 0.001). Concordance was identified in 38 cases; in 33 cases, both proteins were reduced, and in 5, both were preserved. The expression levels in the remaining 4 cases did not correlate (all had reduced β-catenin and preserved E-cadherin expression). E-cadherin/β-catenin complexes were reduced in 37 of 42 cases and preserved in 5 of 42 cases. In all of the oral melanoma cases with reduced membrane β-catenin expression (n = 21), nuclear localization for β-catenin was examined as done for the skin melanomas (n = 2) (Fig. 5). 10 The mean number of tumor cells with nuclear β-catenin positivity per hpf was 2.81 ± 1.40 (range, 1–7 per hpf). The oral melanoma cases without the reduction of membrane β-catenin expression did not show the nuclear positivity of β-catenin. No significant differences were obtained by comparing the mitotic indexes (n = 24) between the melanoma cases with preserved or reduced E-cadherin, β-catenin, and E-cadherin/β-catenin complexes (Table 2) or by examining the association between the expression level of ctnnb1 mRNA and nuclear localization of β-catenin.
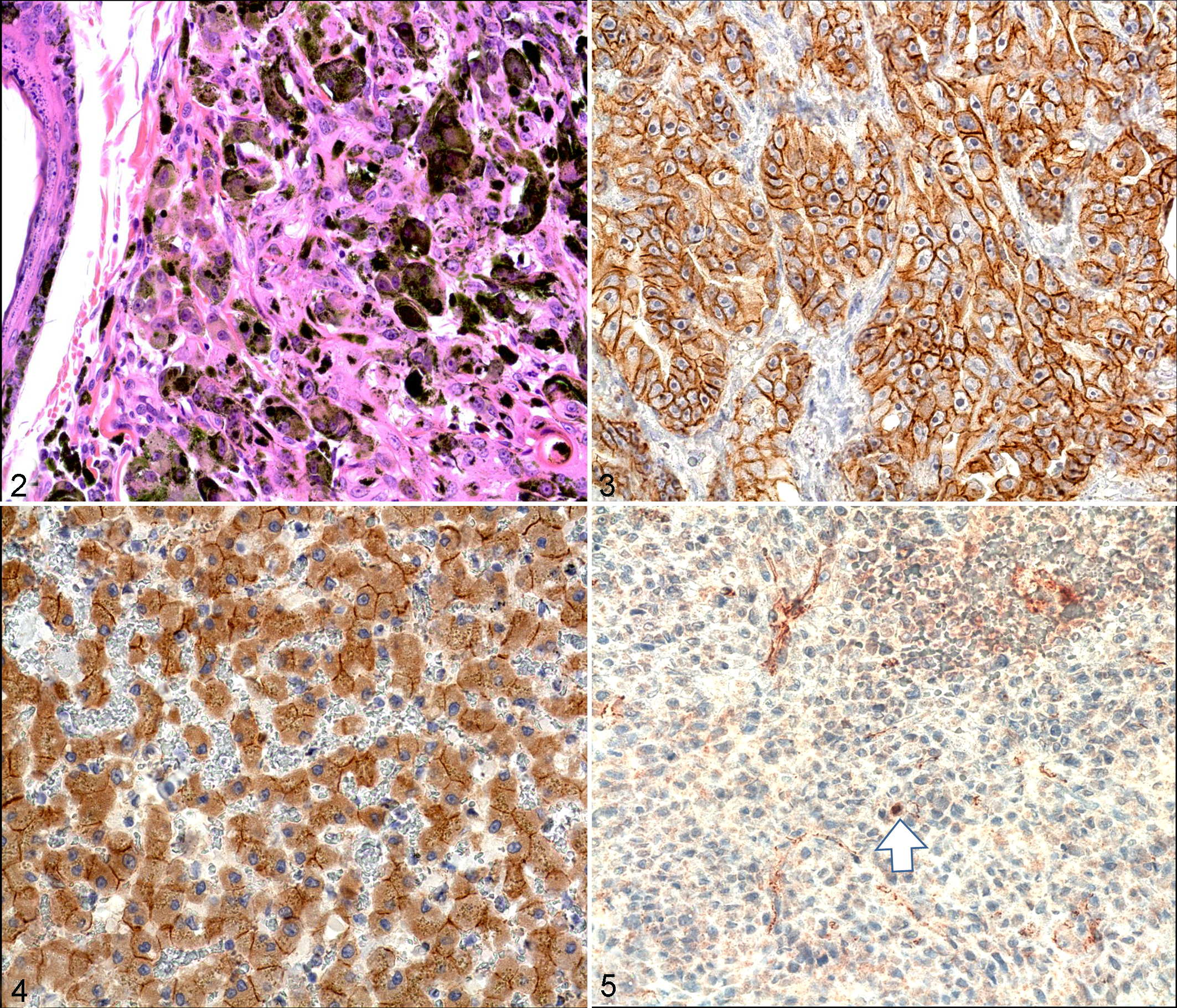
Skin melanoma; dog No. 113. The lesion of the melanoma showing nests, packets, and bundles of neoplastic melanocytes within the dermis. HE.
Mean, Standard Deviations, and P Values of the Mitotic Indexes (per 10 hpf) Grouped for Preserved or Reduced E-Cadherin and β-Catenin Expression, and E-Cadherin/β-Catenin Colocalization in Canine Oral Melanoma (n = 24).
Mutational Analysis of β-Catenin Exon 3
DNA sequencing was performed to determine the frequency of mutation in β-catenin exon 3 in 34 cases that showed cytoplasmic labeling of β-catenin. β-Catenin exon 3 was successfully amplified and sequenced from 29 of the cases we collected (11 melanocytomas and 18 melanomas). When compared with the wild-type sequence of canine β-catenin exon 3, only 1 oral melanoma revealed 13 single nucleotide polymorphisms (SNPs) in the DNA sequence (T8C, G16A, T55C, T85C, T109C, T118C, C127G, C145T, A154G, T181C, C196G, A205G, and A226G). In the translated sequence, however, there were no amino acid changes, indicating that the SNPs did not affect the amino acid sequence of β-catenin exon 3.
Discussion
Like our previous study of canine skin melanotic tumors, this study also demonstrated that dysregulation of β-catenin is present in canine oral melanoma. We detected a consistent increase in ctnnb1 expression in all the cases examined and altered localization of β-catenin protein in 21 of 24 cases (87%). Although increased RNA synthesis does not always equate with increased protein synthesis, the increased expression of ctnnb1 mRNA suggests the possibility that increased β-catenin synthesis may affect the intracellular accumulation of β-catenin protein. Along with the expression pattern of ctnnb1 mRNA, we examined the changes in membrane E-cadherin in skin and oral melanomas and confirmed that the release of both proteins from the cell membrane seems to occur in parallel, irrespective of whether the tumor was benign or malignant. In particular, all skin (n = 2) and oral melanoma (n = 21) samples with reduced membrane β-catenin protein also showed nuclear labeling for the protein. The results of IHC suggest the possibility that the disruption of E-cadherin/β-catenin complexes releases β-catenin into the cell and that this β-catenin enters to the nucleus, resulting in cell division/proliferation through the activation of Tcf/Lef family transcription factors.
The search for β-catenin exon 3 mutations that can induce recognition failure by GSK-3β showed no changes in the amino acid sequence of β-catenin protein, indicating that degradation failure by GSK-3β does not affect the intracellular accumulation of the protein in canine melanotic tumors. In cell line–based studies of human melanomas, exon 3 mutations have been found to be widespread, but studies using surgically extracted skin melanotic tumor tissues have indicated that the mutations rarely occur in clinical cases.8,19 Tumor cells with exon 3 mutations seem to be better at adapting to cell culture conditions, but the exact reason for this behavior is unclear. It also suggests that the results of cell line–based studies, especially for melanomas, should be interpreted with caution. This is why the present study was carried out with surgically extracted tumor tissues.
In a comparison of ctnnb1 mRNA expression levels between skin melanotic tumors and oral melanomas, the skin tumors showed much higher levels of expression than the oral cases, although most of the skin melanotic tumors were benign (16/18, 88.9%). Considering the role of β-catenin in cell division/proliferation, these results suggest that increased synthesis of β-catenin protein may be involved in tumor development; however, this is not a crucial factor in the progression to malignancy in canine melanoma. Rather, it seems that its intranuclear translocation is a crucial factor in determining whether the tumor will proceed to malignancy. To confirm this hypothesis, a future study aimed at correcting abnormal Wnt/β-catenin signaling will focus on blocking the nuclear translocation of β-catenin in a canine melanocyte cell line.
Besides the releasing of β-catenin into the cell cytoplasm, the loss of membrane E-cadherin is also involved in the reduction of intercellular adhesion, in parallel to an invasive growth behavior in human melanoma.12,13 The mechanism of down-regulation of E-cadherin in human melanoma is not fully understood; however, its down-regulation is known to be involved in some processes that potentially promote melanoma development.14,20,22 Results from our present study also demonstrated that significant loss of E-cadherin is a frequent event in canine melanotic tumors regardless of tumor location and its invasiveness; however, its exact mechanism and role in tumorigenesis need to be investigated.
Although extensive research and therapeutic trials are ongoing, the incidence of human melanoma in the Caucasian population has been increasing for several decades. 21 Alterations in Wnt/β-catenin signaling have been found in 30% of human cases. 16 In these cases, increased β-catenin translocation into the nucleus is thought to mediate fundamental roles in the radial and vertical growth of tumor mass, crossing the basement membrane.15,23 Therefore, a mouse model has been developed to study Wnt/β-catenin signaling in vivo. However, this model has limitations. For example, mice have a different location of melanocytes, compared with humans, and no incidence of spontaneous melanoma. 15 In contrast, spontaneous canine melanoma has several similarities to human melanoma, beside the dysregulated β-catenin confirmed by our studies, that make the system advantageous: (1) similar location of melanocytes (epidermis and hair follicle), (2) similar age at incidence of disease onset, and (3) shared histopathological and biological characteristics, especially for short radial growth and rapid vertical growth of the tumor.10,11,25 Although the most common place for canine melanoma is on their gums or on the skin between the nails and toes, meaning that UV exposure is not a crucial factor for canine malanomagenesis, 19 these advantages suggest that spontaneous canine melanoma can be a suitable model for therapeutic trials for human melanomas with altered β-catenin localization.
Footnotes
Acknowledgements
The authors thank staff members of Veterinary Teaching Hospital of Chungbuk and Seoul National University in Korea and North Carolina and Iowa State University in the United States for their help in collecting samples.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This work was supported by the Basic Science Research Program through the National Research Foundation of Korea (KRF) funded by the Ministry of Education, Science and Technology (2010-0024447).