
Abstract
Persistent infection following in utero exposure to bovine viral diarrhea virus (BVDV) early in gestation is a serious cause of morbidity and mortality in cattle industries worldwide. The brain is a primary target of persistent infection. In the current study, the types of cells infected and topography of viral antigen expression were examined in brain sections from 9 BVDV persistently infected crossbred calves, all less than 1 year of age, by immunohistochemical staining using the 15C5 primary monoclonal antibody. BVDV antigen was detected in the brains of all persistently infected calves. A variety of cell types was infected, including neurons, astrocytes, oligodendroglia, blood vessel-associated cells (pericytes, perivascular macrophages, smooth muscle cells), and cells in the leptomeninges (blood vessel-associated cells). Conclusive demonstration of viral antigen in vascular endothelial cells was elusive. The intensity and distribution of viral antigen staining in neurons were highly variable. Viral antigen staining was most consistent and intense in thalamic nuclei, most notably in dorsal and medial nuclear groups, followed by the hippocampus, entorhinal cortex, basal nuclei, and piriform cortex. Staining in other brain areas was often less intense and inconsistent. The variability in the intensity and topography of viral antigen in the brain may explain the heterogeneity in the clinical manifestations of BVDV-induced disease. Additionally, infection of the brain in persistently infected calves may underlie or at least contribute to endocrine disturbances and immunologic deficits that are protean manifestations of BVDV-induced disease.
Bovine viral diarrhea virus (BVDV) is a Pestivirus in the Flaviviridae family. Two major serotypes, types 1 and 2, are recognized. In both serotypes are 2 biotypes, noncytopathic (ncpBVDV) and cytopathic, based on effects in cell culture. BVDV infection is a cause of significant morbidity and mortality in cattle industries and results in serious economic loss. 41, 48 Fetal infection prior to development of immunocompetence (roughly 120 days gestation) results in persistent infection. 15, 77 Calves infected with BVDV in utero and persistently infected serve as a source of infection for other in-contact cattle. 41, 48 Infection of naïve cattle during the appropriate stage of gestation perpetuates the persistently infected state.
Postnatal clinical symptoms in BVDV persistently infected calves are protean. Some calves show no appreciable signs. 64 Others, however, suffer from disturbances in growth, reproduction, and/or immunosuppression, the latter potentially leading to death due to infectious disease early in life. 41, 48, 77 Considerable effort has been expended to better understand the mechanistic basis for these abnormalities in affected calves. In persistently infected calves, viral tropism for a variety of tissues has been documented. Studies have shown evidence of viral infection in target tissues involved in growth and reproduction in persistently infected calves (i.e., reproductive and endocrine organs). 11, 27, 35, 80 Other studies have alluded to specific defects in the immune system in both persistently and acutely infected calves that could contribute to viral persistence and/or immunosuppression. 12, 15, 17, 31, 32, 61, 62, 64 In vitro investigations of target cells in the immune system (i.e., leukocytes and parameters of leukocyte function) have also indicated alterations. 1, 2, 4, 5, 12, 14, 17, 33, 34, 44, 61, 62, 67, 71, 84 Such studies have been valuable, but there is not total agreement between studies with regard to defects in the immune system. Additionally, in vitro studies may not reflect the true in vivo situation. 16 These studies have, however, been used as the basis for elucidating the pathogenesis of BVDV-induced disease. To date, a comprehensive understanding of BVDV persistent infection relative to the clinical manifestations of disease has been elusive. 28, 65, 77
The utility of immunohistochemical (IHC) staining for BVDV antigen has been demonstrated in numerous studies of both persistent as well as acute BVDV infection. 7, 11, 19, 20, 26, 27, 29, 30, 35– 38, 46, 47, 51, 56, 76, 78, 80, 86 Staining for BVDV antigen serves as one of the major means of detecting BVDV persistently infected calves in surveillance efforts. 20, 56 IHC studies have also indicated that the brain is a primary target of fetal infection. 26, 38, 86 Complex interactions occur between the nervous, endocrine, and immune systems during fetal development and later in life, abnormalities potentially impacting growth, reproduction, and immune responsiveness. 8, 10, 24, 25, 53, 63, 66, 68 These interactions have not been adequately evaluated in studies of BVDV persistently infected calves. The purpose of the current study was to re-examine the infected cell types, topography, and intensity of staining for BVDV antigen in the brains of persistently infected calves with regard to the potential impact on nervous-endocrine-immune interactions. Although IHC staining for BVDV antigen in the brains of persistently infected calves, per se, would not specifically indicate functional deficits, the cellular heterogeneity, topography, and intensity of viral antigen expression in the brain of calves could suggest directions for more in-depth evaluation.
Materials and Methods
Animals
Nine BVDV persistently infected calves, identified by routine surveillance testing based on IHC staining of ear notches, were purchased from ranches in Wyoming (n = 3) and Nebraska (2 groups of n = 4 and n = 2). The calves were crossbreds of beef breeds. The owners of the calves reported no apparent health problems in any of the calves prior to purchase. Two BVDV-negative calves served as controls. One was a herd mate to persistently infected calves purchased from one of the ranches in Nebraska, and the other was born and raised at the livestock facility of the Veterinary Sciences Department. The source of the calves and other pertinent data, including isolation of types 1 and 2 ncpBVD viruses from blood samples collected prior to euthanasia, are given (Table 1). Although the ages of all calves were not available from the owners, all were born during the late fall/winter calving season of 2004–2005 and were less than 8 months of age at the time of necropsy. The calves, purchased from outside sources, were transported to the Veterinary Sciences Department and euthanized within 24 hours of arrival. All handling of the calves was conducted according to Institutional Animal Care and Use Committee guidelines and approvals. All calves were sedated with xylazine and euthanized with an overdose of barbiturates.
Calves included in the study.
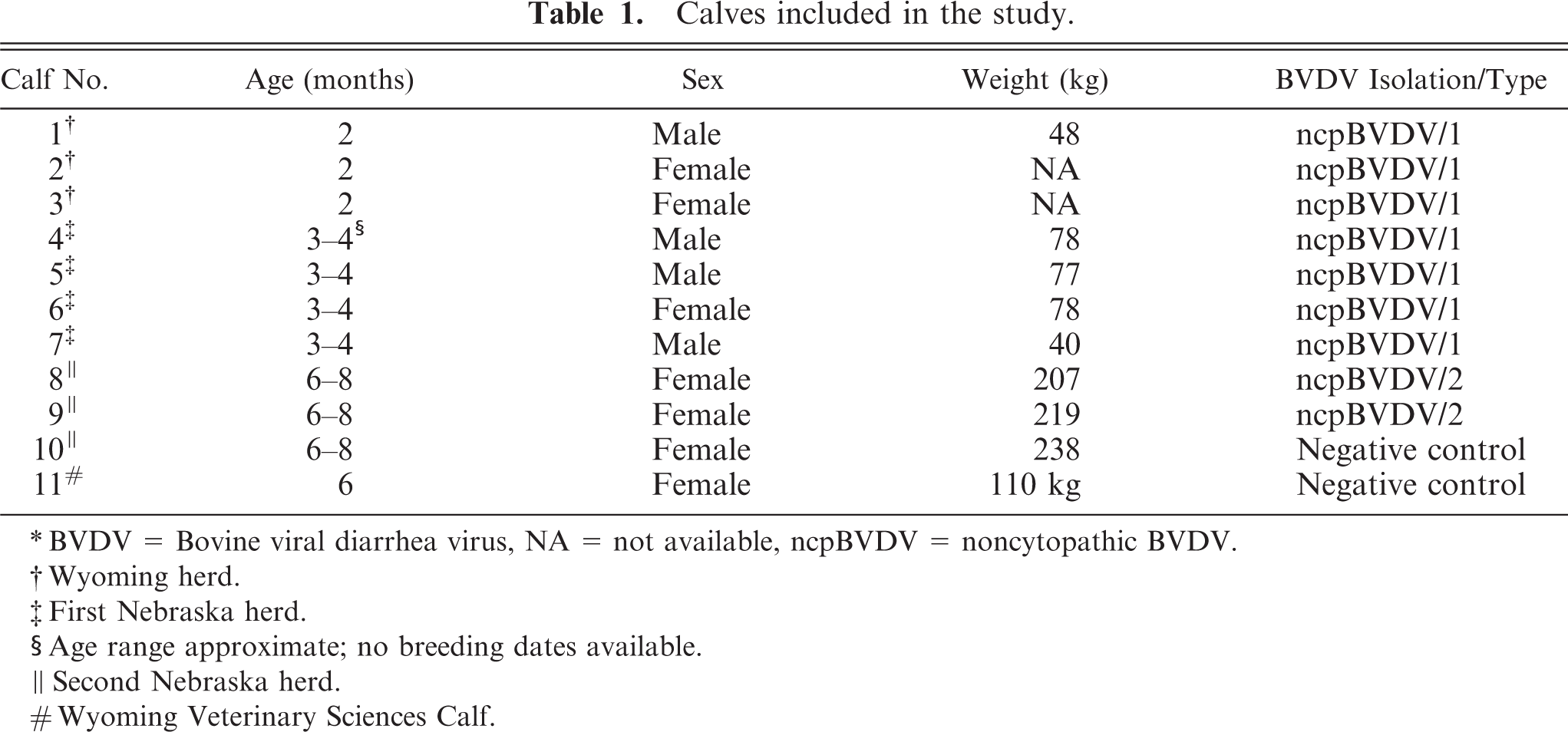
∗ BVDV = Bovine viral diarrhea virus, NA = not available, ncpBVDV = noncytopathic BVDV.
† Wyoming herd.
‡ First Nebraska herd.
§ Age range approximate; no breeding dates available.
∥ Second Nebraska herd.
# Wyoming Veterinary Sciences Calf.
Necropsy and tissue collection
No signs of clinical illness were detected in any of the calves during examination prior to necropsy. One calf from a Nebraska herd (calf No. 7) had a considerably lower body weight than persistently infected herd mates (Table 1). Complete necropsies on all calves were performed immediately following euthanasia. No gross lesions were detected. For this study, intact brains were removed and immersed in 10% neutral buffered formalin and placed on an orbital shaker. Following a 7-day period of fixation, brains were trimmed for histologic examination. From 17 to 21 coronal and hemicoronal sections of brain, representing major neuroanatomic areas, as listed inTable 3, were processed for paraffin embedding, sectioning, and staining, according to accepted histologic technique. Sections were taken to allow bilateral examination of basal nuclei and brain stem structures. An additional sagittal section of the cerebellum was also included. All sections were stained routinely with HE for detection of any relevant microscopic lesions and for more precise neuroanatomic comparisons with IHC staining.
Grading criteria for intensity of neuronal staining for BVDV antigen in persistently infected calves.

∗ BVDV = Bovine viral diarrhea virus.
Intensity of BVDV antigen staining in major brain regions of persistently infected calves.∗
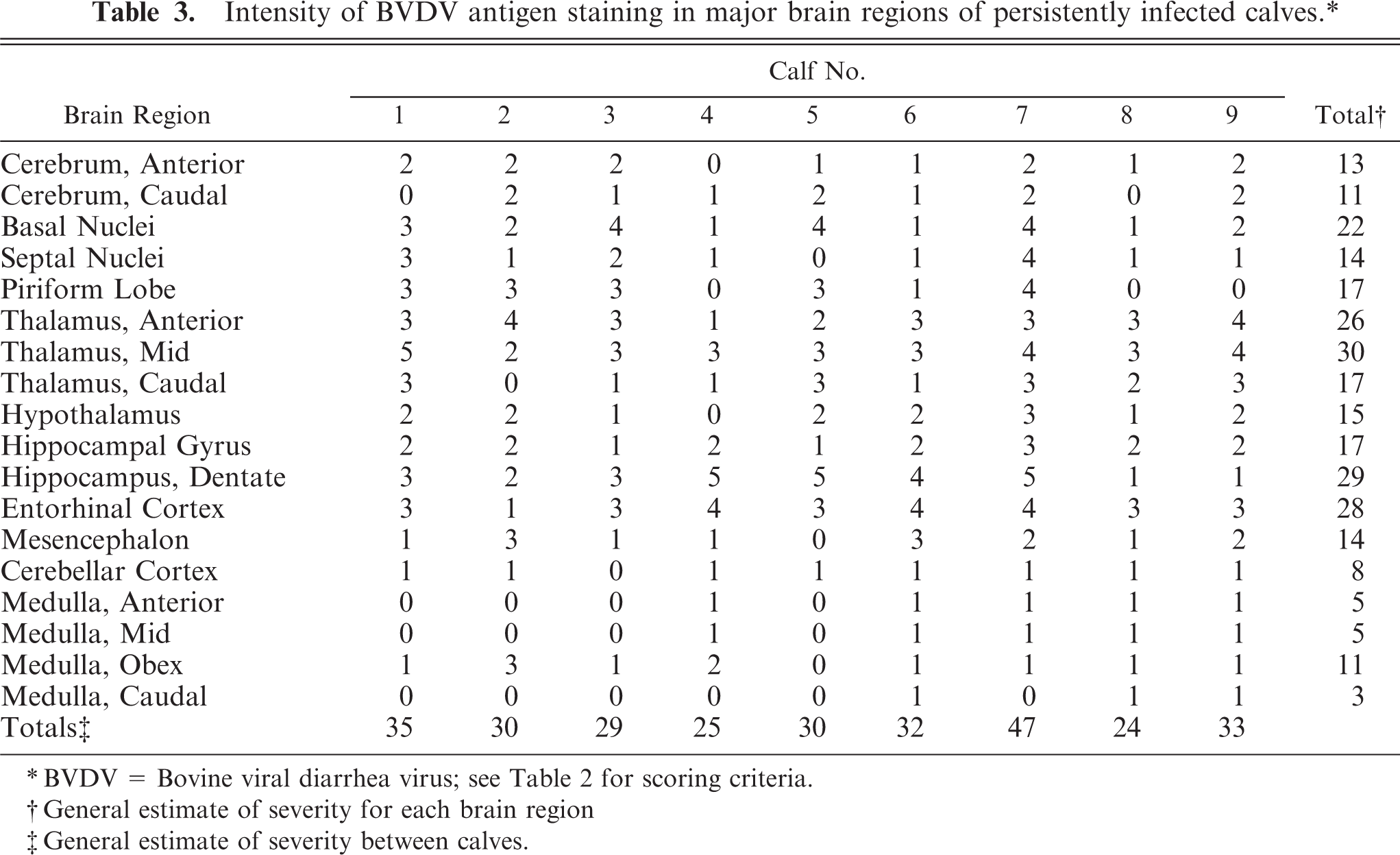
∗ BVDV = Bovine viral diarrhea virus; see Table 2 for scoring criteria.
† General estimate of severity for each brain region
‡ General estimate of severity between calves.
IHC staining
For the detection of BVDV antigen, the primary monoclonal antibody 15C5 (Syracuse Bioanalytical, Ithaca, NY) was employed, and staining was conducted as per published procedure. 20 Briefly, sections were deparaffinize and rehydrated. Following pretreatment with proteinase K and an appropriate blocking step, sections were stained in the primary antibody at a dilution of 1 : 1,000 using an automated immunostainer (DakoCytomation, Carpenteria, CA). Secondary detection used a commercial alkaline phosphatase system (DakoCytomation), fast-red chromogen, and hematoxylin counterstain. The 15C5 antibody was chosen because it is widely used and results are comparable with another monoclonal antibody, 3.12F1 (developed by Dr. J. T. Saliki, Oklahoma State University), on the basis of the author's side-by-side comparisons.
For the primary neuroanatomic reference, the “Navigable Atlas of the Sheep Brain” (http://www.msu.edu/∼brains/brains/sheep/index.html) was used. Although there are some, mainly minor, differences between ovine and bovine brains and terminology is sometimes outdated, this interactive Atlas proved very utilitarian in mapping the distribution of BVDV antigen in the brains of these persistently infected calves. Additionally, this Atlas is readily available to interested readers. For clarity and avoiding confusion, nomenclature used in this Atlas will be followed here.
For the major neuroanatomic areas in each brain section, the distribution and intensity of neuronal BVDV antigen staining were graded (Tables 2,3).
Results
No BVDV antigen was detected in the brains of negative control calves. Positive IHC staining was observed in a variety of cell types in the brains of all persistently infected calves.
Viral antigen in non-neuronal cells
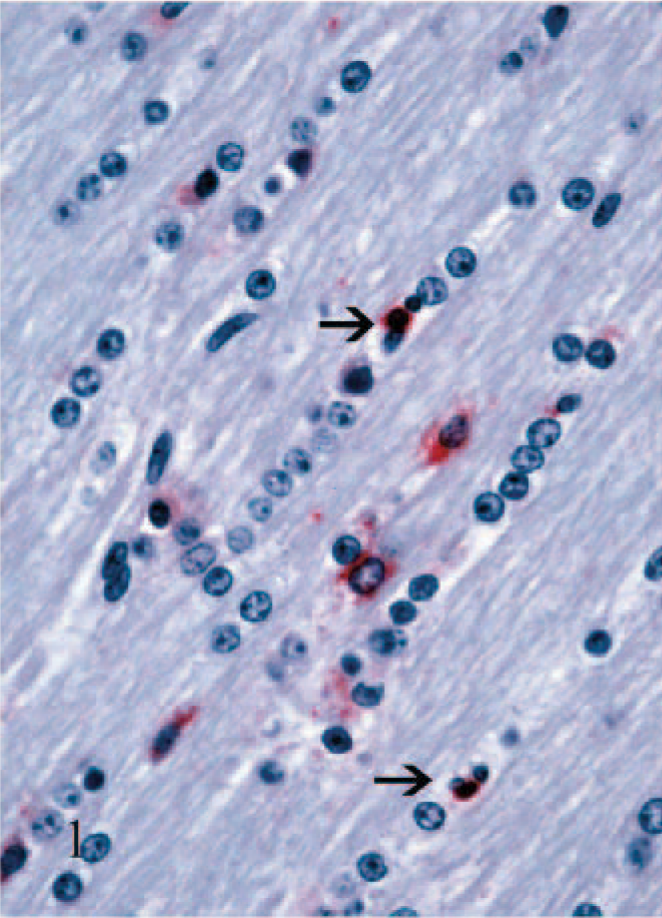
Viral antigen was consistently demonstrated in non-neuronal cells throughout the neuraxis of persistently infected calves. In non-neuronal cells, diffuse-to-focal cytoplasmic staining was granular and faint to intense. In both gray and white matter areas, staining of cells having astrocytic as well as oligodendroglial nuclear morphology (macroglia) was observed (Fig. 1). Staining in macroglial cells was often faint but, as a rule, was more intense and widespread in more anterior brain sections and in the neuropile of severely affected gray matter and contiguous white matter. In more caudal areas of the brain stem where neuronal staining was often minimal, staining of macroglia persisted. Staining in cells having microglial morphology was not detected.
Corpus callosum; calf No. 5. Staining for BVDV antigen in macroglial cells, interfascicular oligodendrocytes (arrows), and astrocytes (larger cells near the center). Alkaline phosphatase, fast-red, hematoxylin counterstain.
Staining of blood vessel–associated cells was conspicuous and generally followed the pattern observed with the macroglia. The most obvious staining was in round cells having macrophage morphology surrounding medium-to-large-caliber blood vessels (arteries, veins, arterioles, venules). Staining in these cells was typically intense (Fig. 2). Other blood vessel–associated staining was notable in small-caliber blood vessels (i.e., capillaries and possibly venules). Here the cells tended to have more spindloid cellular morphology and were in the walls of the small blood vessels (Fig. 3). The identity of these cells could not be determined with certainty due to limitations in the methods used in this study, but on the basis of the absence of endothelial cell staining in larger blood vessels, the cells were interpreted as most likely pericytes. Positively stained cells in the leptomeninges followed a pattern similar to the blood vessel–associated staining, vide supra. Additionally, in some of the larger arteries, staining of medial smooth muscle cells was also noted. No staining was detected in ependymal cells, and only rarely was viral antigen detected in choroid plexus epithelial cells. In a few calves, staining of infiltrating macrophages and/or vascular-associated cells was also noted in the stroma of the choroid plexus, but this was invariably limited.
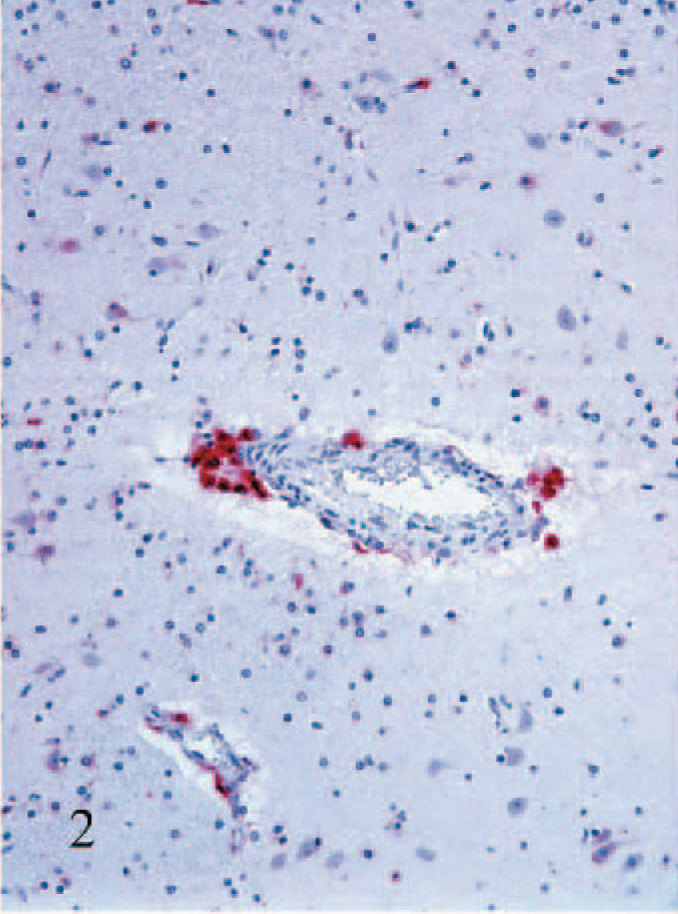
Putamen; calf No. 7. Strong staining for BVDV antigen in perivascular macrophages. Alkaline phosphatase, fast-red, hematoxylin counterstain.
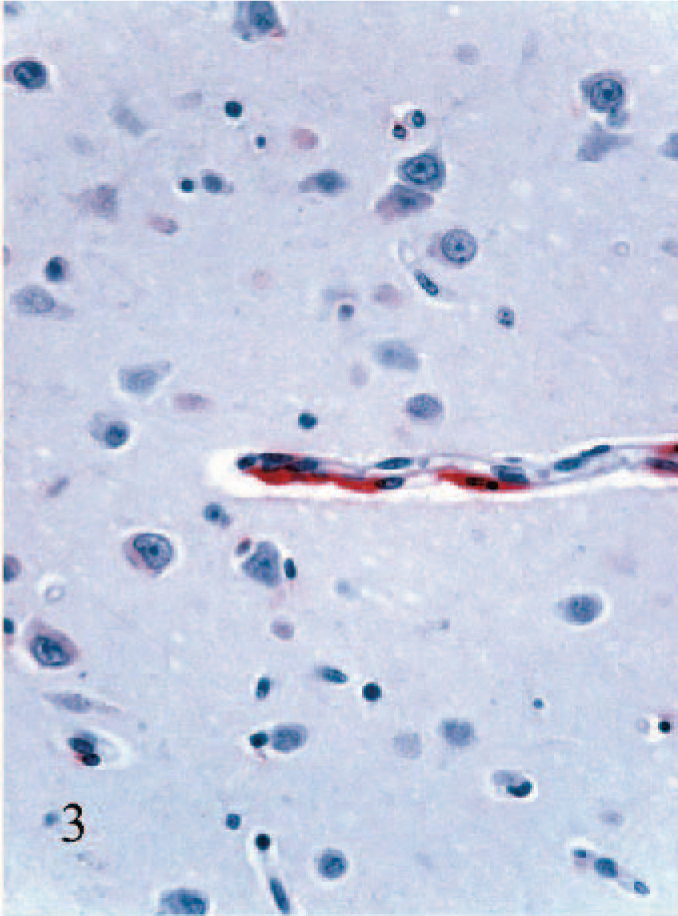
Cerebral cortex; calf No. 2. Staining for BVDV antigen in spindloid cells interpreted as pericytes in the wall of a small blood vessel. Alkaline phosphatase, fast-red, hematoxylin counterstain.
Viral antigen in neurons
Widespread staining of neurons and occasionally of their processes was noted. Neuronal staining was cytoplasmic and varied from dense, granular, and perinuclear to a more diffuse red cytoplasmic blush. The topography and staining intensity in the brains of the 9 calves and results of grading are summarized (Table 3). Where available in the brain sections, bilateral involvement of specific nuclei occurred, but this was not always the case. Where evident in individual calves, the intensity of staining was often unequal in severity. Although variable between calves, the most widespread and intense staining occurred in neurons throughout the thalamus, followed by the hippocampus. For clarity and to provide more detail for the interested reader, the topography of staining in some of the major brain areas will be described.
Cerebral cortex
A characteristic of the staining pattern in cortical neurons was variability. Staining was often patchy in distribution, varying between mild and moderate, and affecting neuronal cell bodies in all lamina, with no appreciable topographic predilection.
Basal nuclei
Intensity of staining in the basal nuclei was essentially similar in individual animals when comparing the caudate nucleus, putamen, and globus pallidus. Staining was consistently present but varied from mild to moderate or moderate with focal areas of more intense staining.
Thalamus
Staining was variable in intensity and distribution but was ultimately evident in virtually all identified thalamic nuclei in the various calves (Fig. 4). In more anterior portions of the thalamus, affected dorsal and midline nuclei (i.e., paraventricular and rhomboid) were sometimes intensely stained while the more lateral nuclei (i.e., anteroventral, anteromedial, and reticular) were not stained. In sections taken from the mid to more caudal thalamus, the pattern of staining extended laterally and to more ventral areas. In these lateral and more ventral thalamic areas, variations in intensity of staining between calves occurred in the named nuclei with little appreciable consistency. Thalamic nuclei that had moderate-to-intense, widespread neuronal staining are listed (Table 4).
Thalamic nuclei with intense, widespread(grades 3–5) neuronal expression of BVDV antigen as demonstrated by IHC staining.

∗ BVDV = Bovine viral diarrhea virus, IHC = immunohistochemical.
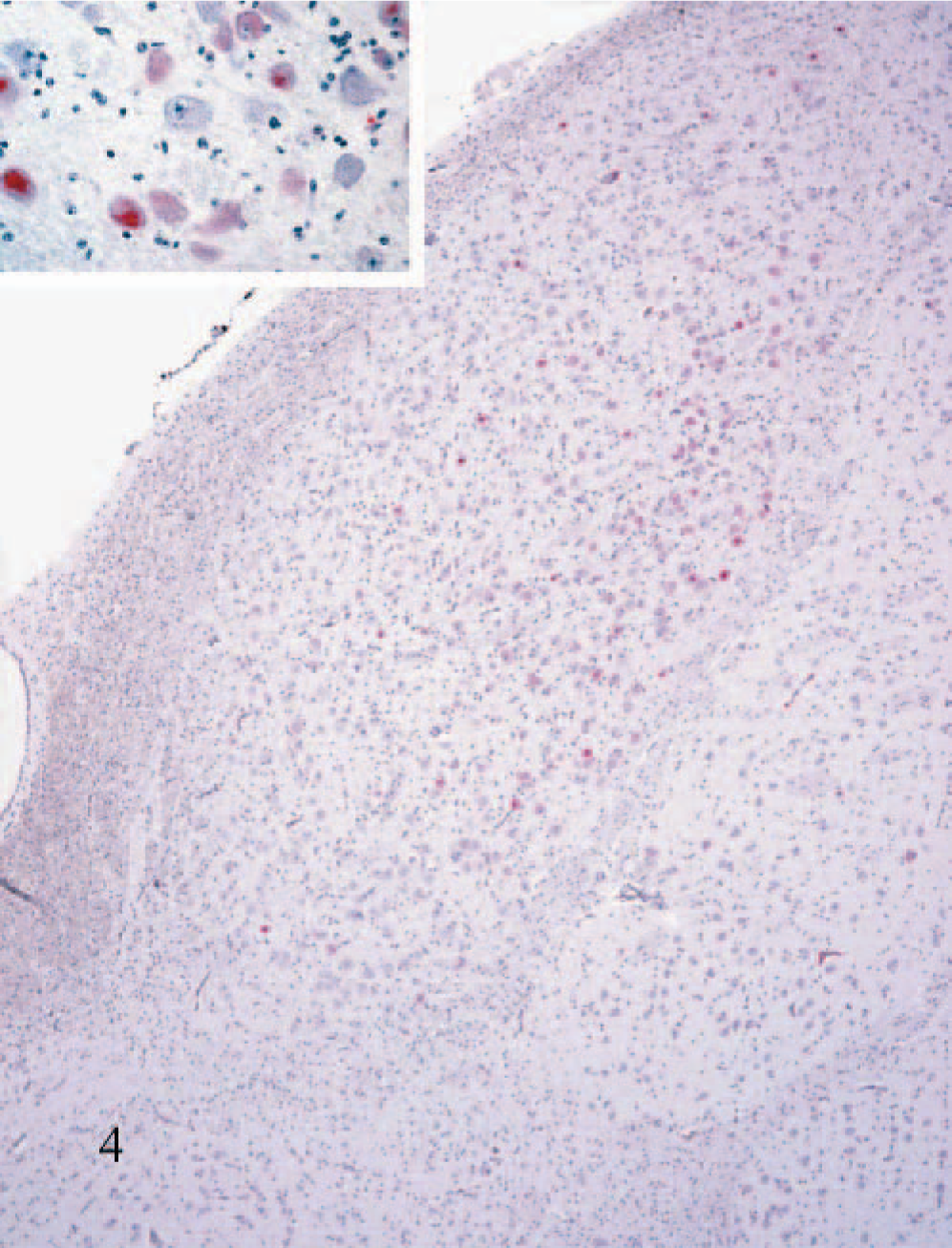
Dorsal and medial thalamus; calf No. 2. Staining for BVDV antigen in many neurons of the dorsal medial thalamic nucleus. Inset: Higher magnification of the neuronal cytoplasmic staining. Alkaline phosphatase, fast-red, hematoxylin counterstain.
Hypothalamus
The major nuclei in the hypothalamus with positive staining for viral antigen were the paraventricular (8 of 9 calves), supraoptic (8 of 9 calves), and periventricular (5 of 9 calves) nuclei. In the paraventricular hypothalamic nucleus, staining was limited to magnocellular neurons. In 2 calves, mild (grade 1) staining was present in the preoptic hypothalamic area. In 1 calf each, mild staining was also found in suprachiasmatic, ventromedial, and arcuate nuclei.
Hippocampal formation
Staining in the hippocampal formation was again variable but prominent in most calves. As a general trend, staining was more intense and widespread in the dentate gyrus (Fig. 5), compared with the hippocampal gyrus. Some, but generally limited, staining was also observed in polymorphous layers. Invariably, ventral areas of the hippocampal formation showed more intense and diffuse staining than the more dorsal aspects.
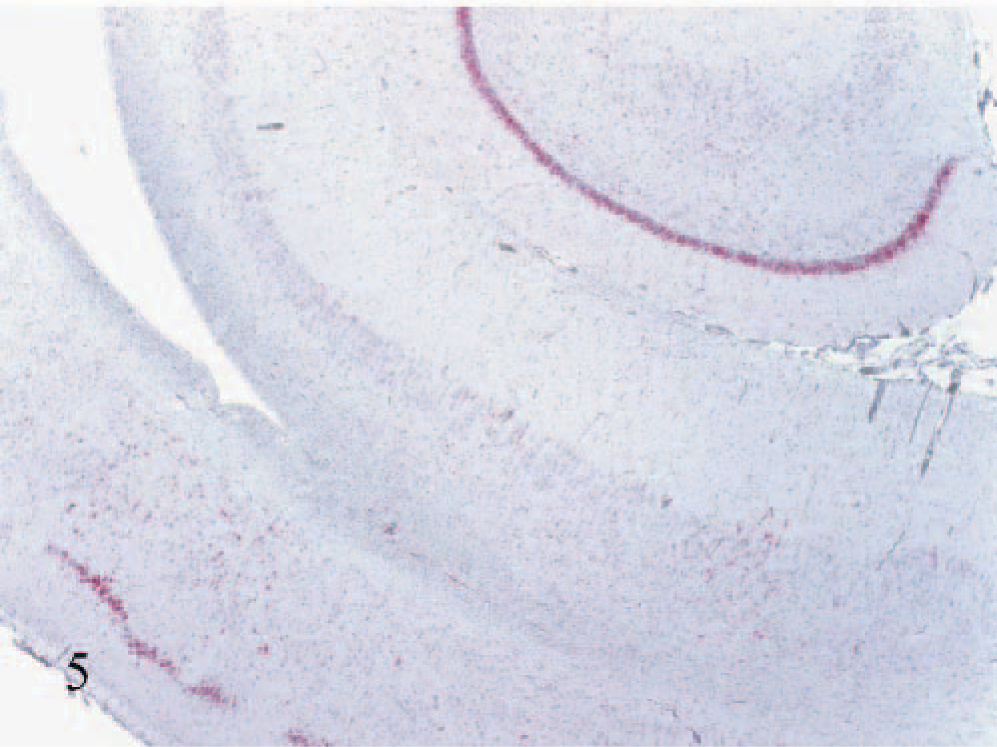
Hippocampus and entorhinal cortex (lower left); calf No. 7. Strong staining for BVDV antigen in neuronal cell bodies of the hippocampal dentate gyrus and entorhinal cortex. Patchy staining is also evident in the hippocampal gyrus and polymorphous layer of the hippocampal formation. Alkaline phosphatase, fast-red, hematoxylin counterstain.
Mesencephalon
When present in the mesencephalon, staining was generally mild. Positive staining was found in the following nuclei (with number of calves indicated in parentheses): pars lateralis of substantia nigra (1), medial geniculate (6), rostral (superior) colliculus (4), lateral nucleus (1), caudal colliculus (4), periaqueductal gray matter (3), linear raphe nucleus (2), and neurons in reticular areas (1).
Cerebellar cortex
Neuronal staining in the cerebellar cortex was minimal to mild in all calves, and no staining was evident in 1 calf. In calves with persistence of an external granule cell layer, mild staining occurred in few scattered cells. In the molecular layer, staining of scattered small cells with little discernible cytoplasm was evident, but it was not possible to conclusively determine the cell types. In the granule cell layer, staining was evident in scattered Lugaro and Golgi cells. Staining was never observed in Purkinje cells or in the granule cells.
Medulla oblongata: With few exceptions, neuronal staining in the medulla was lacking or mild. Nuclei most consistently affected were the hypoglossal, dorsal motor nucleus of the vagus nerve, and ventral raphe nuclei, which were stained in 5 calves.
Discussion
Results of the current and previous studies indicate that the fetal brain is a primary target of persistent infection and that the infection persists well after birth. 26, 38, 86 It is unknown whether the BVDV infection in the brain represents a static process (i.e., cells are infected in utero and remain infected but with little to no spread to new brain areas). Alternatively, it is conceivable that the initial in utero infection represents a dynamic process with changing patterns of cellular antigen expression or infection of cells in additional brain areas. Whether the infection is static or dynamic could have an impact on the clinical manifestations of disease.
In this study, BVDV antigen was detected in macroglial, blood vessel–associated, and neuronal cells. The heterogeneity of affected cell types differs from previous studies employing IHC staining and a variety of primary antibodies on frozen as well as formalin-fixed, paraffin-embedded sections. One of the earliest studies indicated staining only in small neurons or possibly microglia. 57 Later studies indicated that staining was exclusively confined to neurons. 26, 38, 86 A satisfactory explanation for these disparate results could not be determined. Antigenic specificity of the different primary antibodies could have played a role. 26, 38, 80 One study suggested that the 15C5 monoclonal antibody gave results similar to those of other antibodies for the detection of BVDV antigen. 80 Antigenic variability in the infecting strains of ncpBVDV could also be proposed. No major differences were found between calves infected with type 1 or type 2 ncpBVDV in this study, however. The varied cells expressing BVDV antigen observed in this study were more like those reported in sheep following in utero infection with ncpBVDV. 39 It seems exceptional that BVDV would infect only neurons while a variety of cell types are found to be infected in non-nervous system tissues.
The topography of BVDV antigen distribution in the brains of the current persistently infected calves also differs from that in previous studies. In 3 major studies, antigen was detected primarily in cerebrocortical and hippocampal neurons. 26, 38, 86 In other areas, staining was observed in only small groups of neurons in affected nuclei. Cerebrocortical and hippocampal neurons were certainly infected in the current study, the latter sometimes severely, but when the volume of affected brain tissue was considered, the most consistent and intense staining for BVDV antigen occurred in the thalamus. Again, an adequate explanation for the differences between the current and past studies is undetermined.
In past in vivo as well as in vitro studies of organs and target cells involved in growth, reproduction, and immunologic defenses in BVDV-infected tissues and cells, a cause-effect relationship between infection and functional deficits has been alluded to. Conversely, there are few reports of definitive neurologic symptoms in persistently infected calves. 26, 86 Consequently, the functional significance of persistent BVDV infection in the brain has not been determined. Possibly because of the lack of consistent lesions by light microscopy and the absence of defined clinical deficits, the brain has received little attention with regard to pathogenesis of BVDV-induced clinical disease. It would seem naïve to conclude that viral infection in non-neural target cells would lead to functional deficits (i.e., growth retardation and reproductive failure as well as immunosuppression), while despite evidence of widespread infection in the brain, no functional abnormalities related to the nervous system would occur. One dramatic and characteristic finding in the current study is the variability in BVDV antigen expression in different brain areas, in different cell types, and between calves. A question that must be asked is if there is any correlation between this variability in evidence of brain infection and the protean nature of BVDV-induced disease in postnatal calves. Although an exact birth date was not available and the calf was otherwise grossly unremarkable, it is of interest that in calf No. 7, BVDV antigen was the most widespread and intense of any of the calves and this calf had a considerably lower body weight compared with persistently infected herd mates (Tables 1,3).
Complex interactions occur between the nervous, endocrine, and immune systems that fine-tune and orchestrate the functional maturation of these organ systems. 24, 63, 66 Disturbances in utero or early in life can have profound effects on these relationships. 54, 55, 72 In addition to viral infection of target endocrine organs controlling growth, metabolism, and reproduction (i.e., pituitary gland, 86 pancreatic islets, thyroid gland, 80 and reproductive organs 11, 27, 29, 35 ) in BVDV infection, the potential for brain-endocrine-immune disruption via the hypothalamic-pituitary-endocrine axis is well known and needs little further elaboration. 24, 25, 55 Less recognized are other multidirectional pathways between the brain, endocrine, and immune organs. During fetal life, organs of the immune system become richly innervated by autonomic and peripheral nerves that are requisite for normal development and later function. 24, 25, 53 In concert, leukocytes express receptors for neuropeptides and neurotransmitters that modify their functions. 45, 53 Leukocytes also express a variety of receptors for hormones traditionally viewed as having effects on growth, reproduction, and other nonimmune actions that affect the function of these immune-effector cells. 18, 21
Functional interactions between the nervous and immune systems are bidirectional. Cytokines produced in the brain and by peripheral immune-effector cells impact brain and hypothalamic-pituitary-endocrine responsiveness. 23, 40, 49, 52, 70, 74, 81, 85 Systemic as well as localized peripheral immune stimuli modify the response of the brain via afferent nerves or more directly via the blood and vasculature. Endothelial cells, pericytes, and perivascular macrophages have receptors for certain cytokines. These as well as astrocytes and microglia can produce cytokines including interleukin (IL)-1, IL-6, and tumor necrosis factor-alpha. Cytokines can suppress or stimulate inflammatory and immune as well as endocrine responses. 8, 70 Apart from the well-known corticotropin-releasing factor and adrenocorticotrophic hormone produced by the hypothalamus and pituitary gland, respectively, other mediators of neuroendocrine and immune functions are listed along with the references (Table 5). These mediators often interact with other stimuli to culminate in the effect. The ultimate endocrine and/or immune responses reflect the summation and balance of inhibitory and stimulatory influences.
Summary of mediators between nervous, endocrine, and immune systems.
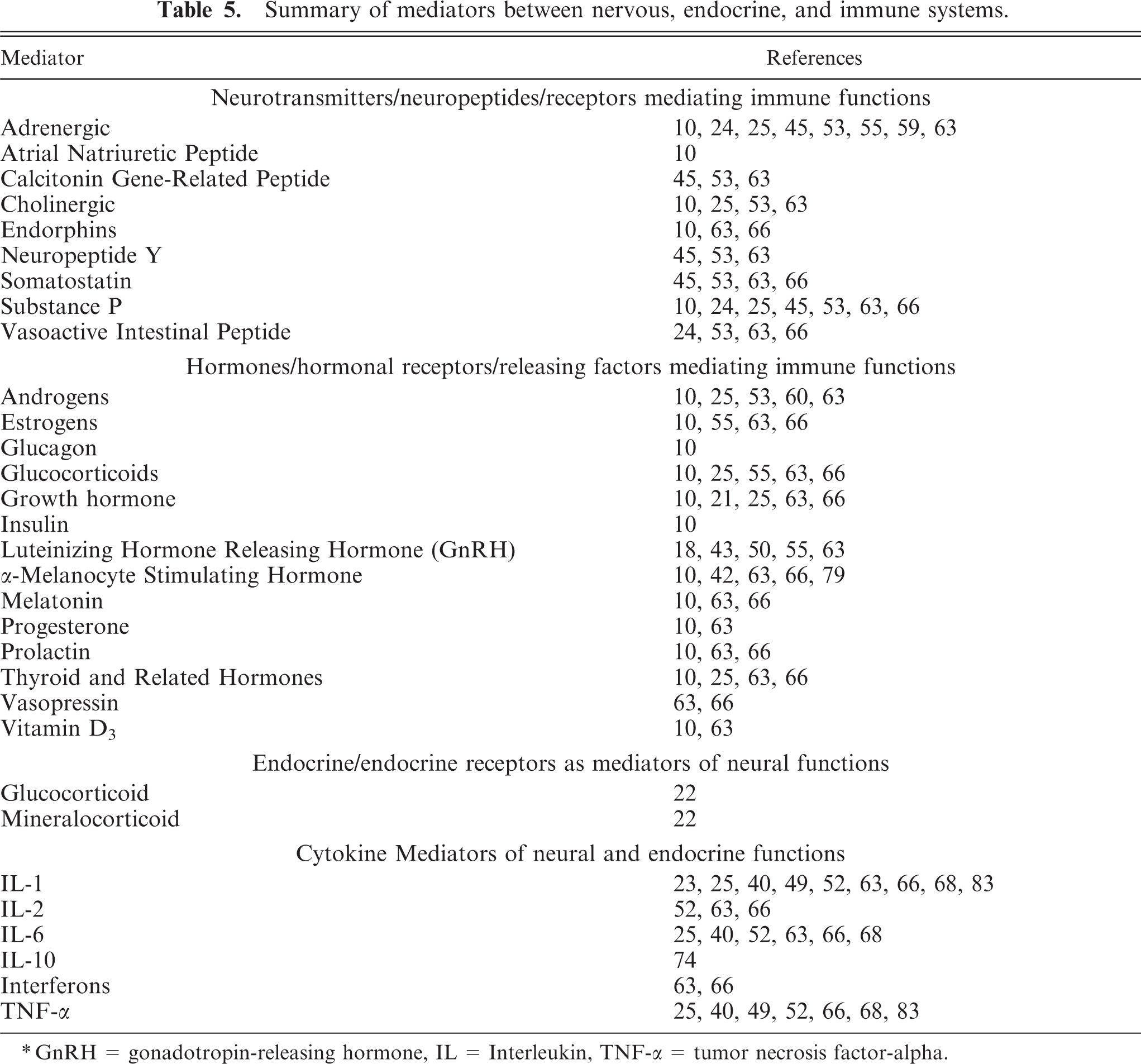
∗ GnRH = gonadotropin-releasing hormone, IL = Interleukin, TNF-α = tumor necrosis factor-alpha.
The current study identifies evidence of BVDV infection in multiple brain cell types and brain areas that have the potential to modify parameters of growth, reproduction, and immune responsiveness. Viral antigen was demonstrated consistently in perivascular macrophages in these 9 persistently infected calves. Perivascular macrophages are migratory cells of the macrophage-monocyte lineage rather than resident cells. 9 As migratory cells, infection could constantly expose the brain microenvironment to viral antigen with production of endogenous cytokines or, conversely, contribute to an anergic state. Antigen expression in cells within the walls of small cerebral blood vessels was demonstrated. Previous studies have suggested that staining of endothelial cells with the 15C5 antibody is spurious, 80 while others have indicated staining of endothelial cells to be valid. 39 In the current study, staining of cells in the walls of smaller blood vessels was interpreted as most likely in pericytes. Arguments in favor of this conclusion include the known limited localization of pericytes to small blood vessels and failure to demonstrate antigen in endothelial cells of larger blood vessels. 3, 6, 73 Pericytes participate in the formation of the blood-brain barrier, transport across the blood-brain barrier, phagocytosis, and immune recognition-response functions. 3, 6, 73 Viral antigen is also demonstrable in astrocytes. These cells participate in and modify immune reactions in the brain by production of a variety of trophic factors and inflammatory mediators that could alter brain function and ultimately endocrine and immune responsiveness. 8, 55, 70, 83 Oddly, the current and several past studies have not reported evidence of BVDV infection in microglia. This seems exceptional considering that peripheral blood monocytes and other brain cells are targets of persistent infection. Microglia could, however, respond to BVDV infection by production of endogenous cytokines and other mediators.
The topography of BVDV antigen expression in the brains of persistently infected calves could also provide clues relevant to the impact of brain infection on growth, reproduction, and immune responsiveness. Paraventricular and supraoptic hypothalamic nuclei, known for their production of releasing factors for hormones produced in the pituitary gland, were affected in most of the calves. Secondly, the dorsal motor nucleus of the vagus nerve, a principal efferent nucleus of the autonomic nervous system innervating peripheral immune organs, was affected in several calves. Infection in the thalamus could modulate the nervous input to other brain sites. Thalamic nuclei are sites for relay, modification, and integration of sensory and motor nerve impulses destined for other brain areas. 75 Recent research has suggested that dorsal and midline thalamic nuclei, sites consistently affected in BVDV persistently infected calves, can modulate immune responsiveness via the hypothalamic-pituitary axis. 58 Other brain areas expressing viral antigen, the hippocampus and entorhinal cortex, can modify hypothalamic-pituitary functions. 13, 22, 82 Although the relevance is undetermined at this time, the thalamic midline and entorhinal cortex are also brain areas, among others, having a high density of interferon-gamma receptors. 69
Research dealing with infection of target cells of the endocrine and immune systems on the clinical manifestations of BVDV-induced disease is important and valuable in elucidating the impact of BVDV persistent infection in calves. Just as these studies are enlightening, the potential for infection in the brain to impact function of the endocrine and immune systems leading to BVDV-induced clinical disease cannot be discounted.
Footnotes
Acknowledgements
This study was funded by a Faculty Grant-in-Aid from the University of Wyoming (UWGIA49315). I thank Paula T. Jaeger and Mercedes Thelen, histotechnologists, for their technical expertise and Drs. R. W. Storts and L. W. Woods for their thoughtful reviews of the manuscript.