
Abstract
In humans, oral SCCs are either caused by papillomavirus (PV) infection or by other carcinogens such as tobacco. As these 2 groups of SCCs have different causes they also have different clinical behaviors. Immunostaining using anti-p16CDKN2A protein (p16) antibodies is used to indicate a PV etiology in human oral SCCs and p16-positive SCCs have a more favorable prognosis. The present study investigated whether p16 immunostaining within feline nasal planum SCCs was similarly associated with the presence of PV DNA and with a longer survival time. Intense p16 immunostaining was visible in 32 of 51 (63%) SCCs. In 30 cats with nonexcised SCCs, cats with p16-positive neoplasms had a longer estimated mean survival time (643 days) than cats with p16-negative SCCs (217 days, P = .013). Papillomavirus DNA was amplified more frequently from p16-positive nasal planum SCCs (28 of 32) than p16-negative SCCs (5 of 19, P < .001). The different survival times in cats with p16-positive and p16-negative SCCs suggests that p16 could be a useful prognostic indicator in these common feline cancers. As the clinical behavior of the SCCs can be subdivided using p16 immunostaining, the 2 groups of SCCs may be caused by different factors, supporting a PV etiology in a proportion of feline nasal planum SCCs.
Keywords
In humans, papillomaviruses (PVs) cause approximately 25% of oral SCCs while 75% are caused by other carcinogens such as tobacco and alcohol. 1 The different causes of the SCCs result in 2 genetically distinct groups of SCCs which have different clinical behaviors. 1 Immunostaining to detect p16CDKN2A protein (p16) is used to determine a PV etiology of a human oral SCC. 3 As PV-induced oral SCCs have a less aggressive clinical behavior, SCCs with p16 immunostaining have a more favorable prognosis. 7
In cats, SCCs are invasive neoplasms that frequently develop on the nasal planum and can be fatal. 4 Although these neoplasms are associated with ultraviolet (UV) light, PV DNA can be detected in around half of feline nasal planum SCCs suggesting that a proportion could be caused by PV infection. 11 If a feline nasal planum SCC is caused by PV infection, it may develop by a different carcinogenic pathway to an UV-induced SCC and could have a different clinical behavior. Therefore, using p16 immunostaining to subdivide feline nasal planum SCCs into those that are caused by PV infection and those that are not may provide valuable prognostic information. In addition, identification of a different clinical behavior within the 2 groups of feline SCCs will further support a causative association between PV infection and neoplasia development.
Fifty-one feline nasal planum SCCs were identified by retrospective surveys of databases at New Zealand Veterinary Pathology Ltd. and Gribbles Veterinary Pathology, Hamilton. Submitting veterinarians were contacted and information requested included whether or not the submitted sample was an excisional biopsy and the clinical outcome of the case. Cats that were lost to follow-up within 6 months of biopsy submission were excluded from the study. Histological examination revealed significant variation within the SCCs. Seven of the SCCs were predominantly confined to the epidermis with only small numbers of neoplastic cells visible in the superficial dermis. Examination of the epidermis within all 7 samples revealed irregular thickening and dysplasia that extended into follicular infundibula. This epidermal change has been described as a feature of Bowenoid in situ carcinomas 4 suggesting that these SCCs could have a PV etiology. Four of the SCCs that had Bowenoid features also contained cytopathic evidence of PV infection including swollen vacuolated cells with smudgy blue cytoplasm, further supporting a PV etiology. To the authors’ knowledge histological evidence of PV involvement in a feline nasal planum SCC has not been previously reported.
Immunostaining to detect p16 was performed as previously reported using a mouse antihuman p16 monoclonal antibody (BD Biosciences, San Jose, CA). 10 The p16 immunostaining within the SCCs was biphasic with either >70% of the cells showing intense nuclear and cytoplasmic immunostaining or weak immunostaining visible in less than 10% (Figs. 1 and 2). All 7 SCCs with histological features suggestive of PV infection exhibited intense p16 immunostaining (Fig. 3). Focal p16 immunostaining was visible within the epidermis adjacent to 3 p16-negative SCCs (Fig. 4). Nasal planum SCCs with p16 immunostaining were compared to p16-negative SCCs by analysis of variance and Kaplan–Meier and Cox regression analyses using PASW Statistics 18 (IBM Incorporated, Armonk, NY) (Table 1). As few cats with SCCs that had been submitted as excisional biopsies died of neoplastic disease, analyses of survival time were only performed in the 30 cats with SCCs that had been taken without curative intent. Cats with p16-positive SCCs had a longer estimated mean survival (643 days) than cats with p16-negative SCCs (217 days, P = .013, Fig. 5).
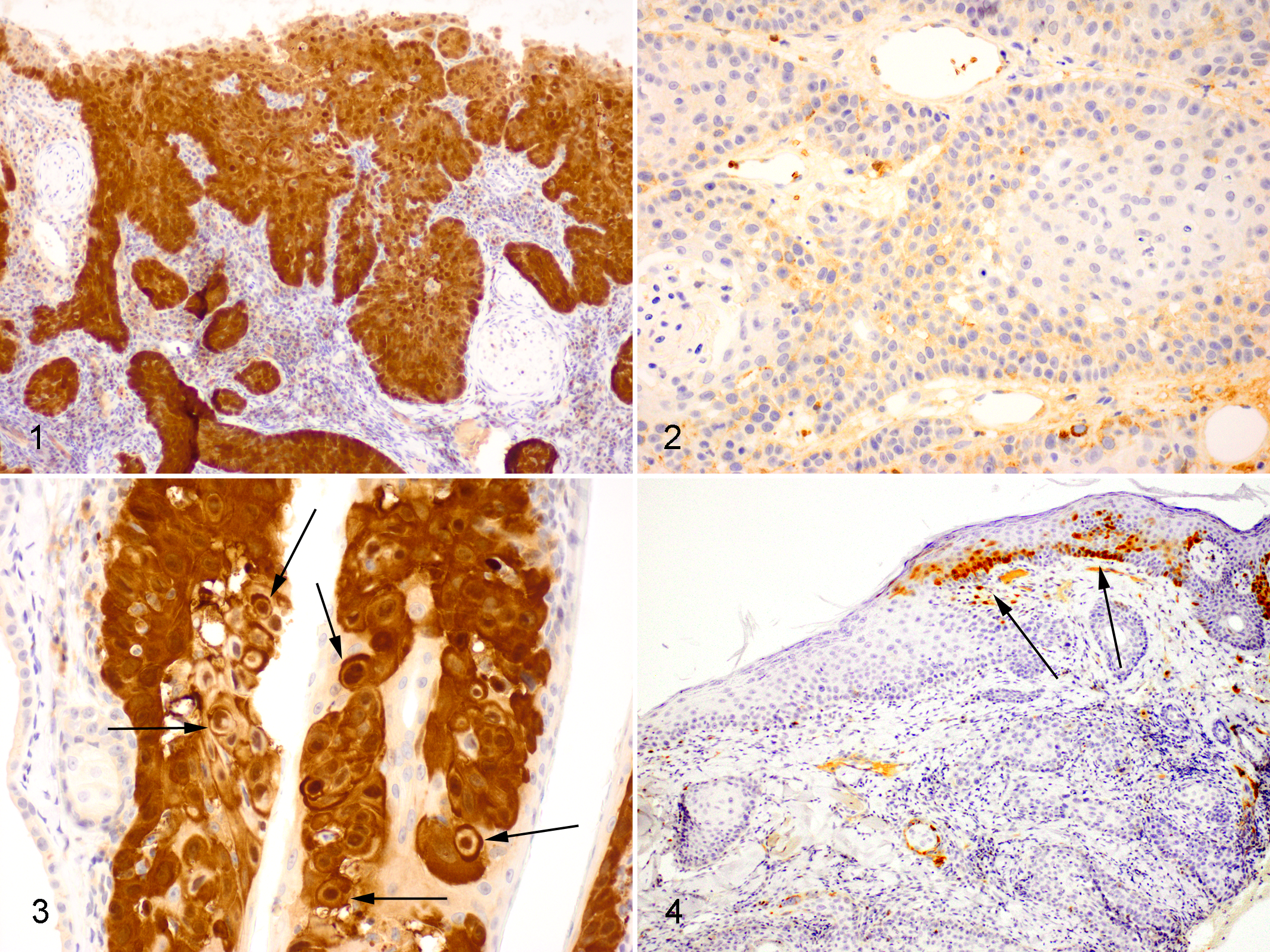
Feline nasal planum squamous cell carcinoma. Neoplastic cells exhibit intense nuclear and cytoplasmic immunostaining to antibodies against the p16CDKN2A protein. Note the presence of nests of neoplastic cells infiltrating from the overlying epidermis into the dermis. Bond Refine Detection staining kit with haematoxylin counterstain.
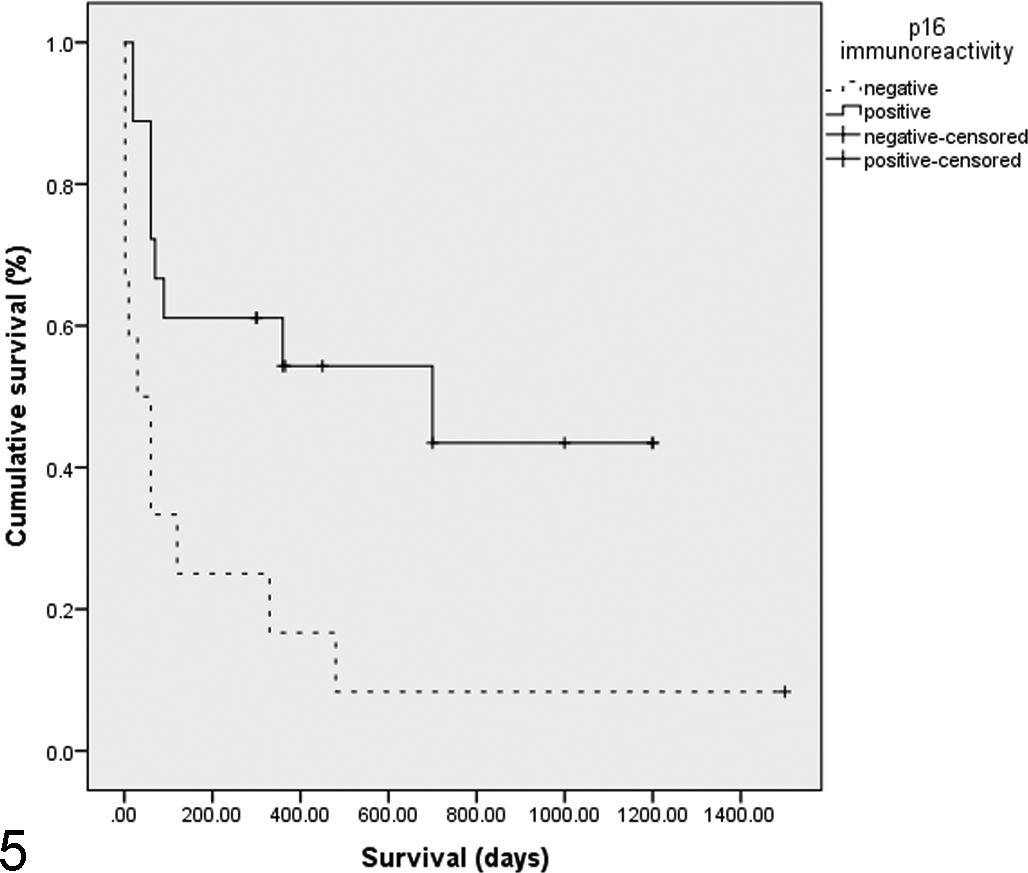
Kaplan–Meier survival curves for 30 cats with nasal planum squamous cell carcinomas (SCCs) where surgical excision was not attempted. The 2 lines illustrate SCCs that contained immunostaining to anti-p16CDKN2A antibodies in greater than 70% of the neoplastic cells (positive) and squamous cell carcinomas that showed minimal immunostaining within few neoplastic cells (negative). Cats with p16-positive SCCs survived significantly longer (P = .013).
Summary of p16 Immunostaining, Papillomaviral (PV) DNA amplification, and Survival Time of Cats With Nasal Planum Squamous Cell Carcinomas (SCCs).
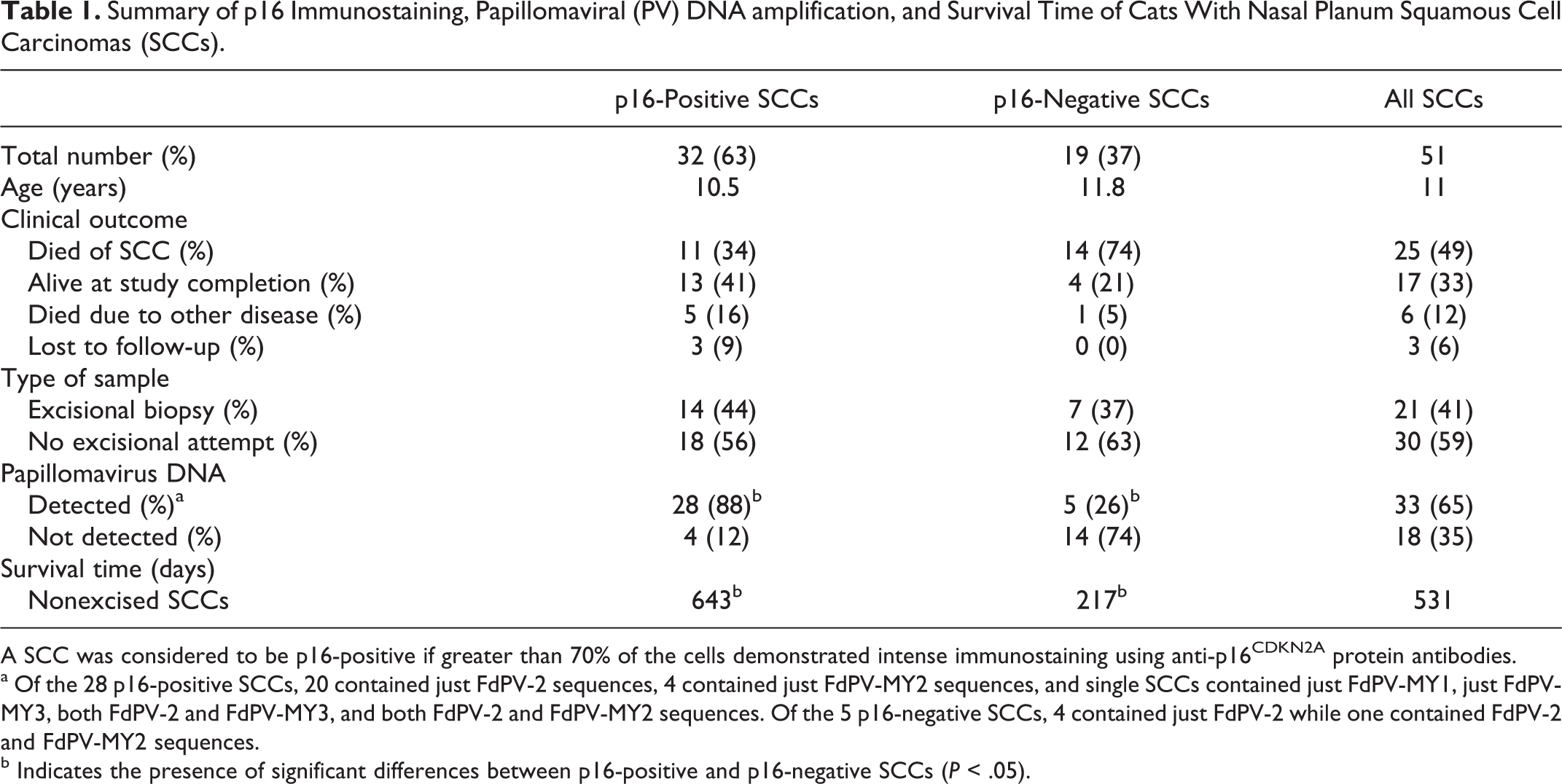
A SCC was considered to be p16-positive if greater than 70% of the cells demonstrated intense immunostaining using anti-p16CDKN2A protein antibodies.
a Of the 28 p16-positive SCCs, 20 contained just FdPV-2 sequences, 4 contained just FdPV-MY2 sequences, and single SCCs contained just FdPV-MY1, just FdPV-MY3, both FdPV-2 and FdPV-MY3, and both FdPV-2 and FdPV-MY2 sequences. Of the 5 p16-negative SCCs, 4 contained just FdPV-2 while one contained FdPV-2 and FdPV-MY2 sequences.
b Indicates the presence of significant differences between p16-positive and p16-negative SCCs (P < .05).
DNA was extracted from formalin-fixed paraffin-embedded samples as previously described. 11 Two primer sets were used to amplify PV DNA from the SCCs. These were the JMPF/R primer set which specifically amplifies Felis domesticus papillomavirus type-2 (FdPV-2) DNA 12 and the MY09/11 consensus primers that amplify many different PV types from multiple species. 2 DNA extracted from a feline viral plaque and a bovine fibropapilloma was used as a positive control for the JMPF/R and MY09/11 primers, respectively. Negative controls did not contain template DNA. The presence of amplifiable DNA within the samples was confirmed by amplifying a section of the feline p53 gene as previously described. 5 DNA amplified by the MY09/11 consensus primers was purified and sequenced as previously described. 11
Papillomaviral DNA was amplified from 33 of 51 (65%) nasal planum SCCs. This is the first evaluation of large numbers of feline nasal planum SCCs and the results confirm those of a previous study that detected PV DNA within 9 of 16 (56%) SCCs from this site. 11 All 7 SCCs with histological evidence of PV infection and all 3 p16-negative SCCs that were surrounded by p16-positive epidermis contained amplifiable PV DNA. The mean age of cats with SCCs that contained PV DNA (10.6 years) was not significantly different to those with SCCs that did not (11.8 years, P = .25). PV DNA was amplified more frequently from p16-positive SCCs than p16-negative SCCs (P < .001). Within the 30 cats with nonexcised SCCs, cats with SCCs that contained PV DNA had an estimated mean survival of 638 days, which was not significantly longer than cats with SCCs that did not contain PV DNA (263 days, P = .22). A supplemental table containing information for each cat is included in the online version of this article.
In this study cats with p16-positive SCCs survived longer than cats with p16-negative SCCs. This suggests that p16-positive SCCs may have a less aggressive clinical behavior. While p16 immunostaining is a well-established prognostic indicator in human oral SCCs, 7 these results are the first to identify a similar prognostic value of p16 within a nonhuman species. A significant advantage of using p16 to predict prognosis is the ‘all or nothing’ pattern of immunostaining. This allows rapid easy assessment and prevents interobserver variation. In humans, p16 immunostaining has also been used to predict the response of oral SCCs to treatment. 6 Variable treatment response is thought to be because p16-positive SCCs are more likely to retain functional apoptosis pathways. 15 It is possible that p16 immunostaining could predict the success of treatments that induce apoptosis in feline nasal planum SCCs.
While this study was designed to conform as closely as possible to the recommended guidelines for the conduct and evaluation of prognostic studies in veterinary oncology, 14 some limitations are recognized. As the SCCs were of variable size with variable depth of infiltration of the dermis it is possible that they were at different stages of development at the time that samples were taken. In addition, as the SCCs included in the study were from numerous primary care and referral veterinary practices it is probable that variable treatments were used in these cats. This lack of standardization in the SCCs would be expected to increase variability within the groups but not to introduce bias. However, a prospective study in which the SCCs and their subsequent clinical outcomes are better defined is required to confirm the findings of the present study.
Immunostaining using anti-p16 antibodies is considered to be synonymous with a PV etiology of a human oral SCC. 3 Human oncogenic PVs degrade retinoblastoma protein (pRb) resulting in increased p16 within the cell. 8 While p16 is increased, the loss of pRb prevents this protein from influencing cell cycling. 8 In cats, previous studies have revealed significant associations between the detection of PV DNA, loss of pRb and p16 immunostaining within cutaneous SCCs. 9 However, as PVs frequently asymptomatically infect feline skin, it is difficult to be certain that PVs cause the SCCs and are not present as secondary infections of the neoplastic epidermis. If all SCCs have the same cause with incidental PV infections present in a proportion, no differences in survival would be expected. Therefore, the significant differences in survival between p16-positive and p16-negative SCCs in the present study suggest that these SCCs develop by different carcinogenic pathways. As p16 immunostaining was associated with PV DNA within the SCCs this suggests a causative association between PVs and a proportion of feline nasal planum SCCs.
PV DNA was not amplified from 4 of 32 p16-positive SCCs. This was considered most likely to be due to degradation of PV DNA by formalin-fixation. The amplification of PV DNA from 5 of 19 p16-negative SCCs was most likely due to the presence of asymptomatic PV infection. The disparity between the detection of PV DNA and the results of p16 immunostaining in these cases most likely prevented detection of significant differences in survival times between cats with SCCs with PV DNA and those with SCCs without PV DNA. These results suggest that p16 immunostaining is a better prognostic indicator than the detection of PV DNA within feline nasal planum SCCs. Similarly, p16 immunostaining has been shown to more accurately predict prognosis than PV DNA detection in human oral SCCs. 7
Four different PV sequences were amplified from the feline nasal planum SCCs. Similar to previous studies, FdPV-2 was detected most frequently. 12 Three other PV DNA sequences were detected within the SCCs. Sequence FdPV-MY2 was the second most frequently amplified PV and has previously been detected in 3 feline nasal planum SCCs. 11 Two SCCs contained sequence FdPV-MY3. This sequence has only previously been detected in feline SCCs from the nasal planum, 11 suggesting that this PV may be restricted to this location. One SCC contained sequence FdPV-MY1 which has been previously amplified from feline viral plaques, 13 but has not previously been detected in a SCC. All 4 DNA sequences were associated with p16 immunostaining within the SCCs. These results suggest that there is some diversity in the PVs that can be amplified from feline nasal planum SCCs, but that all PVs can result in p16 immunostaining in infected cells.
In conclusion, feline nasal planum SCCs can be subdivided into those with p16 immunostaining and those without. Cats with p16-positive SCCs survived longer, suggesting that p16 immunostaining within a nasal planum SCC could provide useful prognostic information. The different survival times of the 2 groups of SCCs suggests that they could have different causes. As the presence of p16 immunostaining was significantly associated with the detection of PV DNA this provides additional evidence that PVs cause a proportion of feline cutaneous SCCs.
Footnotes
Acknowledgements
The authors acknowledge Dr Bronwyn Smits and the numerous veterinarians who provided information regarding the reported cases.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was funded by the Massey University Research Fund grant 11/0170.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.