
Abstract
Cutaneous viral plaques and bowenoid in situ carcinomas (BISCs) in cats are thought to be caused by papillomavirus (PV) infection. There is evidence that PVs may also cause some feline invasive squamous cell carcinomas (ISCCs). Human oncogenic PVs degrade retinoblastoma (RB) protein, impairing cell cycle control. Loss of RB function also increases p16CDKN2A protein (p16), and increased p16 immunoreactivity within a human oral ISCC indicates that the neoplasm was caused by PV infection. In the present study, p16 immunoreactivity was evaluated in 14 feline viral plaques, 14 BISCs, 7 non–solar-induced ISCCs, 11 solar-induced ISCCs, and 14 trichoblastomas. Increased p16 was present within all viral plaques, BISCs, and non–solar-induced ISCCs. In contrast, little p16 immunoreactivity was visible in the solar-induced ISCCs or trichoblastomas. PV DNA was consistently amplified from viral plaques, BISCs, and non–solar-induced ISCCs. However, just 5 solar-induced ISCCs and 1 trichoblastoma contained PV DNA. Given that both increased p16 immunoreactivity and PV DNA were present within viral plaques, BISCs, and non–solar-induced ISCCs, all 3 may be caused by PV infection. This suggests that feline non–solar-induced ISCCs may develop as a result of neoplastic progression from viral plaques and BISCs. Whether PVs promote this progression is unknown; however, evidence from this study suggests the PV that is associated with viral plaques and BISCs is able to disrupt the p16–RB pathway and therefore could have oncogenic potential. Immunohistochemical detection of p16 appears to be a useful technique to investigate the role of PVs in feline skin disease.
Keywords
Papillomaviruses (PVs) are estimated to cause 5.2% of all human cancers and are an established cause of squamous cell carcinomas (SCCs) of the cervix, penis, and mouth. 20 The causative PVs are the “high-risk” α-PV types. 7 An important oncogenic mechanism of these PVs is to impair cell cycle control by disrupting retinoblastoma (RB) protein function. 28 The abnormal RB results in an accumulation of p16CDKN2A protein (p16), which can be detected using immunohistochemistry. 21 Within human oral SCCs, given that high-risk PVs consistently cause RB dysfunction and also that RB dysfunction is rare in sporadic oral SCCs, p16 immunoreactivity indicates that the neoplasm was caused by PV infection. 22
Whether PVs cause human cutaneous SCCs is less certain. 13 Although β-PV DNA can be detected in most SCCs, β-PVs frequently infect human skin, making it difficult to confirm a causative relationship. 1 Increased p16 has been inconsistently reported within a variety of human cutaneous lesions. 3,12
In cats, there is strong evidence that cutaneous viral plaques and bowenoid in situ carcinomas (BISCs) are caused by PV infection. 8,14 Supporting a viral cause are the histological evidence of PV infection within viral plaques and the frequent presence of PV antigen within both viral plaques and BISCs. 5,8,26 BISCs probably represent malignant transformation of a viral plaque, and histological features of viral plaques are present within some BISCs. 26 Felis domesticus papillomavirus-2 (FdPV-2) DNA is detectible in both viral plaques and BISCs and is considered the likely causal agent. 10,14 –16 Whether PVs also cause cutaneous invasive SCCs (ISCCs) in cats is uncertain. 13 Although BISCs have been reported to progress to ISCCs, most ISCCs develop within areas of actinic keratosis as a result of chronic exposure to ultraviolet light. 8 Although FdPV-2 DNA is detectable in 85% of feline ISCCs, 14 50% of cats are asymptomatically infected by FdPV-2, 17 suggesting the PV could be an “innocent bystander” within the neoplasms.
The present study used IHC to detect p16 within feline viral plaques, BISCs, ISCCs, and trichoblastomas. As viral plaques and BISCs are thought to be caused by PV infection, these lesions were considered the most likely to contain increased p16. If p16 immunoreactivity was also increased in ISCCs, this would indicate that these develop because of similar cellular mechanisms, suggesting that PVs may also influence ISCC development. Trichoblastomas have not been associated with PV infection and were used to confirm that p16 immunoreactivity is not common within feline cutaneous neoplasms. Although p16 has been previously investigated in feline ocular sarcomas, 2 to the authors' knowledge, this is the first time that the presence of p16 has been investigated within skin lesions from a nonhuman species. Polymerase chain reaction (PCR) was used to investigate the presence of PV DNA within the samples.
Methods
Feline viral plaques, BISCs, ISCCs, and trichoblastomas were identified by retrospective surveys of databases at Massey University, New Zealand Veterinary Pathology Ltd., Cornell University, and Gribbles Veterinary Pathology Ltd. The fixation conditions of the neoplasms were unknown. The diagnosis within the database was confirmed by a single pathologist (JSM) using histological examination of a hematoxylin and eosin–stained section. ISCCs were further subdivided into solar-induced if they had been removed from the pinna or the nasal planum and non–solar-induced if they were removed from an area of the body not associated with solar exposure.
Sections for IHC were cut at a thickness of 5 μm, deparaffinized in xylene, rehydrated in graded ethanol, and rinsed in distilled water. Endogenous peroxidases were neutralized with 1% hydrogen peroxide in methanol for 15 minutes followed by rinsing for 5 minutes in distilled water. Antigen retrieval was achieved by incubating slides in epitope retrieval solution 2 (Leica Microsystems GmBH, Wetzlar, Germany) for 30 minutes at 100°C with a 20-minute cooldown. Slides were then incubated for 15 minutes with a mouse anti-human p16 monoclonal antibody (BD Biosciences, San Jose, CA) at a dilution of 1:200. A Bond Refine Detection staining kit (Leica Microsystems GmBH, Wetzlar, Germany) using 3,3′-diaminobenzidine substrate was used to visualize the immunoreaction, and sections were counterstained with hematoxylin. Epithelial cells within the dermal papilla exhibited consistent, low-intensity cytoplasmic positivity and were used as an internal positive control. The epidermis surrounding the lesion was used as a negative control. The specificity of anti-human p16 antibodies for feline p16 has been previously documented. 2
The intensity of the immunohistochemical reaction was graded on a scale of 0 to 5 with 0 indicating no visible immunoreactivity and 5 being intense immunoreactivity. The percentage of cells with cytoplasmic and the percentage with nuclear immunoreactivity were estimated by examining 5 different 400× fields within each lesion. Lesions were also assessed to determine whether immunoreactivity was distributed evenly within the neoplasm. Differences between groups were investigated by analysis of variance using SPSS 6 for Windows (SPSS Inc, Chicago, IL).
DNA was extracted from formalin-fixed paraffin-embedded samples as previously described. 15 Two primer sets were used in a nested PCR reaction. Initial amplification was by the consensus primers FAP59/FAP64, which was followed by amplification by the JMPF/JMPR primers, which were designed to specifically amplify FdPV-2. 14 The reaction conditions were as previously described. 14 DNA extracted from a bovine papilloma was used as a negative control. Because the specificity of the JMPF/JMPR primers for FdPV-2 has been previously demonstrated, 14 amplicons were not sequenced. Because of insufficient tissue, DNA could not be extracted from 6 viral plaques.
Results
Sixty feline cutaneous lesions including 14 viral plaques, 14 BISCs, 18 ISCCs, and 14 trichoblastomas were evaluated. Eleven of the ISCCs were classified as solar-induced and 7 as non–solar-induced. The solar-induced ISCCs included 8 from the pinna and 3 from the nasal planum, whereas 4 non–solar-induced ISCCs were from the face, 2 were from the paw, and 1 was from the ventrum.
Histologically, the viral plaques appeared as focal hyperplasia of the epidermis covered by increased quantities of keratin. All viral plaques contained histological evidence of PV infection including the presence of cells containing a dark shrunken nucleus surrounded by a clear cytoplasmic halo (koilocytes) and cells with increased quantities of blue-grey slightly granular or vacuolated cytoplasm (Fig. 1 ). The BISCs were all discrete plaques that contained full-thickness and follicular epidermal dysplasia with groups of cells that had loss of nuclear polarity (wind-blown appearance). Three of the BISCs contained histological features consistent with viral plaques toward the periphery of the lesions. Lesions were classified as ISCCs if penetration of the basement membrane by nests of epidermal cells was present. Central ulceration was visible within all ISCCs. Eight solar-induced ISCCs were surrounded by actinic keratosis that was characterized by focal hyperkeratosis and irregular epidermal hyperplasia and dysplasia. The trichoblastomas appeared as nodular circumscribed proliferations of epithelial cells arranged within broad trabeculae often with peripheral palisading.
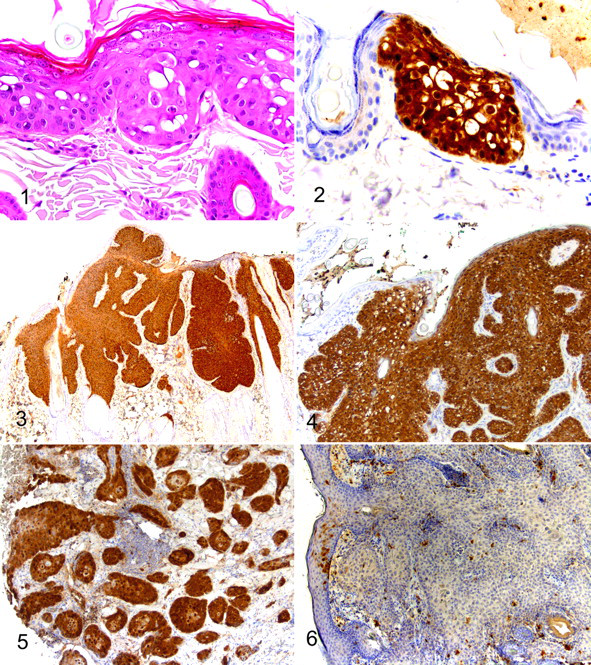
Results of the immunohistochemical evaluation are summarized in Table 1 . Examination of all viral plaques revealed intense immunoreactivity. Immunoreactivity was restricted to histologically abnormal keratinocytes within the plaques, and the surrounding epidermis did not contain p16 immunoreactivity (Fig. 2 ). Within large plaques, p16 immunoreactivity was present throughout all layers of the epidermis. However, immunoreactivity within developing plaques was confined to affected keratinocytes within the deeper layers of the epidermis. Immunoreactivity within BISCs was similar to that observed in viral plaques (Fig. 3 ). Therefore, p16 immunoreactivity was prominent within dysplastic epidermal cells with a sharp demarcation between BISCs and surrounding epidermis (Fig. 4 ). Immunoreactivity to p16 was consistently present in foci of dysplasia within hair follicles but not within surrounding histologically normal follicular epidermis. Nuclear and cytoplasmic immunoreactivity was visible within keratinocytes in both viral plaques and BISCs. FdPV-2 DNA was amplified from 8 of the 8 viral plaques and all 14 BISCs included in this study.
Summary of p16 Immunohistochemistry
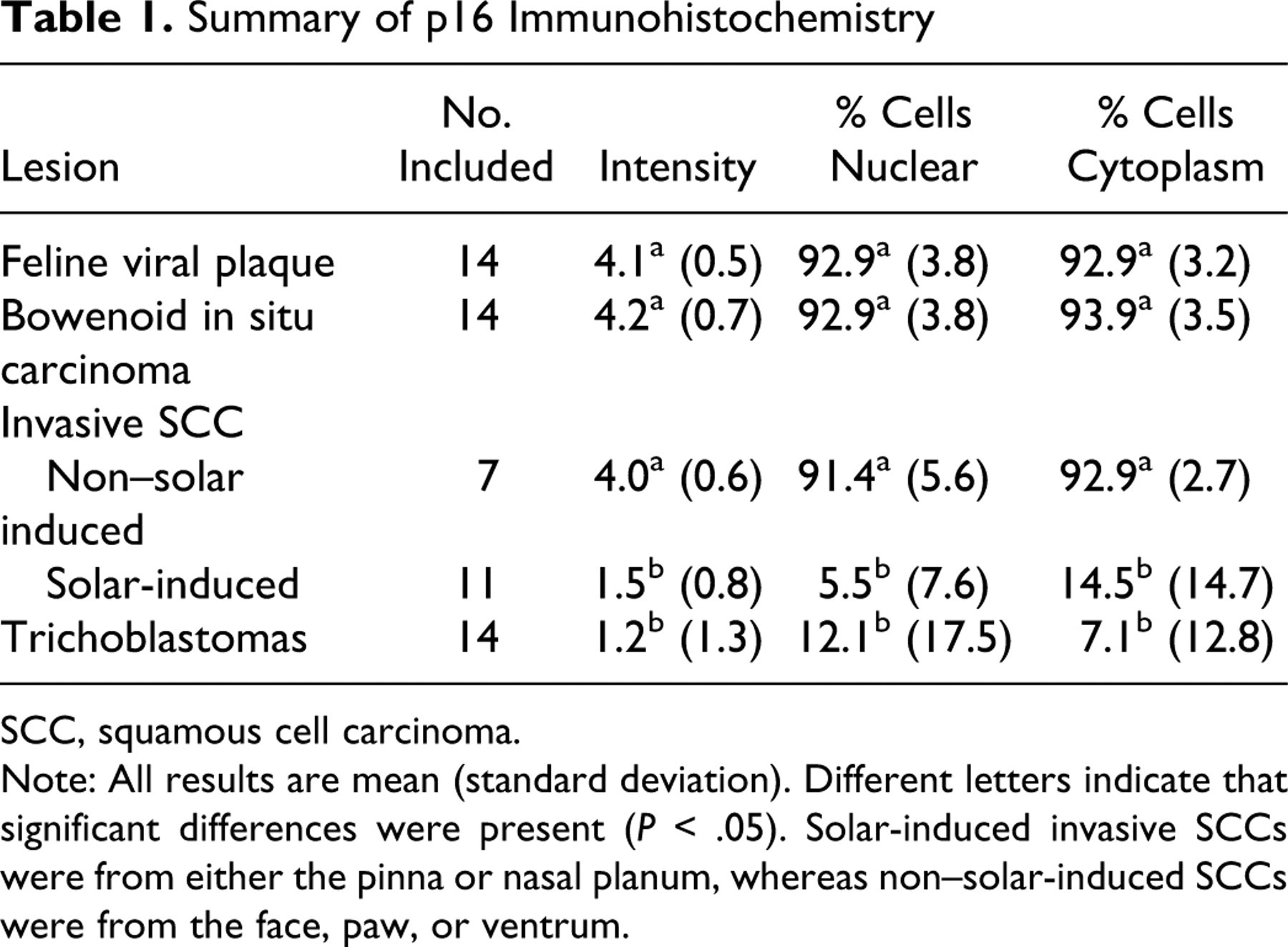
SCC, squamous cell carcinoma.
Note: All results are mean (standard deviation). Different letters indicate that significant differences were present (P < .05). Solar-induced invasive SCCs were from either the pinna or nasal planum, whereas non–solar-induced SCCs were from the face, paw, or ventrum.
Examination of all non–solar-induced ISCCs revealed consistent, intense p16 immunoreactivity. Immunoreactivity remained visible within nests of cells that had separated from the overlying neoplasm and had invaded into the underlying dermis (Fig. 5 ). Immunoreactivity was visible within hyperplastic and dysplastic epidermis at the periphery of the lesion but not within surrounding histologically normal epidermis. There were no significant differences in the intensity of immunoreactivity or proportion of neoplastic cells containing p16 immunoreactivity between non–solar-induced ISCCs and either BISCs or viral plaques. All 7 non–solar-induced ISCCs contained amplifiable FdPV-2 DNA.
In contrast, p16 immunoreactivity was significantly less intense within the solar-induced ISCCs than in the non–solar-induced ISCCs (P < .001). Additionally, the solar-induced ISCCs contained significantly fewer cells with either nuclear or cytoplasmic immunoreactivity (P < .001) (Fig. 6 ). Examination of 2 solar-induced ISCCs revealed p16 nuclear immunoreactivity within 20% of the cells and cytoplasmic immunoreactivity within 30%. One solar-induced ISCC contained cytoplasmic immunoreactivity within 50% of the neoplastic cells. However, the intensity of the immunoreactivity within these 3 ISCCs was low and its significance uncertain. Surrounding foci of actinic keratosis did not exhibit p16 immunoreactivity. Using PCR, FdPV-2 DNA was amplified from 5 of the 11 solar-induced ISCCs. Of the 3 solar-induced ISCCs that contained low-intensity p16 immunoreactivity, 2 contained FdPV-2 DNA whereas no FdPV-2 DNA was detectible in the other.
Examination of the trichoblastomas revealed less intense and less frequent immunoreactivity than that observed within the non–solar-induced ISCCs, viral plaques, and BISCs (P < .001). There were no significant differences in the intensity of immunoreactivity or the proportion of neoplastic cells with p16 immunoreactivity between solar-induced ISCCs and trichoblastomas. Amplifiable FdPV-2 DNA was present in 1 of the 14 trichoblastomas. This neoplasm did not contain increased p16 immunoreactivity.
Discussion
In the present study, cells within viral plaques and BISCs consistently contained intense p16 immunoreactivity. The increased p16 was restricted to hyperplastic and dysplastic keratinocytes, indicating that the cellular processes that altered keratinocyte morphology also increased p16. The present study supports previous studies that suggest FdPV-2 is the cause of viral plaques and BISCs. 10,14,16 The results indicate that FdPV-2 induces viral plaques and BISCs by mechanisms that also increase p16.
The p16CDKN2A protein (also referred to as the p16INK4a protein) inactivates cyclin-dependent kinases 4 and 6, which prevent phosphorylation of RB. 6,19 Unphosphorylated RB tightly binds to the E2F family transcription factors, arresting the cell cycle at the G1 phase. 6,25 As p16 and RB inhibit cell cycling, both are tumor suppressor proteins. 25 In humans, the ability of the high-risk PVs to cause cervical and oral SCCs is well established. 20 These oncogenic PVs disrupt RB function, damaging the G1-S cell cycle checkpoint and promoting neoplasm development. 25,28 The loss of RB also increases p16 within the cell. 21 This increased p16 does not inhibit cell division because RB is required for the tumor suppressor function of p16. 19 In the present study, FdPV-2 resulted in increased p16 within viral plaques and BISCs. However, the mechanism of this increase is unknown. It is possible that as in humans, p16 was increased as a result of PV-induced loss of RB function. If FdPV-2 does impair RB function, this PV is likely to promote neoplasm development. However, it is also possible that FdPV-2 infection increases p16 by a mechanism that does not impair cell cycle control. Therefore, the mechanism by which FdPV-2 increases p16 has to be identified before it can be confirmed that this virus is oncogenic. Additional studies to determine how FdPV-2 infection increases p16 are planned.
It is uncertain whether PVs cause cutaneous ISCC development in cats. 13 Most feline ISCCs develop as a result of malignant transformation of actinic keratosis and are considered to be solar-induced. 8 However, a subset of feline ISCCs develop within areas of the body that have dense hair or are darkly pigmented. The cause of these non–solar-induced ISCCs is unknown.
In the present study, both intense p16 immunoreactivity and amplifiable FdPV-2 DNA were present within all non–solar-induced ISCCs. Given that increased p16 and FdPV-2 DNA were also present within viral plaques and BISCs, all 3 lesions may develop because of a similar process. BISCs are thought to represent malignant transformation of a feline viral plaque. 26 Therefore, it is hypothesized that the non–solar-induced ISCCs initially developed as an undetected viral plaque that subsequently progressed through a BISC to an ISCC. Such progression is currently considered to be uncommon in cats. 8 However, the results of the present study suggest that many non–solar-induced feline ISCCs may develop as a result of malignant transformation of a viral plaque. Although both viral plaques and BISCs are thought to be caused by FdPV-2, 10,14 –16 it is unknown whether the PV influences malignant transformation. However, given that FdPV-2 infection disrupts the p16-RB pathway, it is possible that the PV could promote neoplasm development. These results suggest that if FdPV-2 infection can be prevented, this should reduce viral plaque and subsequent non–solar-induced ISCC development in cats.
Five of the 11 solar-induced ISCCs and 1 of the 14 trichoblastomas contained FdPV-2 DNA. However, none of the solar-induced ISCCs or trichoblastomas contained increased p16 immunoreactivity. Asymptomatic infection of feline skin by FdPV-2 has been detected in 50% of cats. 17 The frequent detection of FdPV-2 in feline skin samples makes it difficult to determine whether the PV has influenced lesion development or is an “innocent bystander.” The present study revealed that FdPV-2 can be present within a lesion without increasing p16 immunoreactivity. Therefore, evaluation of p16 immunoreactivity may allow differentiation between lesions caused by PV infection and lesions asymptomatically infected by PVs. In the present study, PV DNA was detected within feline solar-induced ISCCs. However, given that these did not contain increased p16, there is no evidence that PV infection caused these neoplasms. The possibility that FdPV-2 influences the development of solar-induced ISCCs by mechanisms independent of the p16-RB pathway cannot be excluded.
Increased p16 indicates a PV etiology within human oral SCCs. 22 However, the relationship between PV infection and p16 immunoreactivity in skin lesions is less well defined. 3 Although a strong correlation between the presence of PV DNA and p16 immunoreactivity was reported in one study, 18 no correlation was identified in another. 27 Although most studies report increased p16 immunoreactivity within human cutaneous SCCs, one study reported p16 immunoreactivity within only 10% of SCCs. 12 Similarly, studies of actinic keratosis have reported increased p16 in 7–100% of lesions. 9,23 Considering the inconsistent p16 immunoreactivity that has been reported in a wide range of human skin lesions, it appears that p16 immunoreactivity is a poor indicator of a PV etiology in human skin disease. In contrast, increased p16 was confined to feline viral plaques, BISCs, and non–solar-induced SCCs in the present study. No feline skin lesion contained intense p16 immunoreactivity without also containing evidence of PV infection. Additionally, none of the areas of actinic keratosis visible within the feline sections contained significant p16 immunoreactivity. These results suggest that p16 immunohistochemistry may be more valuable to investigate PV involvement in feline skin lesions than in human skin lesions.
The presence of PVs in feline skin disease can also be investigated using immunohistochemistry to detect PV antigen. Although PV antigen is consistently present in viral plaques, it is only detectible in 11–48% of BISCs. 11,26 In the present study, increased p16 was visible within all viral plaques and BISCs, suggesting that detection of p16 may be a more sensitive indicator of a PV etiology than detection of PV antigen.
In humans, p16 immunoreactivity can be used to predict the behavior of oral SCCs. 22,24 This is because p16 immunoreactivity indicates a PV etiology, and oral SCCs that are caused by PVs have a better prognosis than oral SCCs that are not. 4 In the present study, feline cutaneous ISCCs could similarly be subdivided into those that contain p16 immunoreactivity and those that do not. Additional studies are required to determine whether the detection of p16 can be used to predict the behavior of a feline cutaneous ISCC.
In conclusion, intense p16 immunoreactivity was visible within all viral plaques, BISCs, and non–solar-induced ISCCs in this study. All lesions that contained intense p16 immunoreactivity also contained evidence of FdPV-2 infection. This suggests that along with viral plaques and BISCs, non–solar-induced ISCCs may also be caused by PV infection. Although feline viral plaques have been previously reported to undergo malignant transformation, 8,26 more feline non–solar-induced ISCCs may develop from lesions caused by PV infection than is currently recognized. Although it is unknown whether FdPV-2 influences neoplastic transformation of a viral plaque, the apparent ability of this PV to disrupt the p16-RB pathway suggests a possible oncogenic mechanism. Although some solar-induced ISCCs contained FdPV-2 DNA, none contained increased p16, suggesting that these neoplasms develop by different cellular mechanisms. Therefore, this study provides no evidence that solar-induced ISCCs are caused by PV infection. The consistent and intense immunoreactivity against p16 within viral plaques and BISCs suggested that p16 immunohistochemistry may be a good indicator of PV infection within feline skin lesions. Therefore, this may be a valuable tool to investigate the relationships between PVs and feline cutaneous lesions.
Footnotes
The authors declared that they had no conflicts of interests with respect to their authorship or the publication of this article.
The authors declared that they received no financial support for their research and/or authorship of this article.