
Abstract
Persistent infection (PI) with bovine viral diarrhea virus (BVDV) has been associated with osteopetrosis and other long bone lesions, most commonly characterized as transverse zones of unmodeled metaphyseal trabeculae in fetuses and calves. This study was undertaken to characterize the morphogenesis of fetal long bone lesions. Forty-six BVDV-naïve pregnant Hereford heifers of approximately 18 months of age were inoculated with noncytopathic BVDV type 2 containing media or media alone on day 75 of gestation to produce PI and control fetuses, respectively, which were collected via cesarean section on days 82, 89, 97, 192, and 245 of gestation. Radiographic and histomorphometric abnormalities were first detected on day 192, at which age PI fetal long bone metaphyses contained focal densities (4 of 7 fetuses) and multiple alternating transverse radiodense bands (3 of 7 fetuses). Day 245 fetuses were similarly affected. Histomorphometric analysis of proximal tibial metaphyses from day 192 fetuses revealed transverse zones with increased calcified cartilage core (Cg.V/BV, %) and trabecular bone (BV/TV, %) volumes in regions corresponding to radiodense bands (P < .05). Numbers of tartrate resistant acid phosphatase positive osteoclasts (N.Oc/BS, #/mm2) and bone perimeter occupied (Oc.S/BS, %) were both decreased (P < .05). Mineralizing surface (MS/BS, %), a measure of tissue level bone formation activity, was reduced in PI fetuses (P < .05). It is concluded that PI with BVDV induces cyclic abnormal trabecular modeling, which is secondary to reduced numbers of osteoclasts. The factors responsible for these temporal changes are unknown but may be related to the time required for osteoclast differentiation from precursor cells.
Keywords
Bovine viral diarrhea virus (BVDV) (genus Pestivirus, family Flaviviridae) is taxonomically divided into BVDV1 and BVDV2 genotypes, both of which contain cytopathic (cp) and noncytopathic (ncp) biotypes. 15,16,30 The majority of BVDV infections, regardless of infecting strain type, result in asymptomatic infections that largely go undetected. While acute infections can occur with either biotype, infection with a ncp strain of BVDV, regardless of genotype, appears to be a prerequisite for persistent infection (PI) of the fetus. 5,9,17
NcpBVDV, by virtue of its ability to cross the placenta and infect the fetus at any stage of gestation, has been associated with a myriad of different manifestations, including early embryonic death, abortion, fetal PI, and congenital abnormalities. 2,10,11 The fetal consequences of transplacental infection appear to be primarily determined by the age of the fetus at the time of infection, although other factors may contribute, such as the pathogenicity of the infecting virus strain and the immune and health status of the dam. 4 Transplacental infection of the fetus by ncpBVDV prior to adaptive immune system development, which occurs during the second and third trimesters of gestation, is thought to enable PI and lifelong viral shedding by the animal due to development of highly virus-specific T-cell tolerance to the infecting strain. 8 Transplacental infection of the fetus after adaptive immune system development (beyond ~ day 150) results in acute infection, fetal seroconversion to the infecting viral strain, and clearance of the virus. 10 PI animals are of particular epidemiologic significance in that they represent the reservoir of virus in the population and are responsible for perpetuation of the virus through infection of herd mates. 19
There are only a few published investigations of naturally occurring fetal and neonatal skeletal lesions associated with BVDV infection. 26,27,32 A multitude of different terms, from osteopetrosis to growth arrest lines, has been used in the literature to describe the morphologic character of skeletal lesions associated with BVDV infections. Experimentally produced fetal bone lesions were first described by Done et al as simple growth arrest lines detected by radiography, but the distribution and histologic character were not described. 10 Another experimental infection, using the same viral strain used herein, resulted in thickening of the femoral cortices in PI fetuses. 35 In one study, a majority of BVDV-positive fetuses had fetal femora with multiple transverse zones of radiodensity in the metaphysis corresponding to growth retardation lattices, focal radiodensities corresponding to areas of retained secondary spongiosa, and reduced middiaphyseal diameter. 27
Growth retardation lattices differ from simple growth arrest lines not only in morphologic character but also in the processes responsible for their development. 27,38 While both lesions occur in regionally extensive areas parallel to the physis, growth arrest lines are composed of increased bone volume with enhanced trabecular connectivity and result from impaired longitudinal growth of the physis and normal bone formation. 38 In contrast, growth retardation lattices are composed of increased calcified cartilage core volume, typically resulting from impaired resorption of calcified cartilage. 38 Nuss et al reported a case of transient benign osteopetrosis in a newborn calf PI with BVDV in which long bone metaphyses contained multiple regularly spaced, thick, transverse bands of radiodensity, which resolved radiographically by 5 months of age. 26 Although the term osteopetrosis has been used to describe skeletal lesions associated with BVDV, 26,32 it is probably more appropriately reserved for genetic diseases that result in diffuse, rather than zonal or cyclic, metaphyseal changes. In the previously mentioned case, BVDV antigen was detected by immunohistochemistry in osteoblasts, osteocytes, and other bone marrow hematopoietic cells after necropsy at 13 months. 26 Unfortunately, the presence or absence of BVDV antigen in osteoclasts was not described.
Thus, so far the reported lesions have been largely limited to the long bone metaphyses and have ranged from barely discernible growth arrest lines and growth retardation lattices to diffuse and zonal retention of secondary and
This study was undertaken to characterize the morphogenesis of long bone lesions over the course of experimentally induced fetal PI with BVDV in an attempt to determine the processes and mechanisms responsible for the development of these lesions.
Materials and Methods
Infection and General Growth Parameters
All animal experiments were approved by Colorado State University’s Institutional Animal Care and Use Committee. Forty-six yearling Hereford heifers weighing between 295 and 430 kg were artificially inseminated with semen from a single Angus bull after estrus synchronization. Pregnancy was detected on day 35 and confirmed on day 70 of gestation. All heifers were seronegative for BVDV type 1 and type 2 prior to infection. Heifers were group housed in outdoor pens. On day 75 of gestation, heifers (n = 23) were inoculated intranasally with either 2 ml of α-MEM media containing 4.4 log10TCID50 ncpBVDV type 2 strain 96b2222 39 or α-MEM media alone (n = 23) to produce PI and control fetuses respectively, as previously described. 35 Maternal infection was confirmed by detection of viral RNA in peripheral blood leukocytes on day 7 postinfection by using semiquantitative real-time polymerase chain reaction (qt-RT-PCR) as previously described 35 and by the presence of serum neutralizing anti-BVDV2 antibodies on day 28 postinfection as previously described. 31 Control heifers remained negative for viral RNA and seronegative throughout the study. Eight fetuses, 4 infected and 4 controls, were collected by cesarean section on days 75, 82, 97, 192, and 245 of gestation. Six heifers (3 infected and 3 controls) received an intravenous injection of 9 mg/kg of oxytetracycline (LA-200, Pfizer Animal Health, Exton, PA) on day 175 and again on day 185 of gestation to flurochrome label bone formation for dynamic histomorphometry. Fetuses from these heifers were removed via cesarean section on day 192. Metaphyseal elongation rates for the proximal tibia were calculated from midfrontal sections of flurochrome-labeled fetuses. These calculations were performed by measuring the length, in 4 equidistant locations, of the unlabeled spongiosae subjacent to the physis and dividing the average length by the period (7 days) between administration of the second label and collection. Body, liver, heart, lung, kidney, spleen, thymus, adrenal, and thyroid weights as well as crown–rump lengths and heart girths were recorded. One PI fetus was lost due to abortion on day 230 of gestation.
Radiography
Fetal bones were dissected free of extraneous soft tissues and digital radiographs were produced using a portable X-ray machine set (MinXray, Northbrook, IL) and a digital plate system (Elikin, Carlsbad, CA). Care was taken to precisely align limbs in the same position for these radiographs. Epiphyseal and middiaphyseal widths and physeal-to-physeal length of the femur, tibia, humerus, and radius were obtained from the radiographs, which contained an internal measurement standard, using Imagepro software (MediaCybernetics, Bethesda, MD). Measured parameters that differed between PI and control fetuses were confirmed by manual measurement with dial calipers.
Histology
Bones were dissected free of soft tissues, fixed in 10% buffered formalin, and cut on an ISOMET bone saw (Beuhler, Ltd, Lake Bluff, IL) to yield anatomically standardized midfrontal sections of the proximal tibial metaphysis. The sections were decalcified in 5% formic acid (Sigma, St Louis, MO) over a period of 3–7 days depending on the fetal age, before being routinely processed and paraffin embedded. Five-micrometer-thick sections were prepared on a rotary retracting microtome and stained with hematoxylin and eosin. Serial 5-μm-thick sections were incubated in 0.2 M Tris-HCL, pH 9.0, solution at 37°C for 30 minutes to reactivate acid phosphatases, 25 then stained with a commercial kit (Sigma) to detect cells expressing tartrate-resistant acid phosphatase (TRAP). In addition to quantitative histology this approach facilitated qualitative evaluation of osteoclasts for evidence of active resorption in Howship lacunae and number of nuclei per osteoclast as well as signs of degeneration or apoptosis.
Immunohistochemistry was performed on formalin-fixed, nondecalcified, paraffin-embedded sections to avoid potential adverse effects of the decalcifying agent on viral antigen and to exactly duplicate the tissue processing utilized for positive and negative control tissues. Spleen and thymus from day 192 control and PI fetuses served as negative and positive control tissues. The combined effect of approximately 1-cm2 samples of metaphyseal bone and the low degree of mineralization of fetal bone at this location facilitated the production of high-quality histologic sections using routine paraffin embedding and sectioning on a rotary microtome utilizing disposable blades. The mouse monoclonal anti-BVDV clone 15C5 (IDEXX Laboratories, Westbrook, ME) was used as previously described, 14 and staining was visualized with a Vector Elite Kit (Vector Laboratories, Burlingame, CA) per manufacturer’s recommendations. Antigen retrieval was performed with 20 μg/ml of Proteinase K (Promega, Madison, WI).
Formalin-fixed facing sections of tibia that were produced during the initial bone sectioning were embedded in glycol methacrylate as previously described and sectioned on a bone saw (Maruto, Tokyo, Japan). 20 The resulting nondecalcified semithin sections were ground down to 30-μm thickness for assessment of formation parameters. Femoral middiaphyseal cross sections were prepared by embedding the sample in Sample-Kwick (Beuhler, Ltd) and sectioning on an IsoMet saw (Beuhler). The resulting sections were ground down to 100-μm thickness.
Histomorphometry
Tibial sections were divided into 4 zones of interest as illustrated in Figure 1. The zones were designed such that the subphyseal zone would entirely encompass the primary spongiosa, which was found to not exceed 2 mm in length. The subjacent 3 zones were then determined by dividing the remaining metaphysis equally in thirds. Histomorphometric analysis of static parameters was performed manually using a modified Mertz grid reticle at a standardized magnification (200×) in a tissue area of 4 mm2 from the central portion of each zone. Histomorphometric analysis of dynamic parameters was performed on a Nikon E 800 fluorescent microscope (Melville, NY) equipped with an ultraviolet filter, using Imagepro software, in a 15-mm2 area of interest from the central portion of middle and distal zones. The subphyseal and proximal zones were not analyzed for dynamic parameters due to the complete lack of labeling and predominance of diffuse labeling, respectively. The definition of the parameters evaluated, their corresponding referents, standardized nomenclature, units of measurement, and criteria for evaluation, where applicable, are listed as follows: bone volume—trabecular bone volume including calcified cartilage cores normalized to tissue volume (BV/TV, %); cartilage volume—longitudinal calcified cartilage core volume normalized to bone volume (Cg.V/BV, %); total bone surface (BS, mm2); number of osteoclasts (TRAP positive) per mm2 bone surface (N.Oc/BS, #/mm2); osteoclast surface—TRAP-positive osteoclast-covered bone surface normalized to bone surface (Oc.S/BS, %); single-labeled surface—area of bone surface with a single flurochrome label normalized to bone surface (s.LS/BS, %); double-labeled surface—area of bone surface with 2 labels normalized to bone surface (d.LS/BS, %); mineralizing surface—equal to half the area of single-labeled surface plus the area of double-labeled surface normalized to bone surface (MS/BS, %); mineral apposition rate—average distance between first and second label divided by the number of days in the interlabel period (MAR, μm/day); and bone formation rate—equal to the mineralizing surface normalized to bone surface multiplied by the mineral apposition rate (MS/BS * MAR, mm3/mm2/day). Femoral middiaphyseal apposition rates were determined by averaging the width of nonlabeled subperiosteal plexiform bone at 5 standardized sites and dividing this distance by the period (7 days) between administration of the second label and collection. Hematopoietic cell density was determined by measuring the proportion of marrow space occupied by such cells. Proximal tibia physeal histomorphometry was performed on decalcified day 192 section to determine whether there were differences between PI and control fetuses in the volume and number of calcified cartilage cores being incorporated into primary spongiosae. The areas occupied by chondrocyte lacunae and by matrix in the hypertrophic zone were determined in a 4-mm2 area and expressed as a ratio. The number of chondrocyte columns per transverse mm of physeal width was determined along the central most 1 cm aspect of the physis. All bone histomorphometry nomenclature, units, and calculations are consistent with current recommendation for standardization. 28
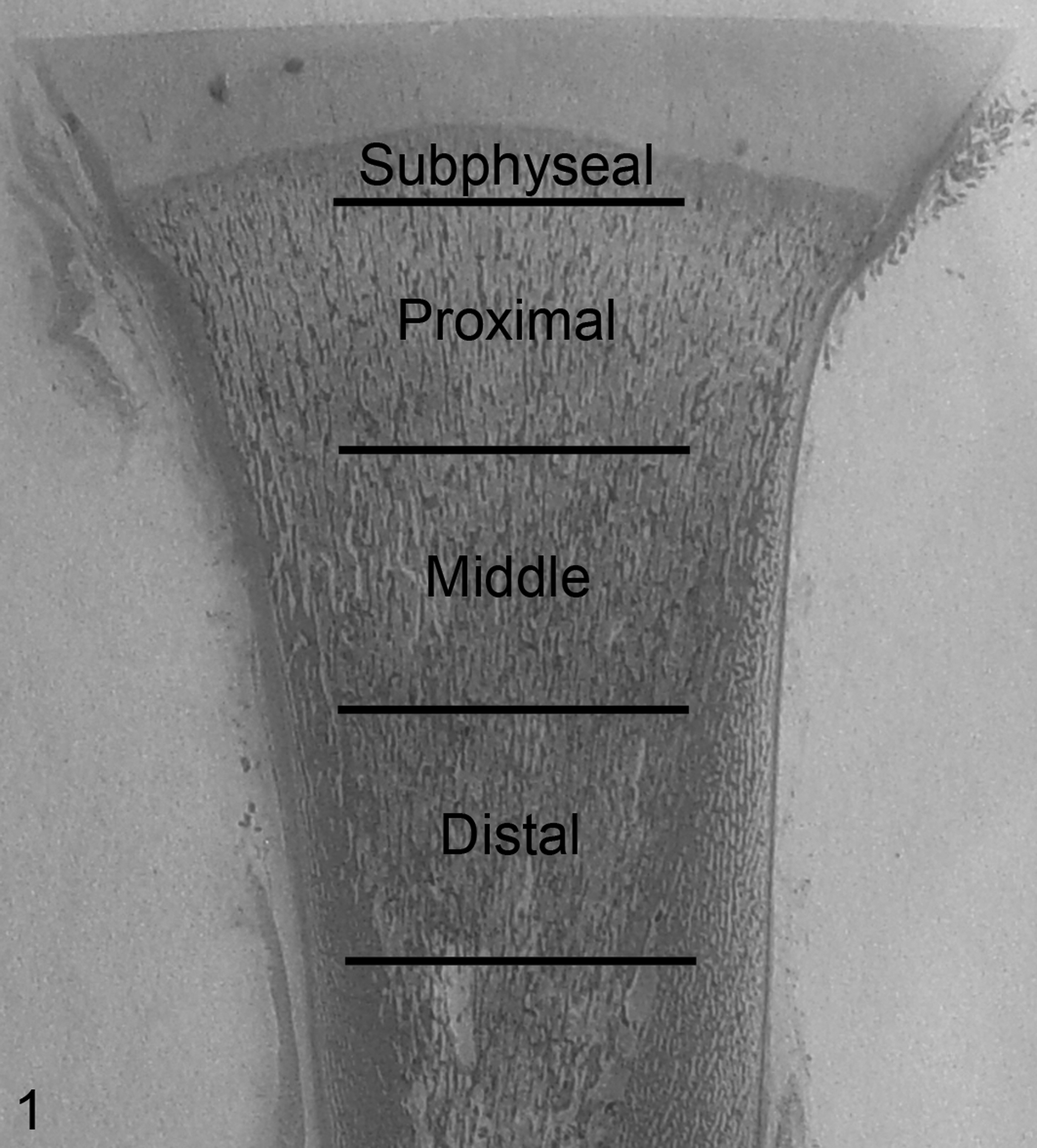
Proximal tibial metaphysis; bovine fetus, 192 days of gestation. Subdivision of the proximal tibial metaphysis into zones for histomorphometry. The subphyseal zone extends 2 mm distal from the physeal–metaphyseal junction. All other zones are 7 mm in proximal to distal length. (HE).
Statistical Analysis
Data were compared using Student t test or Mann-Whitney test depending on the result of Shapiro-Wilk test for normality. Differences at P < .05 were considered significant, while differences at P < .10 are described as tendencies. Data are presented as mean ± standard deviation.
Results
Infection and General Growth Parameters
Fetal PI was confirmed by qt-RT-PCR for BVDV RNA and immunohistochemical staining for BVDV antigen (Table 1). Fetuses from the PI group on day 82 as well as control fetuses from all collection days were negative for BVDV RNA and antigen. Two of 4 PI fetuses on day 89 were positive for BVDV antigen. However, 1 PI fetus tested positive for BVDV RNA by qt-RT-PCR (due to the technical difficulties in sample collection, blood was obtained from only 1 of 4 PI fetuses on day 89). All PI fetuses from days 97 to 245 were BVDV positive by both methods.
Presence or Absence of Radiographic Lesions Within the Tibial Metaphyses and Infection Status of PI Fetusesa

aDay 82 fetuses from PI group as well as control fetuses from all time points were negative for virus. Controls did not have any radiographic lesions. PI, persistent infection; qt-RT-PCR, semiquantitative real-time polymerase chain reaction; BVDV, bovine viral diarrhea virus.
bqt-RT-PCR for BVDV RNA in umbilical cord blood and BVDV antigen in brain as previously described. 3
cUmbilical cord blood was only successfully obtained from 1 fetus at this time point.
Crown–rump length, weight, heart girth, and major organ weights did not differ between PI and control fetuses on any of the collection days (data not shown). The fetal sex distribution by infection status was as follows: day 192, control, 3 male, 4 female; day 192, PI, 2 male, 5 female; day 245, control, 2 male, 2 female; day 245, PI, 3 male, 0 female. There was no statistically significant effect of fetal sex on any of the study parameters reported from day 192 fetuses. The effect of sex on study parameters for day 245 could not be determined due to the lack of female fetuses in the PI group. On days 192 and 245 there was an enhanced waisting or fluting of the metaphysis in PI femora, characterized by similar physeal widths and reduced middiaphyseal outer diameter compared to controls (Table 2). This indicated that lateral appositional growth of the physis was not significantly impaired by PI. In middiaphyseal cross sections, neither cortical thickness nor total cortical area divided by cortical diameter differed between the groups (Table 2). Day 192 PI fetuses had slower rates of periosteal apposition at this site than control fetuses, 23.6 μm/day ± 4.7 versus 35.9 μm/day ± 4.9 respectively.
Femoral Dimensions From Control and PI Fetusesa
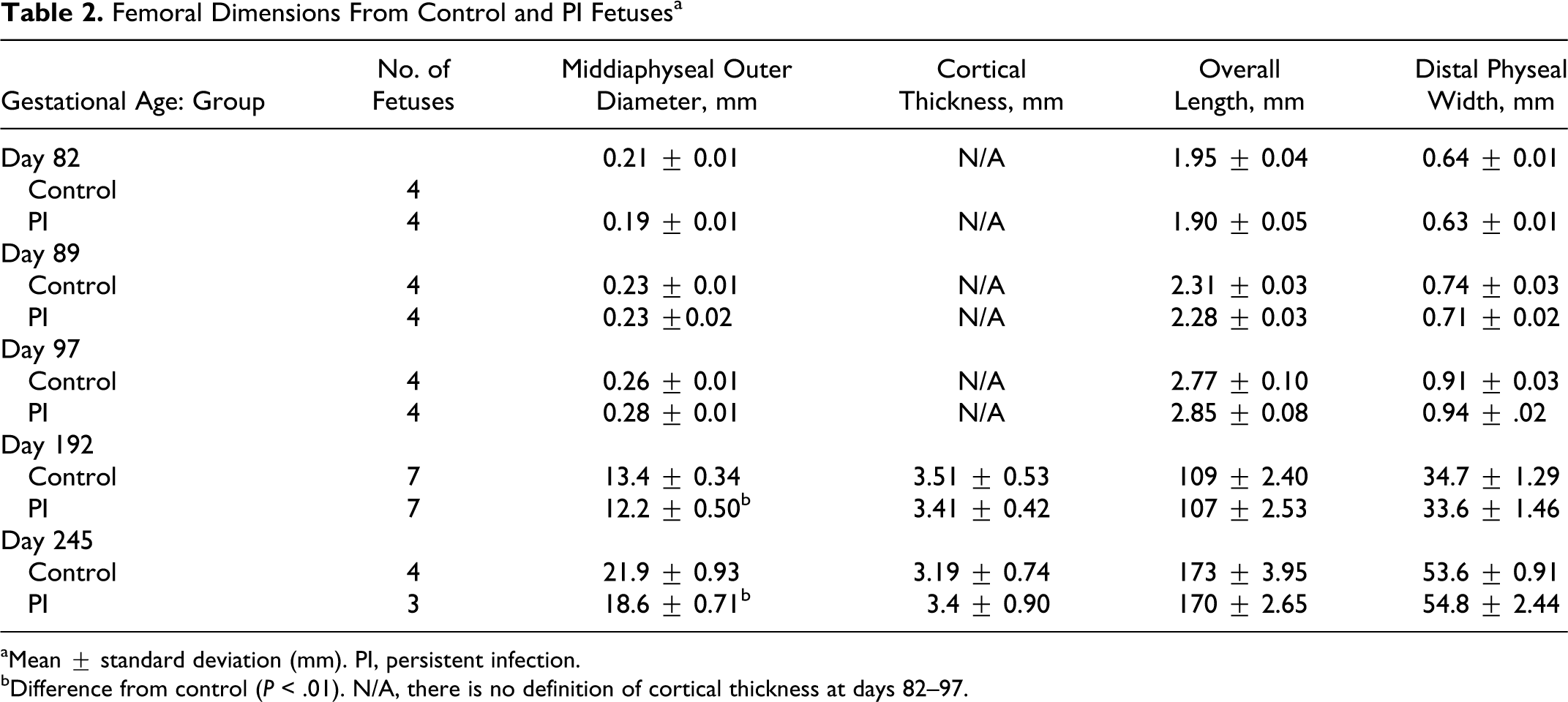
aMean ± standard deviation (mm). PI, persistent infection.
bDifference from control (P < .01). N/A, there is no definition of cortical thickness at days 82–97.
Radiography
Radiographs of the axial skeleton, skull, and limbs were taken at each time of collection. No radiographic abnormalities were detected prior to day 192, after which they were limited to the long bones of the appendicular skeleton. Relative development of secondary and tertiary centers of ossification did not differ between PI and control fetuses. Radiographs of 6 out of 7 day 192 PI fetuses contained metaphyseal lesions. These consisted of focal densities in 4 out of 7 fetuses and multiple transverse radiodense bands in 3 of 7 fetuses (Figs. 2, 3). Within the metaphysis, the distribution of the focal densities appears to be random, although they were most frequently identified within the distal femur and proximal tibia metaphyses.
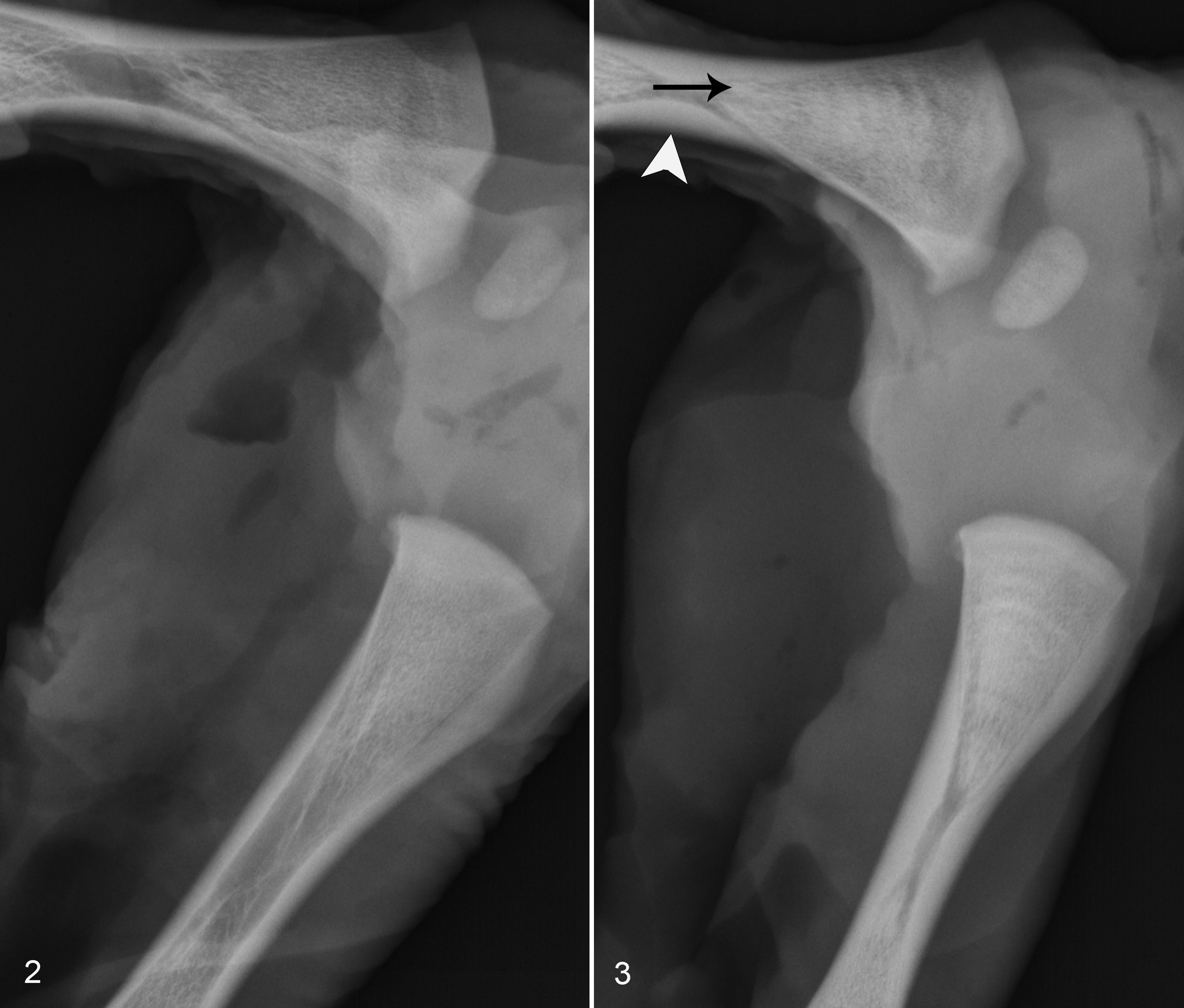
Radiographs of day 245 fetal metaphyses had similar bands of alternating radiodensity in 2 of 3 PI fetuses (not shown). There was marked variation among PI fetuses in lesion severity, which ranged from lesions easily appreciable to barely discernible lines. The distribution of such lesions between individuals differed very little. The most commonly affected metaphyses were distal radius, proximal tibia, distal femur, and proximal metacarpal/metatarsal. Radiographic lesions were not observed in the scapula or phalanges.
Metaphyseal elongation rates from day 185 to day 192 did not differ between PI (250 ± 30 μm/day) and control (310 ± 50 μm/day) fetuses and were utilized to approximate the periods between the transverse bands in fetuses, which had easily discernable transverse bands as shown in Figure 3. Acknowledging the nonlinear nature of long bone growth during the period of lesion development (days 97–192), bands of radiodensity were on average approximately 3 to 4 mm in width and would reflect an approximately 15-day period of growth separated by periods of 15 days or greater between the bands.
Histology and Histomorphometry
Histologic lesions and histomorphometric abnormalities were not detected prior to day 192, although day 97 PI fetuses had similar bone and calcified cartilage volumes and they tended to have fewer TRAP-positive osteoclasts (N.Oc/BS, #/mm2) within the proximal tibia metaphysis, but this was not statistically significant (data not shown, P < .10). Overall, the histologic lesions in PI fetuses were subtle and characterized by regionally extensive transverse areas of increased trabecular bone volume with increased amounts of longitudinal cartilage core volume and focally extensive areas of increased transverse connectivity between trabeculae (Figs. 4 –7). The former feature radiographically corresponded to the radiodense transverse bands and the latter to the focal radiodensities. There was no difference in hematopoietic cell density between PI and control fetuses (data not shown). Immunohistochemical staining for BVDV in day 192 fetal bone sections failed to demonstrate any viral antigen within osteoclasts or bone forming cells (data not shown).
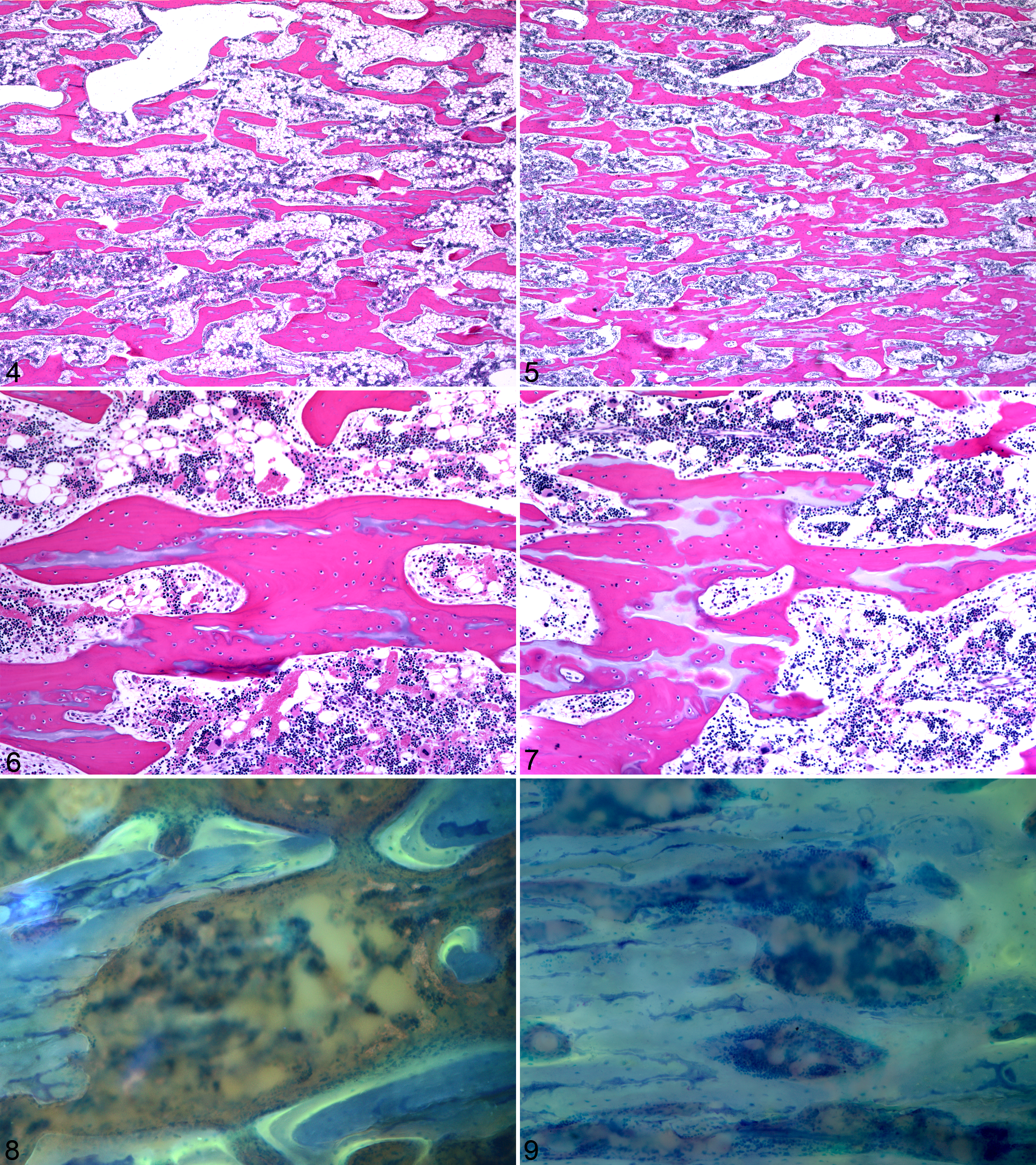
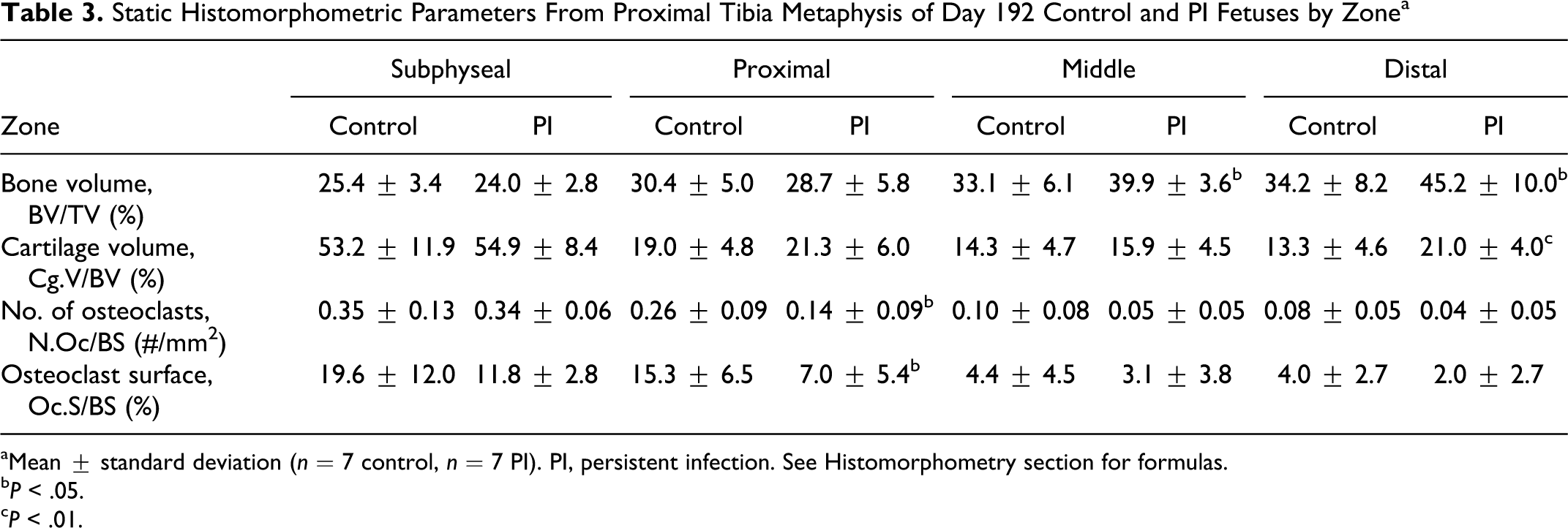
Histomorphometric parameters from the proximal tibial metaphyses of day 192 fetuses are detailed by zone in Table 3. Analysis by zone demonstrated increased bone volume within the middle and distal zones of PI tibia as well as increased cartilage core volume within the distal zone. Within the proximal zone, there were decreased numbers of TRAP-positive osteoclasts and reduced TRAP-positive osteoclast surface (OC.S/BS, %) in PI tibia.
Static Histomorphometric Parameters From Proximal Tibia Metaphysis of Day 192 Control and PI Fetuses by Zonea
aMean ± standard deviation (n = 7 control, n = 7 PI). PI, persistent infection. See Histomorphometry section for formulas.
b P < .05.
c P < .01.
Collectively, when all zones were pooled and evaluated as one zone of interest, PI tibia had increased bone volume and cartilage core volume with decreased number of TRAP-positive osteoclasts and TRAP positive-osteoclast surfaces. Qualitatively, osteoclasts from PI tibia were present within deep Howship lacunae and on crenated surfaces and did not differ in morphologic character or number of nuclei per osteoclast to those from control tibia. Histologic features of degeneration, necrosis, and apoptosis were not identified in osteoclasts or hematopoietic cells at any time point evaluated. In the evaluation of the physis at its junction with the metaphysis, there were no differences in the mean number of chondrocyte columns per transverse millimeter of physeal–metaphyseal junction, 24.4 ± 1.5 versus 23.7 ± 1.0, control versus PI, respectively, or ratios between chondrocyte lacunar area and matrix area, 1.6 ± 0.4 versus 1.3 ± 0.3, between control and PI fetuses.
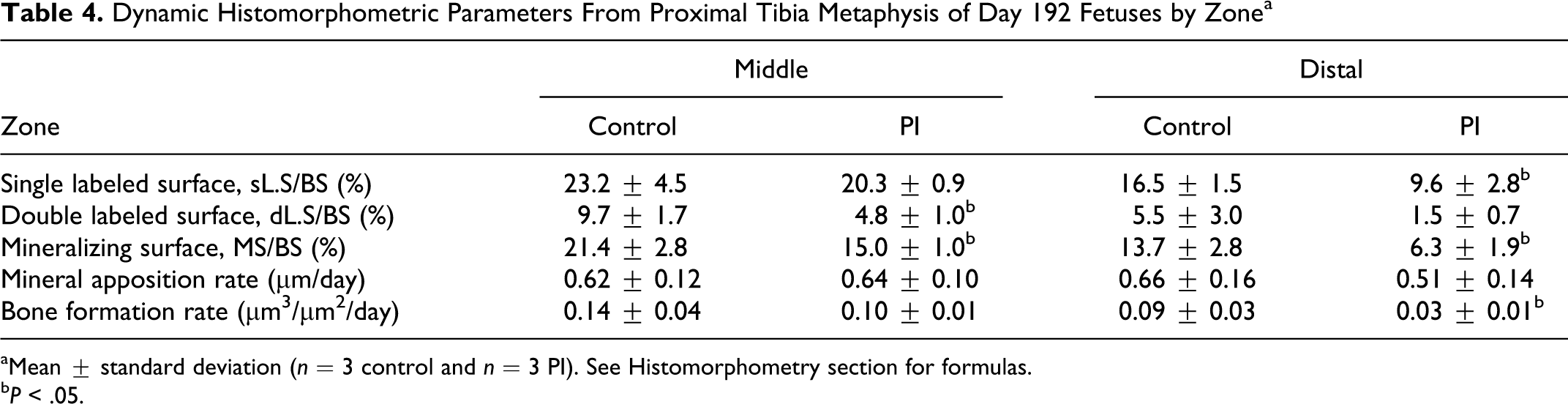
Bone formation parameters in the middle and distal zones from the proximal tibial metaphyses of day 192 fetuses are shown in Table 4. The majority of flurochrome-labeled bone surface was diffusely labeled, but a considerable amount of discrete single and double labels was present to facilitate analysis (Figs. 8, 9). Single-labeled surface was less within the distal zone as was double-labeled surface within the middle zone of PI tibias. Mineralizing surface was less in PI tibia within both zones. There was no difference in mineral apposition rates in either zone. However, bone formation rates were slower within the distal zone of PI tibia. Collectively, when the middle and distal zones were pooled and evaluated as one zone of interest, there was decreased single-labeled surface, double-labeled surface, and mineralizing surface in PI fetuses when compared to control fetuses. Bone formation rates tended to be slower in PI tibia (P < .10). There was no difference in mineral apposition rates between the groups.
Dynamic Histomorphometric Parameters From Proximal Tibia Metaphysis of Day 192 Fetuses by Zonea
aMean ± standard deviation (n = 3 control and n = 3 PI). See Histomorphometry section for formulas.
b P < .05.
Discussion
The 2 major findings that emerged from this study were the negative impact of PI on tissue-level bone formation in trabecular and cortical bone as well as on trabecular modeling. The histomorphometric parameters correlated well with the radiographic findings. The majority of radiodense transverse bands were present within the middle and distal zones, where increased bone and cartilage core area were found. The increased radiodensity of these bands was associated with increased volumes of calcified cartilage and bone, the former contributing to the radiographic appearance of the bands due to its greater radiodensity relative to immature bone. 13
For greater cartilage core volume to be present distally in the metaphysis, cartilage resorption must have been impaired during the initial modeling of these trabeculae. It is hypothesized that in the primary spongiosa, a lesser amount of residual intercolumnar cartilage was resorbed before osteoid formation commenced. The alternative possibility is that the increased volume results directly from an increased volume of intercolumnar matrix with a lower lacunar: matrix ratio and less chondrocyte column density in the physis. The physeal morphometry results support the former hypothesis, as there was no difference in these parameters between PI and control fetuses at the time of sample collection. The periodicity of transverse bands certainly suggests a cyclic insult or physiologic alteration that manifests through reduced trabecular modeling of primary spongiosa. The transverse bands reflect a period of approximately 15 days of “normal” modeling in between periods of approximately 15 days of abnormal modeling. This period is based on our estimate of metaphyseal elongation rates and the assumption that they are not altered by this process. The reason that trabecular modeling resumes normally in between these insults is not known. However, the cyclic nature of the lesions significantly enhances the difficulty of studying the morphogenesis of these lesions, as lesions did not appear within the same region or occur during the same developmental time within all fetuses. For example, a specific metaphyseal zone from a particular PI fetus may contain no bands, when the same zone of another PI contains 1 or 2.
Metaphyseal lesions have been described in association with a variety of different viral infections in growing animals. Growth retardation lattices similar to those found in this study have been described in growing dogs infected with canine distemper virus, 1 in cats infected with feline herpesvirus, 18 in sheep infected with border disease virus, 6 and in pigs that survive acute infections with classical swine fever virus. 38 In contrast to the lesions described in this study, the growth retardation lattices caused by viral infections in other species are typically singular and lack the cyclic nature characterized herein. The pathogenesis of metaphyseal lesions caused by other pestiviruses, border disease virus, and classical swine fever virus may be similar to those described in this study, while growth retardation lattices due to feline herpesvirus and canine distemper virus infections appear to be due to the direct effect of these viruses on osteoclasts, resulting in osteoclast necrosis during the acute phase of infection. 1,18
Abnormal trabecular modeling can result from altered osteoclast production, function, migration, and survival, all of which can manifest as similar histomorphometric findings. The morphologic changes do not appear to be due to decreased osteoclast survival, as evidenced by the lack of osteoclast apoptosis or necrosis and similar numbers of osteoclast nuclei per osteoclast at all study time points. Osteoclast function appears to be normal in PI fetuses with osteoclasts being found on crenated surface and deep within Howship lacunae. Likewise, osteoclast migration did not appear to be altered, as osteoclasts were present in similar numbers at the leading edge of vascular growth at the physis. Regardless of these observations, the cyclic nature of these lesions would necessitate evaluation of samples from additional time points in the period between days 97 and 192, as well as application of specific assays to determine the cause with any certainty. Reduced numbers of osteoclasts found in the proximal zone of PI tibias are presumably responsible for the abnormal modeling, but the reason for this is not clear. Impaired osteoclast differentiation is a possible explanation, and investigation of this process may aid in explaining the time course of the cyclic nature of these lesions. It has been well established for some time that the osteoclasts are derived from bone marrow precursor cells of the monocytic linage. 22,23,34 Although the time required for generation of functional osteoclasts from permissive precursor cells in the bovine fetus is unknown, estimates from in vitro experiments in other species suggest that it ranges from 14 to 21 days for humans and approximately 11 days for mice. 12,33 This time course of development would appear to correlate with the estimated period of 15 days determined in this study.
PI with BVDV may impair osteoclast differentiation either due to a direct effect of viral infection on precursor cells or as a secondary effect as a result of the action of cytokines. Monocytes infected in vitro with BVDV have been shown to release an IL-1 inhibitor. 21 Inhibition of IL-1 activity, which positively regulates osteoclastogenesis, 24 has been postulated to be responsible for impaired osteoclast differentiation in PI animals. 32 PI fetuses have been shown to have a chronic moderate type I interferon response, 35 and it is plausible that chronic production of interferon β, a potent negative regulator of osteoclast differentiation, 37 negatively affects osteoclastogenesis.
BVDV is known to infect cells of the innate immune system, including monocytes, macrophages, and dendritic cells. 36 Recent studies have demonstrated that BVDV-infected monocyte-derived macrophages have significantly impaired functional capacity, as measured by phagocytosis and cytokine production in vitro. 7,29 Because macrophages and osteoclasts originate from a common precursor cell, both cell types may be affected by a similar mechanism. Similar to the findings of others, 26,32 attempts to demonstrate BVDV antigen in osteoclasts by immunohistochemical methods were not successful, even though positive control tissues from other organs that were processed in the same manner as the bone sections resulted in positive staining. The reason for this is unclear but may be due to the amount of virus present being below the threshold of detection or absence of viral antigen altogether within osteoclasts.
The second important finding was reduced tissue level bone formation in PI fetuses suggesting a decrease in the extent of osteoblast surface actively involved in bone formation. The cell-level parameter of bone formation, mineral apposition rate, appeared to be normal, suggesting that PI impairs recruitment of osteoblasts into active formation or there is cyclic impairment of cell-level bone formation not apparent in the intralabeling period (days 175–185). Other investigators have reported the presence of BVDV antigen in osteoblasts and osteocytes, 26 but this was not corroborated in present study. Thus, the potential primary effect of ncpBVDV on bone-forming cells is uncertain. However, BVDV infection of osteoblasts could affect osteoclastogenesis indirectly through impaired receptor activator for nuclear factor kappa B (RANK) signaling and provide an alternative explanation for the observed effects on both cell types.
One common theme throughout this study was the marked individual variation in parameters measured and lesion severity. Certainly, one of the most important contributing factors was the cyclic nature of the lesions, which inevitably resulted in some PI fetuses demonstrating features of normal modeling while other PI fetuses demonstrated those of abnormal modeling within the same anatomic region of the metaphysis. It would appear that this reflects inherent individual biological variation or possibly a complex combination of undetermined factors that may have further contributed to the variation in the degree of lesion severity. This was a consideration in focusing the study on the day 192 fetuses because they possessed the most severe radiographic lesions and a superior sample size when compared to other collection times.
In summary, PI of bovine fetuses with ncpBVDV results in decreased numbers of osteoclasts, which impair trabecular bone modeling in a cyclic fashion. Experiments aimed at clarifying the mechanisms responsible for decreased osteoclast numbers in PI are underway. Additionally, PI impairs tissue-level bone formation activity in the trabecular and cortical envelopes. These results may provide one explanation for the growth abnormalities observed in PI cattle. To our knowledge this study provides novel insight to the time of occurrence of long bone lesion during development in PI, the variable presence and severity of such lesions, and their cyclic nature with an estimation of the duration of the period of abnormal modeling. It is also the first study to demonstrate the successful use of in vivo flurochrome labeling of fetal bone, via maternal administration, to document impaired bone formation and periosteal apposition.
Footnotes
Acknowledgements
We thank Drs. Luiz Henkes and Ryan Ashley for assistance with the fetal collections and Rebecca Setterberg for preparing calcified bone sections.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This work was supported by National Research Initiative Competitive Grant No. 2008-35204-04652 to TH and No. 2011-67012-30705 to BW from the USDA National Institute of Food and Agriculture; and Colorado State University, College of Veterinary Medicine and Biomedical Sciences, College Research Council award to NS and BW. BW was supported by NIH 5T32RR007072-10.