
Abstract
A 15-year-old American Quarter horse mare was euthanized because of poor response to therapy for severe diarrhea. Significant gross findings were limited to the large intestines. The walls of the cecum and colon were thickened with widely scattered nodules in the mucosa and submucosa that extended into the enlarged colic lymph nodes. Microscopically, there was severe granulomatous typhlocolitis, lymphangitis, and lymphadenitis, with many intralesional Gram-positive, non-acid-fast coccobacilli and few cyathostomes. Intralesional bacteria were immunohistochemically and polymerase chain reaction (PCR) assay positive for Listeria monocytogenes. Concurrent infection with Salmonella enterica serovar Typhimurium was detected by PCR and culture. Infection with L. monocytogenes in horses is rare, and coinfection with Salmonella and small strongyles probably contributed to the development of granulomatous typhlocolitis.
Listeria monocytogenes, a facultative intracellular bacterium, is a ubiquitous and highly adaptable, opportunistic pathogen that is resistant to adverse conditions and survives long term in the environment. It has been isolated from surface soils, decaying vegetation, pasture herbage, silage, sewage sludge, feces, and rivers. 5
Among domestic animals, L. monocytogenes infection is most common in ruminants, which are often infected via ingestion of contaminated foodstuffs (eg, silage). Infection often manifests as encephalitis and associated neurologic disease, because the bacterium invades the oral mucosa and travels retrograde to the brain via the trigeminal nerve. 5 Intrauterine infection leads to ruminant abortion 7 or neonatal sepsis. 5 Infected small ruminants, such as sheep, can develop suppurative abomasitis and enteritis or serve as asymptomatic carriers.3,9
Listeriosis is rarely reported in horses. In Icelandic horses with diarrhea and gastroenteritis, L. monocytogenes infection was most often attributed to contaminated feed, including silage. The organism was isolated from feces and tissues of fatal equine cases; however, gross and histologic lesions were not described. 4 In the current report, the clinical course, necropsy, histologic, and immunohistochemical findings, as well as microbiological characterization, indicated concurrent infection of an adult horse with L. monocytogenes, Salmonella enterica serovar Typhimurium, and cyathostomes.
History
Fresh and formalin-fixed large intestine and colic lymph nodes were received from a dead 15-year-old American Quarter horse mare from northwestern Georgia. One horse had died on the premises during the spring or summer of each of the 3 previous years. All 4 horses had had similar clinical signs, including lethargy, weight loss, and profuse watery diarrhea, followed by death within 1 week of the onset of illness. The current case was the first horse from the farm to receive postmortem diagnostic evaluation. The horse had lost over 136 kg in body weight and was euthanized because of poor response to a 10-day course of metronidazole treatment. Horses were dewormed 3 to 4 times a year. The 12 horses on the farm shared the same pasture, which had previously been used for beef cattle.
At necropsy, performed immediately after euthanasia, the horse was in poor nutritional condition. Other salient gross findings were limited to the cecum and all colonic segments. The serosal, mucosal, and cut surfaces of the large intestine and colic lymph nodes had numerous, 5- to 20-mm-diameter, pale gray/tan nodules, with a yellow/tan exudate on cut section. Intestinal mucosa was diffusely and irregularly thickened, with widely scattered, shallow ulcers.
Diagnostic Methods
Tissue samples were fixed in neutral-buffered, 10% formalin solution. Trimmed tissues were routinely processed, embedded in paraffin, and sectioned at approximately 5 μm. All tissue sections were stained with hematoxylin and eosin (HE); some sections were also stained with Lillie Twort Gram’s stain and Ziehl-Nielsen acid-fast stain.
The DNA was extracted from large intestinal mucosal scrapings using an UltraClean fecal extraction kit (MO BIO, Carlsbad, California) and amplified using traditional and real-time polymerase chain reaction (PCR) procedures for the detection of Salmonella enterica subsp enterica, 2 Yersinia spp, 8 Listeria spp, 6 and L. monocytogenes. 1 Bacterial culture was performed on intestinal contents and on a colonic mural nodule. The samples were plated on blood, MacConkey, and phenyl alcohol blood agars (Remel, Lenexa, Kansas) and incubated overnight at 37°C. β-Hemolytic colonies were isolated and further characterized by Gram staining and biochemical reactions using BBL Crystal GP (BD, Sparks, Maryland).
Immunohistochemistry for L. monocytogenes was performed. Briefly, paraffin-embedded intestinal tissues were sectioned at approximately 4 μm, deparaffinized, and rehydrated. The antibody was a highly purified, horseradish peroxidase (HRP)–conjugated, rabbit polyclonal IgG antibody that recognizes Listeria whole cells (L2650-09A; US Biological, Swampscott, Massachusetts). The optimal dilution of the anti-Listeria antibody was determined as 1:100 with an incubation time of 45 minutes by titration on positive control tissue sections. The anti-Listeria antibody was diluted using Dako Antibody Diluent (S0809; Dako, Carpinteria, California). Antigen retrieval consisted of Proteinase K (S3020; Dako) treatment for 5 minutes. Endogenous peroxidase was blocked using 3% hydrogen peroxide (H312-500; Fisher Scientific, Fair Lawn, New Jersey). All other blocking was completed with Power Block (HK085-5 K; Biogenex, San Ramon, California). Brain from a Listeria-infected goat was used as a positive tissue control. Purified rabbit immunoglobulins supplied as a ready-to-use reagent replaced the anti-Listeria antibody as a negative antibody control (NC495; Biocare Medical, Concord, California). A section of normal equine intestine was used as negative tissue control. The MACH 1–HRP polymer detection system (M1U539; Biocare Medical), followed by the betazoid 3,3′-diaminobenzidine chromogen kit (BDB2004; Biocare Medical), was applied for visualization of Listeria sp antigen. The tissue sections were counterstained with Gill’s II hematoxylin and bluing; dehydrated in alcohol levels of 70%, 95%, and 100%; cleared in xylene; and mounted with xylene-based mounting medium.
Results
Histologic lesions included granulomatous typhlocolitis, lymphangitis, and colic lymphadenitis. The colon and cecum had widely scattered nodules of epithelioid macrophages and multinucleated giant cells, centered about foci of liquefactive necrosis with many neutrophils and cellular debris. The nodules extended from the multifocally denuded mucosal surfaces through the submucosa and serosa. Where the nodules extended into the mesentery, they were usually arranged around dilated, focally disrupted lymphatic vessels and extended into the cortex and medulla of regional lymph nodes (Figs. 1–3). Within most of the densely cellular nodules (including those in the submucosa and lymph nodes), macrophages, multinucleated giant cells, and neutrophils contained many clusters of Gram-positive, non-acid-fast coccobacilli that were immunohistochemically positive for L. monocytogenes (Fig. 4). A few nodules of epithelioid macrophages were arranged around nematodes embedded in the mucosa or, less commonly, in the submucosa. The nematodes had an annulated cuticle, a thin hypodermis, somatic musculature, and pseudocoelom; a single layer of epithelial cells with multiple nuclei lined the intestine, which had a distinct brush border.
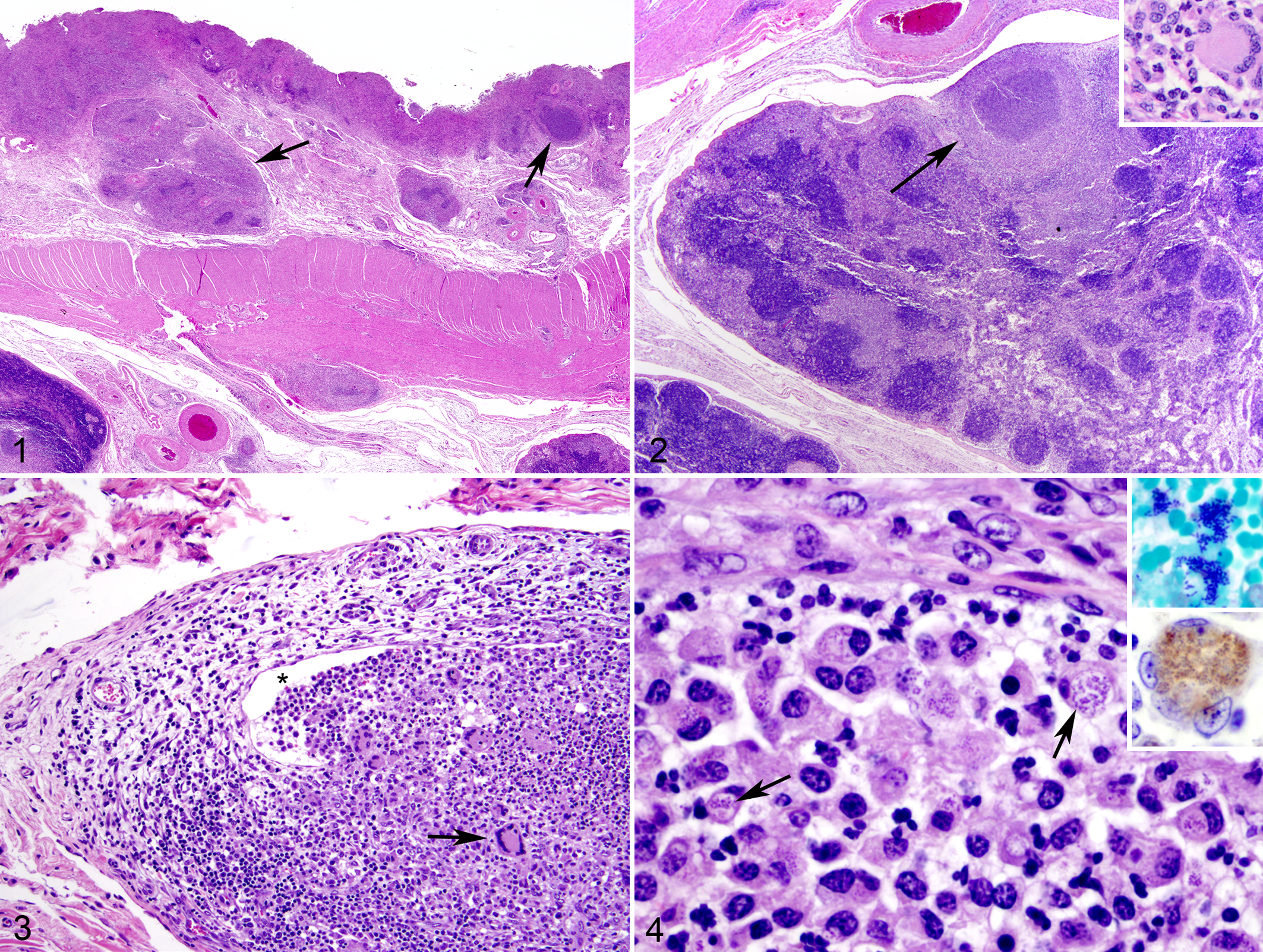
Colon; horse. The eroded mucosa is hypercellular with deep mucosal and submucosal nodules (arrows). Hematoxylin and eosin (HE).
Polymerase chain reaction assays on intestinal contents were positive for Salmonella enterica subsp enterica (S. enterica) and Listeria spp and L. monocytogenes, as well as negative for Yersinia enterocolitica. S. enterica was cultured from the intestine, and the isolate was serotyped as Typhimurium var 5. Bacterial culture from an intestinal wall nodule resulted in moderate growth of short, Gram-positive, non-acid-fast rods characteristic of L. monocytogenes.
Discussion
The histologic findings of granulomatous typhlocolitis in this horse are characteristic of an infectious disease process with multiple causes, supported by concurrent detection of intestinal L. monocytogenes, S. enterica, and nematodes (most likely cyathostomes based on microscopic features and location). However, the presence of numerous intracellular, Gram-positive coccobacilli that were immunohistochemically positive for Listeria spp within the lesions supports L. monocytogenes as the main cause of disease. Epidemiologic factors that may have contributed to the development of enteric listeriosis on this farm include a 4-year history of sporadic seasonal, watery diarrhea, previous use of the pasture for a beef cattle operation, location (northwest Georgia), age of the horse (15 years), and lack of antibiotic treatment response.
The thickened intestinal walls, mucosal nodularity, and enlarged lymph nodes are consistent with an infiltrative disease process, such as alimentary lymphoma or proliferative typhlocolitis, the latter of which is often associated with various granulomatous diseases, including chronic salmonellosis, cyathostomiasis, mycobacteriosis, and yersiniosis. Less likely entities in the differential diagnosis included fungal or algal infections. Alimentary lymphoma was ruled out based on histologic findings. Likewise, mycobacteriosis and yersiniosis were excluded on the basis of histochemistry and microbiologic results, including PCR tests. Although cyathostomes were detected histologically in the mucosa and submucosa, the numbers were few, and the nematodes were not associated with the lymphangitis or lymphadenitis.
Infection with Listeria spp has been documented in many species, but listeriosis, sometimes with associated neurologic disease or abortions, is most common in domestic ruminants. 5 Infection with L. monocytogenes in horses is rarely documented. Coinfection with Salmonella and small strongyles in the present horse probably contributed to infection with Listeria and to the development of the granulomatous typhlocolitis. Although the chronology of infections in this case is unknown, S. enterica and cyathostome infections could have shifted the immune response to L. monocytogenes infection to a cell-mediated (granulomatous) reaction in contrast to the suppurative abomasitis and enteritis described in sheep with gastrointestinal listeriosis. 3
The source of L. monocytogenes that potentially led to severe diarrhea in multiple horses over a 4-year period on the present farm remains undetermined, but possibilities include environmental contamination by wildlife reservoirs or the previously housed beef cattle, or contaminated feed. The route of infection in the current horse was probably via ingestion. It is possible that long-term persistence of L. monocytogenes in the environment, originating from the previous cattle on the pasture, is a potential source of infection for horses, as well as other animals on the farm.
In summary, although uncommon, L. monocytogenes is a potential cause of granulomatous typhlocolitis in horses. Diagnostic tools, such as immunohistochemistry, Gram’s and other bacterial stains, and PCR tests, in addition to bacterial culture, were used in the present case to aid in determination of L. monocytogenes as the major cause of enteric disease.
Footnotes
Acknowledgements
Ms Paula Bartlett, Ms Ingrid Fernandez, and Ms Sarah Bates are commended for excellent assistance with the bacterial isolation and molecular diagnostic testing.
Declaration of Conflicting Interests
The author(s) declared no conflicts of interest with respect to the authorship and/or publication of this manuscript.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The current work was supported by the Athens Veterinary Diagnostic Laboratory, College of Veterinary Medicine, University of Georgia.