
Abstract
Although papillomaviral (PV) DNA is frequently present in feline cutaneous squamous cell carcinomas (SCCs), a causative association cannot be proven. Oncogenic human PVs cause neoplastic transformation by inhibiting retinoblastoma (pRb) and p53 activity. Therefore, absence of pRb and p53 immunostaining, along with increased p16 immunostaining, indicates a PV cause in some human SCCs. If PVs cause cutaneous feline SCCs, it was hypothesized that a similar immunohistochemistry profile, along with PV DNA, would be detectable. This was investigated using 5 feline viral plaques, 10 Bowenoid in situ carcinomas, 19 SCCs from ultraviolet-exposed (UV-exposed) skin, and 11 SCCs from UV-protected skin. Papillomaviral DNA was amplified by polymerase chain reaction from 30 of 45 lesions. Reduced pRb immunostaining was present in 26 of 45; increased p16 immunostaining was in 30; and p53 immunostaining was in 19. Both reduced pRb immunostaining and increased p16 immunostaining were more frequent in lesions containing PV DNA. In contrast, no association was observed between p53 immunostaining and the presence of PV DNA. SCCs from UV-protected skin more frequently contained PV DNA, reduced pRb, and increased p16 than UV-exposed SCCs. UV exposure was not associated with p53 immunostaining within the SCCs. These results suggest that feline PVs alter cell regulation by degrading pRb. Unlike oncogenic human PVs, there was no evidence that feline PVs degrade p53. These results provide further evidence that PVs may cause feline cutaneous SCCs, especially those in UV-protected skin, and they suggest a possible mechanism of this oncogenic action.
Keywords
Squamous cell carcinomas (SCCs) are the most common malignant skin neoplasm of cats. 16 While ultraviolet (UV) exposure is considered to cause most feline cutaneous SCCs, 16 papillomaviral (PV) DNA can be detected within a proportion of neoplasms, most frequently those from UV-protected areas of the body. 23 Although this detection of PV DNA suggests a possible causal association between PVs and feline cutaneous SCCs, PV DNA can be detected within non-SCC feline skin samples, making it difficult to determine whether the PVs cause SCC development or are present as “innocent bystanders.” 23
In humans, PVs are an established cause of genital and oral SCCs. 33 Oncogenic human papillomaviruses (HPVs) cause neoplastic transformation by inactivating and degrading both the retinoblastoma protein (pRb), which subsequently increases p16CDKN2A protein, and the transformation-related protein 53 (p53). 5,11,34,35 This action of HPVs is so consistent that immunohistochemistry can be used to determine if a oral SCC is caused by HPV infection. 6,36 A HPV-induced SCC demonstrates an absence of pRb and p53 immunostaining with an increase in p16. 10 ,20 In contrast, a SCC that develops due to chronic UV exposure typically demonstrates p53 and pRb immunostaining without an increase in p16. 8,32
The purpose of the present experiment was to determine whether the presence of PV DNA within a feline cutaneous SCC is associated with a immunohistochemical profile similar to that seen in PV-induced human cancers. Therefore, it was hypothesized that feline cutaneous SCCs could be divided into 2 groups. In the first group, neither pRb nor p53 immunostaining would be visible, but increased p16 and PV DNA would be present. This immunohistochemical prolife would suggest a PV cause, and these SCCs would be mostly from UV-protected areas of the body. The second group would contain pRb and p53 immunostaining without any increased p16 immunoreactivity, more consistent with the SCCs being UV induced. SCCs with this immunohistochemical profile would occur more commonly in UV-exposed skin and would less frequently contain PV DNA. As they are believed to be caused by PV infection, 24,26,29,38,41 feline viral plaques and Bowenoid in situ carcinomas (BISCs) were included with SCCs from UV-exposed and UV-protected areas of skin. Polymerase chain reaction was used to detect PV DNA within all lesions. To our knowledge, this is the first time that antibodies against human pRb have been validated and used in a nonlaboratory animal species.
Material and Methods
Feline viral plaques, BISCs, and invasive SCCs were identified by retrospective surveys of databases at Massey University, New Zealand Veterinary Pathology Ltd, and Gribbles Veterinary Pathology Ltd. The diagnosis was confirmed with histologic examination of an hematoxylin and eosin–stained section based on previously described criteria. 16 For the purpose of the study, locations on the body were subdivided into UV exposed, if the area contained little protective hair, whereas densely haired areas were considered UV protected. Therefore, SCCs on the pinna, nasal planum, eyelid, or third eyelid were considered UV exposed while UV-protected SCCs were from the face, digit, thigh, or neck.
DNA was extracted from formalin-fixed, paraffin-embedded samples as previously described. 25 Four primers sets were used to amplify PV DNA from the DNA extracted from the samples. These included the MY09/11 consensus primers that have been shown to amplify PV DNA from multiple species 9 and the JMPF/R primers that amplify Felis domesticus papillomavirus 2 (FdPV-2) DNA. 24 The JMY2F/R and JMY3F/R primers were used to amplify 2 PV DNA sequences that have been detected in feline cutaneous SCCs. 22 All polymerase chain reactions were carried out in duplicate. DNA extracted from a feline viral plaque was used as a positive control for the JMPF/R primers, while DNA extracted from a bovine fibropapilloma was used for the MY09/11 primers. DNA extracted from a bovine fibropapilloma was used as a negative control for the JMPF/R, JMY2F/R, and JMY3F/R primers, while the negative control for the MY09/11 primers did not contain template DNA. DNA amplified by the MY09/11 primers was purified and sequenced as previously described. 25 Results were compared with known sequences in GenBank (http://www.ncbi.nlm.nih.gov/genbank) using the basic local alignment search tool (http://www.ncbi.nlm.nih.gov/blast). Due to the previously demonstrated specificity of the JMPF/R, JMY2F/R, and JMY3F/R primers, the DNA amplified by these primers was not sequenced. 22,24
As antibodies against pRb have not been used in cats, Western blotting was used to confirm the specificity of the antibodies used to feline pRb. Briefly, tissue lysates of feline liver were prepared and diluted in Laemmli buffer to a final protein concentration of 2.5 mg/mL and heated at 100°C for 5 minutes. A Jurkat cell lysate (BD Biosciences, San Jose, CA) was used as a positive control. Samples were loaded onto a SDS-PAGE 7.5% gel (Criterion Precast Tris-HCl Gel, BioRad, Hercules, CA) and separated by electrophoresis at 100 V for 60 minutes in 1× Tris-glycine-SDS buffer. Proteins were transferred onto nitrocellulose membrane, and nonspecific binding was blocked by incubation in 5% skimmed milk in PBS/Tween 20 for 1 hour at room temperature. Membranes were incubated overnight at 4°C with mouse anti-human pRb (BD Biosciences) at a dilution of 1:100. Membranes were incubated with horseradish peroxidase–conjugated secondary antibodies (BD Biosciences) for 1 hour at room temperature, and immune complexes were visualized using a chemiluminescent detection system (Immun-Star HRP Chemiluminescent Kit, BioRad).
Sections for immunohistochemistry were cut at a thickness of 5 μm, deparaffinized in xylene, rehydrated in graded ethanol, and rinsed in distilled water. To perform immunohistochemistry to detect pRb, sections were heated in a pressure cooker for 10 minutes in 0.01M citrate buffer, pH 6.0, with a 10-minute cooldown. Endogenous peroxidases were blocked by incubating slides in 0.3% hydrogen peroxide in 0.05M Tris-buffered saline, pH 7.6. Nonspecific staining was blocked by incubating slides with equine serum (Vector Laboratories, Burlingame, CA) for 20 minutes. Slides were incubated for 60 minutes with anti-human pRb antibodies (BD Biosciences) at a dilution of 1:40. Slides were incubated for 30 minutes with biotinylated horse anti-mouse/rabbit secondary antibody (Vector Laboratories). Slides were then incubated for 30 minutes with a biotin–avidin complex (Vector Laboratories), followed by 3,3-diaminobenzidine substrate (Liquid DAB Substrate Chromagen System, Dako, Carpinteria, CA) and counterstained with Gill’s hematoxylin. Cells within the adjacent nonneoplastic epidermis were used as internal positive controls, while the primary antibody was omitted in negative controls.
Immunohistochemistry to detect p16 was based on a mouse anti-human p16 monoclonal antibody (BD Biosciences) at a dilution of 1:200 as previously reported. 21 Epithelial cells within the dermal papilla exhibited consistent low-intensity cytoplasmic positivity and were used as an internal positive control, while the epidermis surrounding the lesion was used as a negative control.
To detect p53, a mouse anti-human p53 clone pAb 240 antibody (BD Biosciences) was used as previously reported 1 except that the antibody was diluted to 1:100. A human neoplasm known to contain p53 mutations was used as a positive control, and the epidermis surrounding the lesion was used as a negative control. The pAb 240 p53 clone was used because it has also been reported to produce less background staining than the CM-1 p53 clone, which has been reported to react with feline p53. 1
For all examined antibodies, the percentage of cells with nuclear immunoreactivity was estimated by examining 5 different 400× fields within each lesion. Lesions were also assessed to determine if immunoreactivity was distributed evenly within the neoplasm. Lesions were considered to contain increased p16 when greater than 75% of the cells contained intense immunostaining. When pRb was assessed, lesions were divided into immunostaining visible in less than 25% of the cells, in 25% to 50% of the cells, and in over 50% of the cells; pRb was considered to be reduced within a lesion if less than half the cells contained immunostaining. Lesions were classified as having immunostaining against p53 if intense immunostaining was visible within over 20% of the cells. Differences between groups were investigated by analysis of variance using SPSS 16 (SPSS Inc, Chicago, IL).
Results
Forty-five samples were included in this study, including 5 feline viral plaques, 10 BISCs, and 30 invasive SCCs. Of the invasive SCCs, 19 were from UV-exposed skin, including 8 from the pinnae, 6 from the nasal planum, 4 from the eyelids, and 1 from the third eyelid. Eleven SCCs were from UV-protected skin, including 5 from the face, 2 from the lips, 2 from the digits, and 1 each from the ventrum and neck.
The 4 sets of primers used in the experiment amplified PV DNA from 30 samples, including all viral plaques and BISCs (Table 1). Of 30 SCCs, 15 contained PV DNA. PV DNA was detected more frequently in SCCs from UV-protected areas (9 of 11) than in UV-exposed SCCs (6 of 19) (P = .01). Of the 30 samples that contained PV DNA, 29 contained DNA that was amplified by the primers specific for FdPV-2. One SCC from a UV-protected area contained only the previously described FdPV-MY2 DNA sequence. 26 Both the FdPV-MY2 sequence and FdPV-2 were detected in 2 SCCs and a BISC. One viral plaque contained FdPV-2 and the previously described FdPV-MY1 DNA sequence, 27 while one SCC contained FdPV-2 and the previously described FdPV-MY3 DNA sequence. 22
Polymerase Chain Reaction and Immunohistochemistry Results, No. (%) a
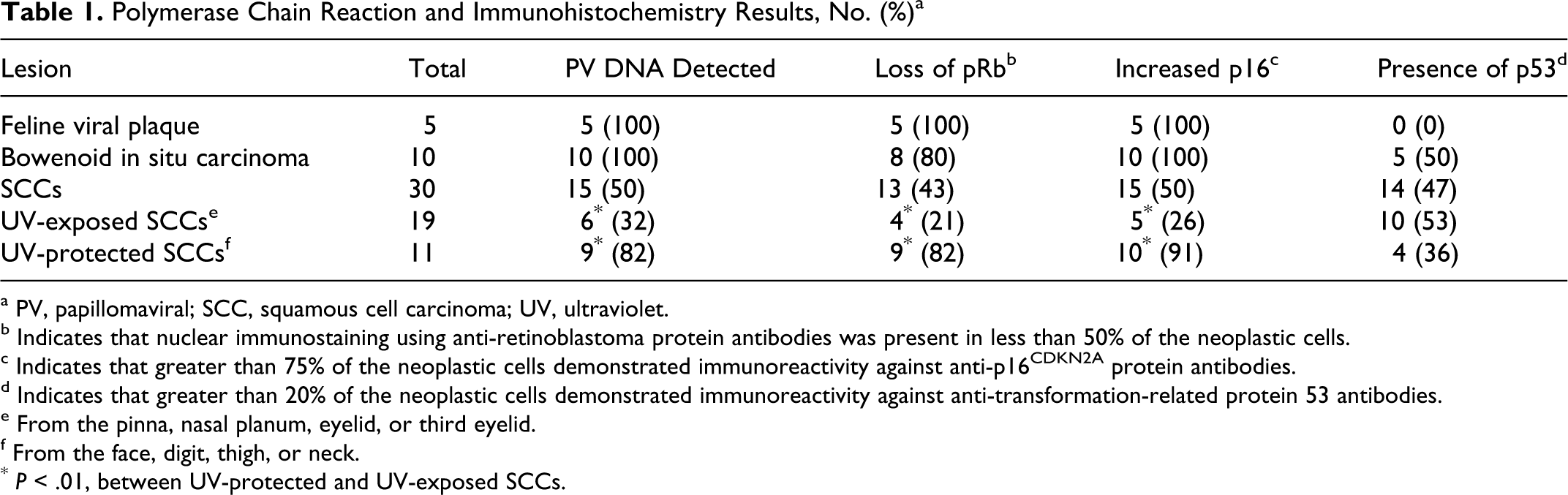
a PV, papillomaviral; SCC, squamous cell carcinoma; UV, ultraviolet.
b Indicates that nuclear immunostaining using anti-retinoblastoma protein antibodies was present in less than 50% of the neoplastic cells.
c Indicates that greater than 75% of the neoplastic cells demonstrated immunoreactivity against anti-p16CDKN2A protein antibodies.
d Indicates that greater than 20% of the neoplastic cells demonstrated immunoreactivity against anti-transformation-related protein 53 antibodies.
e From the pinna, nasal planum, eyelid, or third eyelid.
f From the face, digit, thigh, or neck.
* P < .01, between UV-protected and UV-exposed SCCs.
Western blotting using anti-pRb antibodies demonstrated a 110-kDA fragment from the positive control and feline tissues, confirming the specificity of this antibody for feline pRb. Immunoreactivity using anti-pRb antibodies was confined to the nucleus within all samples. Within the 45 lesions, pRb immunoreactivity was visible within less than 25% of the cells in 18 lesions: 3 viral plaques, 5 BISCs, and 10 SCCs (Figs. 1 –3). Immunoreactivity was present in 25% to 50% of the cells within 2 viral plaques, 3 BISCS, and 3 SCCs. Therefore, reduced pRb immunoreactivity was observed in 26 lesions, including all 5 viral plaques, 8 BISCs, and 13 SCCs. Reduced pRb immunoreactivity was more frequent in lesions that contained PV DNA (24 of 30) than in lesions without detectable PV DNA (2 of 15) (P < .001) (Table 2). Within the 30 SCCs, p53 immunostaining was not visible more frequently in SCCs from UV-exposed skin (10 of 19) than in SCCs from UV-protected skin (4 of 11) (p = 0.32).
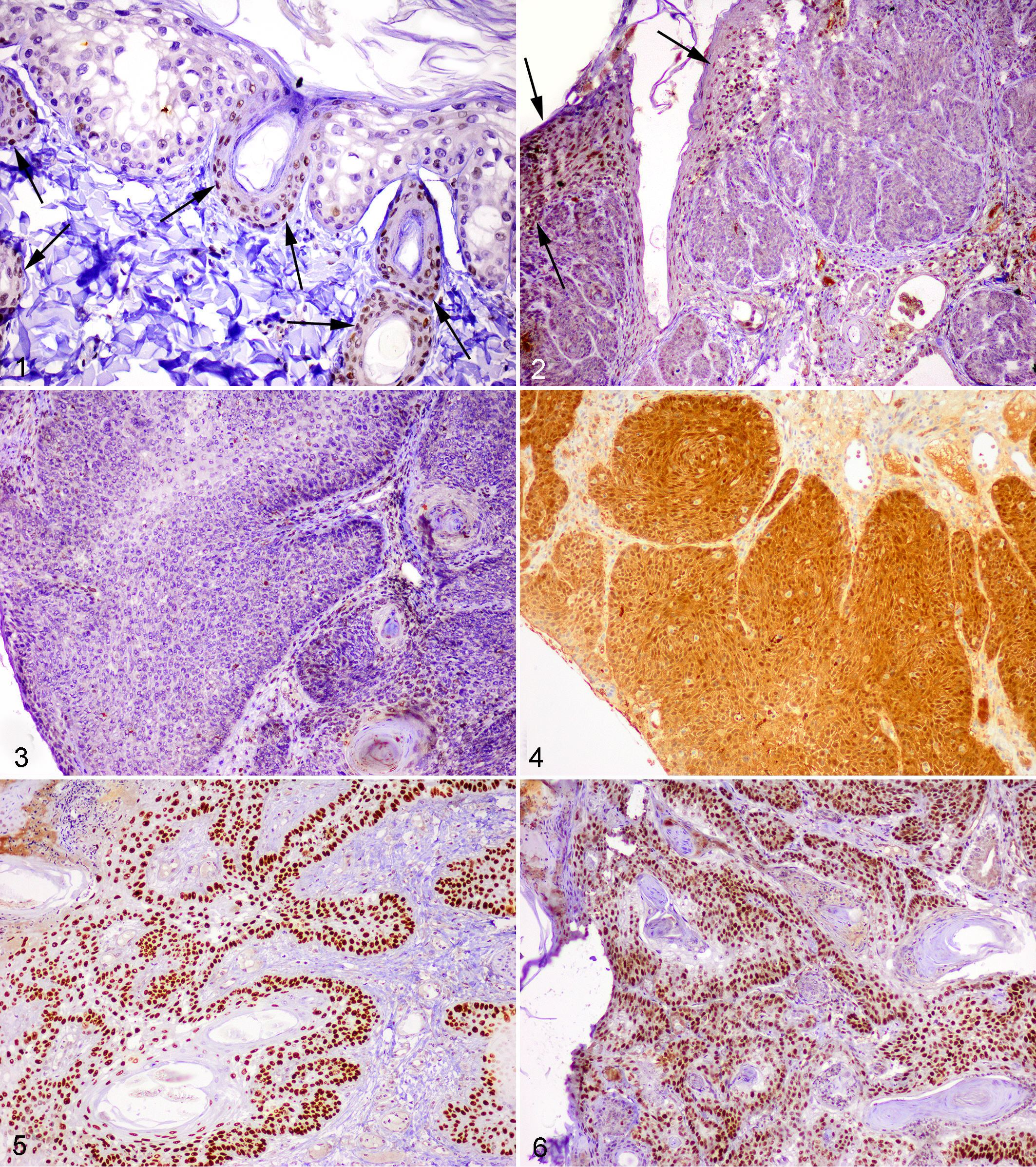
Feline viral plaque. Cells within the epidermis show cytopathologic changes consistent with papillomavirus infection, including enlargement and cytoplasmic vacuolation. Cells showing cytopathologic changes contain loss of nuclear immunoreactivity against anti-retinoblastoma protein antibodies. Keratinocytes within the surrounding normal epidermis have retained nuclear immunostaining (arrows). Avidin–biotin–peroxidase with hematoxylin counterstain.
Presence of Papillomaviral DNA Within a Lesion, No. (%) a
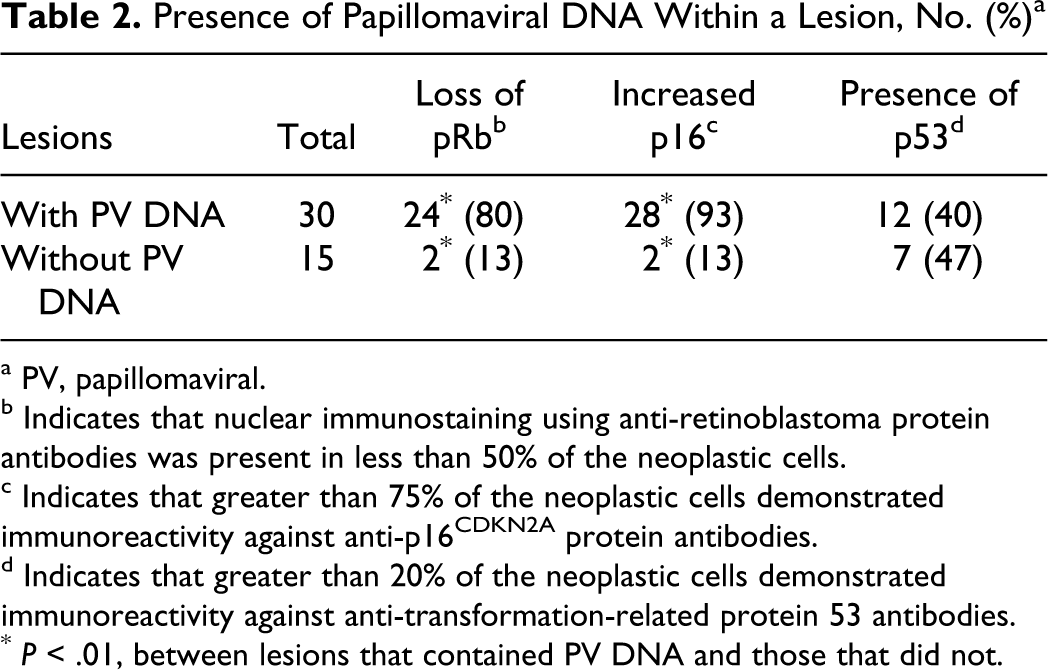
a PV, papillomaviral.
b Indicates that nuclear immunostaining using anti-retinoblastoma protein antibodies was present in less than 50% of the neoplastic cells.
c Indicates that greater than 75% of the neoplastic cells demonstrated immunoreactivity against anti-p16CDKN2A protein antibodies.
d Indicates that greater than 20% of the neoplastic cells demonstrated immunoreactivity against anti-transformation-related protein 53 antibodies.
* P < .01, between lesions that contained PV DNA and those that did not.
Immunoreactivity against p16 was present within both the nuclei and the cytoplasm. All lesions contained either marked immunoreactivity within most cells or minimal immunoreactivity within less than 25% of the cells. Intense immunoreactivity was present within all viral plaques and BISCs and within 15 SCCs. Increased p16 was present more frequently in lesions that contained PV DNA (28 of 30) than in lesions without detectable PV DNA (2 of 15) (P < .001). Within the SCCs, increased p16 was more frequent in SCCs from UV-protected sites (10 of 11) than from UV-exposed sites (5 of 19) (P = .001). Of the 30 lesions that contained increased p16, loss of pRb immunoreactivity was present within 26 (Fig. 4). No lesions contained reduced pRb immunostaining without a concurrent increase in p16 immunostaining.
Immunostaining using anti-p53 antibodies was confined to the nuclei. Immunoreactivity was visible within 19 of 45 lesions—5 BISCs and 14 SCCs (Figs. 5, 6). There was no significant difference in the frequency of p53 immunostaining between lesions that contained PV DNA (12 of 30) and those that did not (7 of 15) (P = .45). There were no significant differences in p53 immunostaining between lesions with reduced pRb (P = .183) or those with increased p16 (P = .227). Within the 30 SCCs, more SCCs from UV-exposed skin (10 of 19) did not contain p53 immunostaining than SCCs from UV-protected skin (4 of 11) (P = .32).
Discussion
In cats, there is strong evidence that PVs cause viral plaques and BISCs. 24,26,29,41 Viral plaques can progress to BISCs, 41 and BISCs have been reported to progress to SCCs, 16 but the proportion of feline SCCs that develop from BISCs and the role that PVs play in this progression are unresolved. PV DNA was amplified from 50% of the SCCs in the present study. However, as PVs can asymptomatically infect feline skin, 28 the detection of PV DNA within a cutaneous SCC does not prove that the virus influenced neoplasm development. As has been reported, 22 PV DNA was detected in the present study more frequently in SCCs from UV-protected areas than in SCCs from UV-exposed skin. In people, protection from UV light decreases asymptomatic PV skin infection. 14 This suggests that the increased detection of PV DNA within feline cutaneous SCCs that were protected from UV light was not due to an increased frequency of asymptomatic infection.
As with cats, many human PV infections are asymptomatic, 3 making it difficult to determine the significance of HPV DNA within a neoplasm. To help differentiate between an HPV cause and an asymptomatic infection, immunohistochemistry can be used to detect changes to cell regulatory proteins that are caused by HPVs. One of these is the HPV-induced inactivation and degradation of pRb. 5 ,11 The pRb inhibits progression from the G1 phase to the S phase of the cell cycle, and loss of pRb removes an important checkpoint preventing cell division. 37 This mechanism of neoplastic transformation is so consistent that demonstration of reduced pRb within an oral or anogenital SCC is considered indicative of a HPV cause. 2,20,40 In the present study, reduced pRb immunostaining was present in 26 of the 45 feline preneoplastic and neoplastic lesions. Of the 26 lesions that demonstrated reduced pRb, PV DNA was amplified from 24. As in previous studies, 24,30 FdPV-2 was the PV most frequently amplified from the feline lesions. The ability of HPVs to interact with pRb is dependent on a LXCXE motif within the E7 gene. 11 This motif has been identified in FdPV-2. 18 Therefore, the significant association between the presence of PV DNA and reduced pRb suggests that FdPV-2 can bind to and degrade pRb. Within the PVs that infect nonhuman species, only the cottontail rabbit papillomavirus has been shown to degrade pRb. 15,17 However, this experiment was performed in cell cultures, and to our knowledge, an association between PV infection and loss of pRb has not been demonstrated in samples of naturally developing lesions from a nonhuman species.
PV DNA was not amplified from 2 of the 26 lesions that contained reduced pRb immunostaining. This could suggest that loss of pRb is not specific for a PV cause in feline cutaneous SCCs. Alternatively, it is possible that infection by a PV had reduced pRb, but the PV was not detected within the SCC. Multiple PV types, including HPVs, have been detected within feline cutaneous SCCs. 25,29,30 Therefore, while consensus primers were used in this study, it is possible that other PV types were present that were not amplified by these primers. Additionally, the tissues used in the study had been formalin fixed. Such fixation can fracture DNA, making amplification difficult, especially if only small quantities of PV DNA are present. 19
In humans, the presence of increased p16 immunostaining is also used to indicate a PV cause within SCCs from some anatomical locations. 10,20 As p16 regulates cell division by a mechanism that is dependent on the presence of pRb, 37 loss of pRb increases p16 within the cell. 34 In the present study, all feline lesions with reduced pRb immunostaining also contained increased p16 immunostaining, suggesting that inactivation of pRb consistently increased p16 within the feline lesions. However, 2 SCCs and 2 BISCs contained increased p16 without reduced pRb immunostaining. While this could be evidence that p16 can increase independently of pRb, studies of HPVs suggest that the primary action of the PV is to inhibit pRb function, with accelerated protein degradation as a secondary function. 5,11 Therefore, pRb can remain immunohistochemically detectable within human PV-induced SCCs despite being functionally inactive. 20 Associations between increased p16 and the presence of PV DNA within feline cutaneous SCCs have been reported. 21,22 However, this is the first evidence to suggest that feline PV infection could increase p16 by degrading pRb. While these results suggest that feline PV infection may alter cellular regulation by disrupting the pRb/p16 pathway, it cannot be confirmed that this action influences neoplastic transformation of the cells.
In humans, a PV-induced SCC is expected to contain PV DNA with reduced pRb, and increased p16, immunostaining. In the present study, all PV-induced plaques and BISCs contained PV DNA and increased p16 with reduced pRb visible in 13. PV DNA with reduced pRb and increased p16 immunostaining was also observed in 11 SCCs. This suggests that 11 of the 30 SCCs in the present study were caused by PV infection. Nine SCCs with this profile developed in UV-protected skin, suggesting that they may have developed as a progression of a PV-induced BISC.
The strong correlation between pRb loss and increased p16 observed in the present study suggests that it is unnecessary to evaluate both proteins to detect changes in the pRb/p16 pathway. Immunoreactivity against p16 was biphasic within the feline lesions, making determination of p16 status quick and easy. In contrast, detection of reduced pRb by immunohistochemistry required estimating the proportion of cells that retained pRb immunostaining and so was more time-consuming. Therefore, immunohistochemistry to detect p16 appears to be a better method to detect changes in the pRb/p16 pathway within feline samples. In human oral SCCs, immunohistochemistry to detect p16, rather than pRb, is currently used (along with molecular techniques) to investigate a PV cause. 6,36
The p53 protein functions as a tumor suppressor, maintaining genetic integrity and promoting apoptosis in response to DNA damage. 37 Mutations within the p53 gene prevent normal p53 function and result in a stable p53 protein that is detectable immunohistochemically. 13,42 Oncogenic HPVs degrade p53 7,31,35 so that p53 immunostaining is not present in human PV-induced SCCs. 20 In contrast, in the present study, there was no significant association between p53 immunostaining and the presence of PV DNA within the lesions, which suggests that feline PVs do not degrade p53. This is consistent with bovine PV-induced equine sarcoids that also demonstrate p53 immunostaining in a proportion of tumors. 4 Likewise, no interaction between cottontail rabbit papillomavirus proteins and p53 could be detected. 15 In the present study, immunostaining against p53 was visible in 5 of 10 BISCs. A previous study of 22 BISCs detected p53 immunostaining in just 4. 12 The reasons for the different rates of p53 immunoreactivity are uncertain. However, it is possible that p53 mutations develop spontaneously as dysplasia within the BISC increases. This is supported by the absence of p53 immunostaining and dysplasia within the 5 viral plaques. If p53 mutations develop as a preneoplastic lesion progresses to neoplasia, it is possible that the BISC lesions included in the present study were more advanced than the BISC lesions included in the previous study. 12
In the present study, p53 immunostaining was present within 14 of 30 cutaneous SCCs. Previous studies of feline SCCs have detected p53 in 13 of 20 cutaneous feline SCCs 39 and 7 of 9 feline cutaneous and oral SCCs. 1 In people, UV exposure causes p53 gene mutations, and p53 immunostaining is frequently present in human UV-induced skin cancers. 32 In cats, p53 immunostaining has been associated with UV exposure in preneoplastic cutaneous lesions. 12 In contrast, no significant association between UV exposure and p53 immunostaining was observed in the present series of feline SCCs. This could be due to the development of additional mutations within the p53 gene that result in the loss of immunostaining within some SCCs and the development of immunostaining in others.
In conclusion, the presence of PV DNA within the feline preneoplastic and neoplastic cutaneous lesions was associated with reduced pRb immunostaining. This suggests that, similar to oncogenic HPVs, feline PVs may influence cell regulation by inactivating and degrading the pRb protein. Reduced pRb was associated with increased p16 within the neoplastic cells, and p16 immunostaining appears to be a reliable indicator of reduced pRb function within a cell. Loss of pRb and, subsequently, increased p16 is considered a specific indicator of a PV cause in human oral and anorectal SCCs. 10,20 If loss of pRb within feline lesions similarly indicates a PV cause, the results of the present study suggest that PV infection could cause most feline SCCs from UV-protected skin and a significant proportion of UV-exposed SCCs. The results of this study provide additional evidence that the pathogenesis of feline cutaneous SCCs shows some similarities to the pathogenesis of human cervical and oral SCCs. However, in contrast to oncogenic HPVs, feline PVs do not appear to degrade p53.
Footnotes
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The research was supported by a grant from the Massey University Research Fund.