
Abstract
The primary complication of seasonal influenza in humans is viral pneumonia. A conventional animal model—intranasal inoculation of ferrets with 106 median tissue culture infectious dose of virus—results in disease that is neither consistent nor comparable with severe viral pneumonia in humans. Therefore, the authors modified the experimental procedures by increasing the median tissue culture infectious dose to 109 and by inoculating via the intratracheal route, testing these procedures with H1N1 strains (A/Bilthoven/3075/1978 and A/Netherlands/26/2007) and H3N2 strains (A/Bilthoven/16190/1968 and A/Netherlands/177/2008) of seasonal influenza virus. The ferrets of all groups (n = 3 per virus strain) had clinical signs, increased body temperature, virus excretion from day 1, loss of body weight, and increased relative lung weight at 4 days postinoculation. All ferrets had severe pulmonary consolidation, and histologic examination revealed moderate to severe necrotizing bronchointerstitial pneumonia with severe edema, necrosis of alveolar epithelium, inflammatory infiltrates in alveolar septa and lumina, epithelial regeneration, and perivascular and peribronchiolar inflammatory infiltrates. The lesions were associated with the presence of influenza virus antigen in respiratory epithelium by immunohistochemistry. Although all 4 virus strains caused pulmonary lesions of comparable severity, virus isolation in the lungs, trachea, nasal concha, and tonsils showed higher mean virus titers in the H1/07 and H3/68 groups than in the H1/78 and H3/08 groups. In conclusion, the above H1N1 and H3N2 strains cause severe pneumonia in ferrets by use of the modified experimental procedures and provide a good model for pneumonia caused by seasonal influenza A virus infection in humans.
Seasonal human influenza viruses cause viral pneumonia as a primary complication in the elderly, immunosuppressed, and very young. 17 In currently used animal models, seasonal influenza virus infection does not consistently cause viral pneumonia.10,13,15 Therefore, it is difficult to study the pathogenesis of this complication or evaluate preventive or therapeutic measures against it.
Ferrets are commonly used as experimental animals to study the pathogenesis of human influenza and test vaccines and antivirals against influenza virus infection. Ferrets and humans have similarities in their respiratory tract and in their interaction with human influenza viruses. Ferrets have similar lung physiology and airway morphology as humans. 9 Human influenza viruses show a similar pattern of attachment to ferret respiratory tract epithelium. 16 Both species are susceptible to infection with human influenza viruses.7,9 Both develop similar disease: usually upper respiratory tract disease, which is mild, and occasionally pneumonia, which is more severe or even fatal.7,9 That human influenza virus infection usually causes only mild disease in ferrets is a drawback if one wishes to have an animal model for severe influenza viral pneumonia.
Outcome of experimental influenza virus infection can be modified in different ways, including route of administration, viral dose, and virus strain. Intranasal inoculation of ferrets with seasonal human H1N1 influenza virus causes rhinitis without involvement of the lower respiratory tract,10 while intratracheal inoculation of the same virus strain causes bronchointerstitial pneumonia. 15 Increasing the dose of viral inoculum from 10 4 to 10 8 mean tissue culture infectious dose (TCID50) results in a higher mortality rate.15 Influenza virus strains differ in pathogenicity. For example, the strain of H1N1 virus that emerged in 2009 and caused a human pandemic causes more severe disease in different species of experimental mammals than the strain of H1N1 virus that has been circulating for decades in the human population.5,6,15
Making use of the above knowledge, we adapted the experimental procedures with the objective of establishing a model of human seasonal influenza virus infection that consistently induces pneumonia in ferrets. In this model, we used an intratracheal route of inoculation and a high viral dose of 10 9 TCID50. We tested these modified experimental procedures for subtypes H1N1 and H3N2 and for recent and older strains of these subtypes.
Materials and Methods
Study Design
We chose for intratracheal inoculation because this method delivers the viral inoculum closer to the site of pneumonia than does intranasal inoculation. Also, we have shown that intratracheal inoculation more consistently induces pneumonia in ferrets than intranasal inoculation does, both for seasonal H1N1 influenza virus15 and for highly pathogenic avian H5N1 influenza virus. 2
We chose for an inoculum dose of 10 9 TCID50 on the basis of a previous study, where we had used doses ranging from 10 4 to 10 8 TCID50 to inoculate ferrets intratracheally with seasonal H1N1 influenza virus.15 Because even ferrets inoculated with a dose of 10 8 TCID50 did not show mortality by 7 days postinoculation (dpi), we chose for a dose 10× higher, 10 9 TCID50, which is the highest dose practical for most virus strains by use of current laboratory techniques.
We chose for an H1N1 virus strain (H1/07) and an H3N2 virus strain (H3/08) as representative for humans seasonal influenza viruses circulating most recently in the human population. For contrast, we chose an H3N2 virus strain (H3/68) that had been isolated soon after this subtype had been introduced into the human population in 1967 and an H1N1 virus strain (H1/78) that had been isolated soon after this subtype had been reintroduced into the human population in 1977. We considered that the older viruses might be more pathogenic in ferrets because they had not had as much time to adapt to replication in the mammalian host as more recent viruses.
For each virus (4 groups), a group of 3 ferrets was inoculated intratracheally with 10 9 TCID50 in a 3-ml volume under anesthesia with ketamine (Nimatek, Eurovet Animal Health BV, Bladel, Netherlands) and medetomidine hydrochloride (Domitor, Orion Pharma, Espoo, Finland). From 0 to 4 dpi, ferrets were monitored daily for clinical signs, and under ketamine–medetomidine hydrochloride anesthesia, body weight was measured and pharyngeal swabs collected. 1 At 4 dpi, necropsies were performed, and samples were taken from respiratory and extrarespiratory tissues for virologic, pathologic, and immunohistochemical analyses. The experiments were performed under an animal study protocol approved by the Institutional Animal Welfare Committee and performed compliant with national and European legislation. All experiments were performed under biosafety level 3 conditions at the Netherlands Vaccine Institute.
Virus Preparation
Four seasonal influenza viruses were isolated from patients during different influenza seasons: H1N1 strains A/Bilthoven/3075/1978 (H1/78) and A/Netherlands/26/2007 (H1/07) and H3N2 strains A/Bilthoven/16190/1968 (H3/68) and A/Netherlands/177/2008 (H3/08). The isolates were passaged 3 times in Madin-Darby canine kidney cells and titrated according to standard methods. Subsequently, the viruses were purified and concentrated about 20-fold using sucrose gradient centrifugation, reaching an infectious virus titer of 1 × 10 9 TCID50 per milliliter.1,11
Ferrets
Twelve 8-month-old purpose-bred female ferrets—seronegative for antibodies against circulating influenza viruses H1N1, H3N2, pandemic (H1N1) 2009, and Aleutian disease virus—were maintained in standard housing and provided with commercial food pellets and water ad libitum until the start of the experiment. All ferrets were female (body weight, 690–1010 g). Two weeks preinfection, the animals were anesthetized with ketamine and medetomidine hydrochloride, and a temperature logger (DST micro-T Ultra Small Temperature Logger, Star-Oddi, Reykjavik, Iceland) was placed in the peritoneal cavity. This device recorded the body temperature of the animals every 15 minutes. Changes in body temperature were calculated by subtracting the mean day and night temperature measured on 2 successive days in the period before the challenge from the mean day and night temperatures postinfection. For the relative lung weight of control ferrets, the lungs of 22 noninfected female ferrets were weighed, expressed as a percentage of the total body weight.
Pathology
The animals were euthanized by exsanguination under ketamine anesthesia at 4 dpi and necropsied according to a standard protocol. The trachea was clamped off so that the lungs would not deflate upon opening the pleural cavity, thereby allowing visual estimation of the area of affected lung parenchyma. Samples for histologic examination were stored in 10% neutral buffered formalin (lungs after inflation with formalin), embedded in paraffin, sectioned at 4 μm, and stained with hematoxylin and eosin for examination by light microscopy. The following tissues were examined by light microscopy: left lung (4 slides with either cranial or caudal lobe with longitudinal or cross section), nasal turbinate, nasal septum, larynx, trachea, bronchus, tracheobronchial lymph node, eyelid, tonsil, heart, liver, spleen, kidney, and brain.
Semiquantitative assessment of influenza virus–associated inflammation in the lung was performed as reported earlier: 14
For the extent of alveolitis and alveolar damage: 0, 0%; 1, 1%–25%; 2, 25%–50%; 3, > 50%.
For the severity of alveolitis, bronchiolitis, bronchitis, and tracheitis: 0, no inflammatory cells; 1, few inflammatory cells; 2, moderate numbers of inflammatory cells; 3, many inflammatory cells.
For the presence of alveolar edema, alveolar hemorrhage, and type II pneumocyte hyperplasia: 0, no; 1, yes.
For the extent of peribronchial, peribronchiolar and perivascular infiltrates: 0, none; 1, 1–2 cells thick; 2, 3–10 cells thick; 3, more than 10 cells thick.
Slides were examined without knowledge of the identity of the animals. The cumulative scores for size and severity of inflammation of all slides provided the total score per animal. As a control animal for histopathology and immunohistochemistry, a ferret was sham inoculated with phosphate buffered saline and euthanized at 4 dpi.
Immunohistochemistry
For detection of influenza A virus antigen, tissues were stained with a primary antibody against the influenza A nucleoprotein as described previously. 12 Alternatively, binding of the primary antibody was detected using a peroxidase labeled goat-anti-mouse IgG2a (Southern Biotech, Birmingham, AL). Peroxidase activity was revealed using 3-amino-9-ethylcarbazole (Sigma, St Louis, MO), resulting in a bright red precipitate. In each staining procedure, an isotype control was included as a negative control, and a lung section from an experimentally influenza-inoculated cat (H5N1) was used as positive control.
Semiquantitative assessment of influenza virus antigen expression in the lungs was performed as reported earlier: 14 for the alveoli, 25 arbitrarily chosen fields (20× objective) of lung parenchyma from 4 lung sections were examined by light microscopy for the presence of influenza virus nucleoprotein, without the knowledge of the identity of the animals. The cumulative scores for each animal were presented as number of positive fields per 100 fields. For the bronchi and bronchioles, the percentage of positively staining bronchial and bronchiolar epithelium was estimated on every slide, and the average of the 4 slides provided the score per animal: 0, 0%; 1, 1%–25%; 2, 25%–50%; 3, > 50%.
Virology
After collection on 0, 1, 2, 3, and 4 dpi, pharyngeal swabs were stored at –70°C in the same medium as that used for the processing of the lung samples. Quadruplicate 10-fold serial dilutions of lung and swab supernatants were used to determine the virus titers in confluent layers of Madin-Darby canine kidney cells as described previously. 11
Samples of all lobes of the right lung and the accessory lobe, nasal concha, trachea, tonsil, tracheobronchial lymph node, spleen, brain, third eyelid, liver, kidney, and heart from all animals were collected at 4 dpi and stored at –70°C until further processing. The samples were weighed and subsequently homogenized with a FastPrep-24 (MP Biomedicals, Eindhoven, Netherlands) in Hank’s balanced salt solution containing 0.5% lactalbumin, 10% glycerol, 200 U/ml of penicillin, 200 μg/ml of streptomycin, 100 U/ml of polymyxin B sulfate, 250 μg/ml of gentamycin, and 50 U/ml of nystatin (ICN Pharmaceuticals, Zoetermeer, Netherlands), then centrifuged briefly before dilution.
Statistical Analysis
The one-way analysis of variance test for multiple comparisons was used to assess differences between the virus groups for body weight, relative lung weight, percentage affected lung tissue, histologic scores, immunohistochemical scores, and viral load. A posttest was performed only when P < .05 (independent t test). Differences were considered significant when P < .05.
Results
Clinical Findings
The ferrets became lethargic at 2 dpi with loss of appetite, mild dyspnea, and raised body temperature (Fig. 1). Sneezing was not observed in any of the ferrets. The body temperature profile after inoculation of the ferrets was similar in the different groups with an increase in body temperature starting 4 hours after inoculation until 1 dpi with a temperature increase up to 2°C from the baseline. The differences in body temperature and body weight loss among groups (Table 1) were not statistically significant.
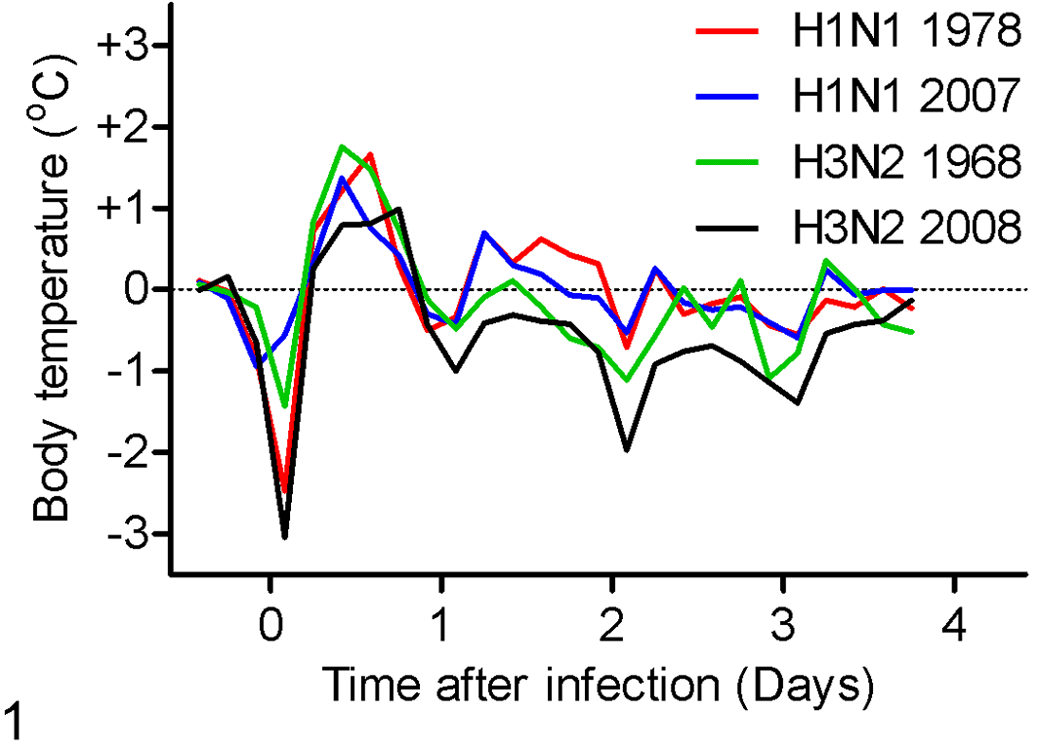
Changes in body temperature from the baseline in ferrets infected with recent and old H1N1 and H3N2 seasonal human influenza viruses at 4 days postinoculation.
Ferrets With Recent and Old Seasonal Human Influenza Viruses a

aPer group, n = 3; 4 days postinoculation.
bThe relative lung weight is expressed as a percentage of the total body weight. The median relative lung weight of control ferrets was 0.6, with a range of 0.5 to 0.8.
Gross Pathology
At necropsy at 4 dpi, all ferrets had multifocal or coalescing pulmonary consolidation, with dark red, raised, and firm areas. The median percentage of lung tissue affected was 40% in all groups (Table 1). The relative lung weights of all ferrets were similar, with only small differences between the groups. The differences in percentage lung tissue affected and relative lung weight among groups were not statistically significant.
Histopathology
By histopathology (Figs. 2 –10), all groups showed a multifocal to diffuse, moderate to severe necrotizing bronchointerstitial pneumonia. This lesion was characterized by flooding of alveolar lumina with edema fluid, fibrin, and cellular debris mixed with alveolar macrophages, neutrophils, and erythrocytes (Fig. 5). The alveolar walls were thickened and had necrosis of the lining epithelium and multifocal type II pneumocyte hyperplasia. The bronchiolar walls had multifocal epithelial necrosis with moderate infiltrates of neutrophils and peribronchiolar multifocal infiltration of moderate numbers of macrophages, lymphocytes, and few neutrophils and plasma cells. In the bronchiolar lumina, there were moderate numbers of macrophages, neutrophils, and erythrocytes, mixed with fibrin, edema fluid, and cellular debris (Fig. 6). Histopathologic changes in the bronchi were similar but less severe (Fig. 7). Multifocally, there was moderate lymphocytic and suppurative tracheobronchoadenitis, tracheitis, and rhinitis. In the other organs, there were no significant changes.
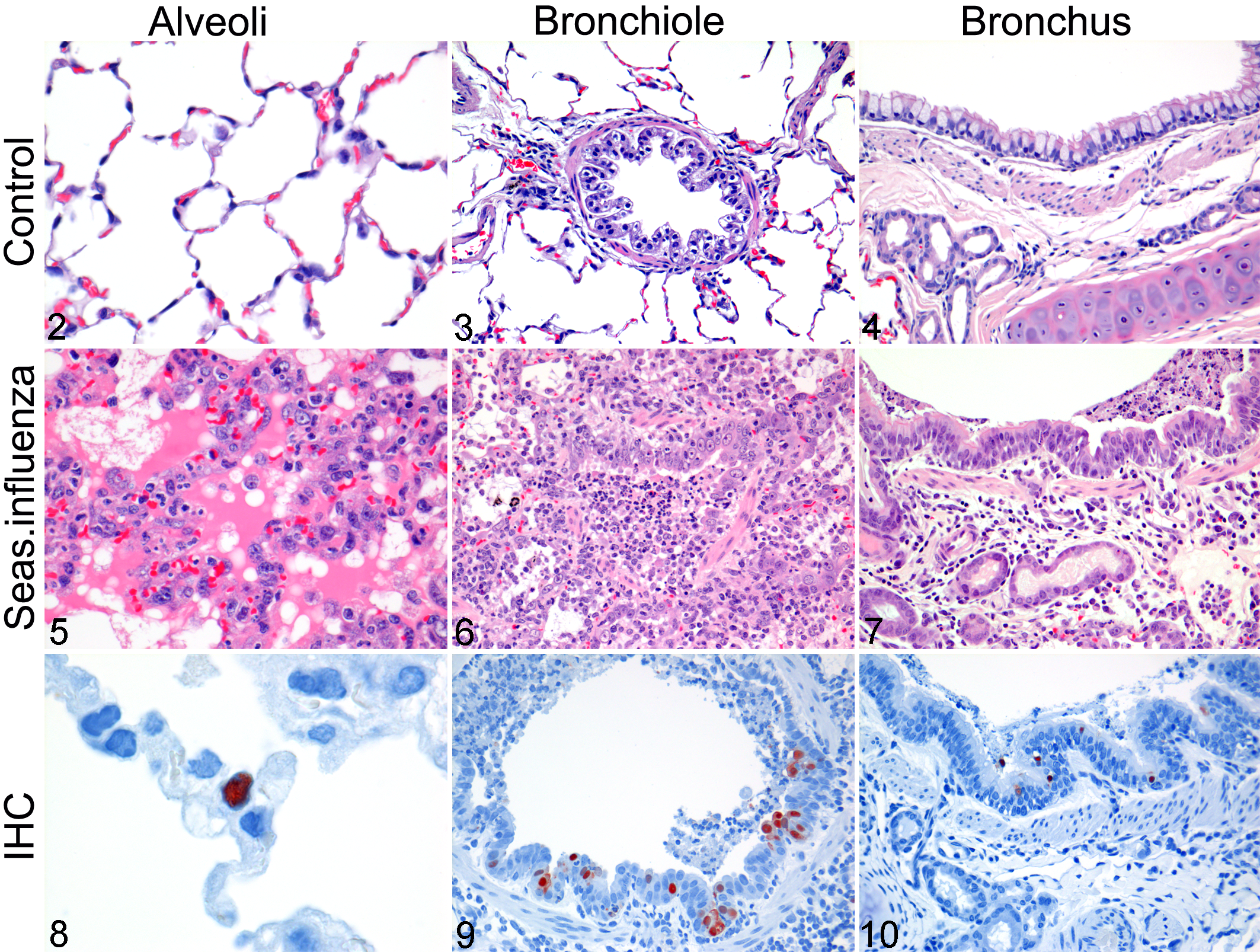
Lung, alveoli; uninfected control ferret. No lesions. HE.
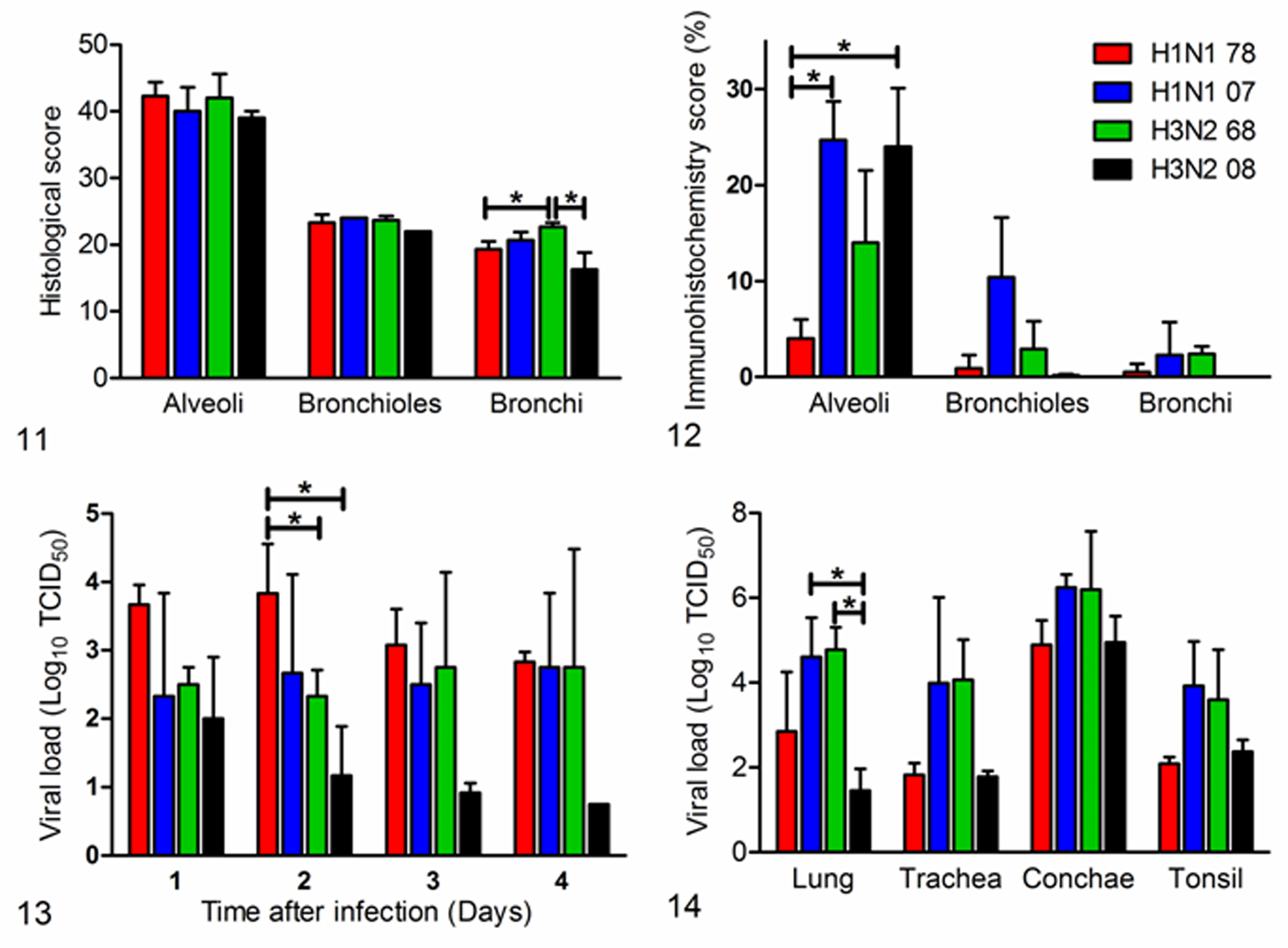
Histologic scoring of the lungs of ferrets infected with recent and old H1N1 and H3N2 seasonal human influenza viruses. Asterisks indicate that values differ significantly between the groups (P = .049 and P = .043, respectively).
Semiquantitative histologic scoring of the respiratory tract showed that the alveolar, bronchiolar, and bronchial lesions were comparable among the viruses (Fig. 11), except for the bronchi, where the scores were significantly higher in the H3/68 group when compared to the H1/78 group (P = .049) and the H3/08 group (P = .043). The differences in histologic scoring of the alveoli and bronchioles were not statistically significant.
Immunohistochemistry
By immunohistochemistry, influenza virus antigen expression was visible as diffuse to granular red staining, which usually was stronger in the nucleus than in the cytoplasm. Influenza virus antigen expression was closely associated with the presence of histologic lesions at different levels of the lower respiratory tract. Influenza virus antigen expression was seen predominantly in type 1 and 2 pneumocytes, alveolar macrophages, bronchiolar epithelial cells, and bronchial epithelial cells (Figs. 8 –10). In the nose, variable numbers of epithelial cells expressed viral antigen that was more present in the H1/07 and H3/08 groups.
Semiquantitative scoring showed that, in the alveoli, the expression of viral antigen was significantly higher in the H1/07 group (P = .005) and the H3/08 group (P = .021) than in the H1/78 group (Fig. 12). In the bronchioles and bronchi, the differences were not statistically different. None of the ferrets had influenza virus antigen expression in extrarespiratory tissues.
Virology
In the pharyngeal swabs of all ferrets, there was excretion of virus up to 4 dpi, except for H3/08 with excretion up to 3 dpi. During the 4 days, there was not a clear peak in virus excretion (Fig. 13). Virus excretion was significantly higher in the H1/78 group when compared to the H3/68 group (P = .049) and H3/08 group (P = .011). At 4 dpi, in the lungs, the virus titers were significantly higher in the H1/07 and H3/68 groups when compared to the groups infected with H3/08 (respectively, P = .013 and .001) (Fig. 14). The differences in virus titers between the other tissues and groups were not statistically significant. In the bronchial lymph nodes, the virus titer was below the detection limit of 0.75 TCID50 per gram of tissue. In nonrespiratory organs, 1 ferret inoculated with H1/07 virus had virus in the heart (101.6 TCID50 per gram of tissue), and 7 animals had virus in the brain: 2 animals each in the H1/07 and H1/78 groups and all 3 in the H3/68 group (range, 101.0–102.1 TCID50 per gram of tissue). No virus was detected in other tissues.
Discussion
In this study, we show that our experimental procedure of using a seasonal influenza virus inoculum of 10 9 TCID50 via the intratracheal route consistently causes a moderate to severe pneumonia in ferrets. Colocalization of histologic lesions with virus antigen expression confirms that the lesions are caused by influenza virus infection. The histologic character of the lesions in the ferrets—necrotizing bronchointerstitial pneumonia—are comparable to the diffuse alveolar damage seen in human fatal cases of pneumonia from seasonal human influenza virus infection. 4 Therefore, this is a suitable animal model for pneumonia caused by seasonal influenza in humans. In the meantime, seasonal H1N1 has been replaced by pandemic (H1N1) 2009. We have shown that pandemic (H1N1) 2009 caused a more severe pneumonia in ferrets than seasonal H1N1 influenza virus but not as severe as highly pathogenic avian influenza virus H5N1. 15
In the present experiment, virus antigen expression was detected in the alveolar epithelial cells as well as the bronchiolar and bronchial epithelial cells. In human cases of seasonal influenza infection, there was rarely virus antigen expression found in the alveolar epithelial cells. 3 Possible explanations for this difference in antigen expression between humans and ferrets include the direct intratracheal inoculation route8,15 with the high doses of virus in the ferrets, the later stage of disease at which the lung sample is usually taken in humans, 7 the naïve immune status for influenza in ferrets when compared to humans, and a possible difference in tropism.
Our results did not show a difference in severity of lesions between H1/78 and H1/07 or between H3/68 and H3/08. This means that this pneumonia model functions as well for recent and older strains of seasonal influenza viruses. The high dose of the inocula seems to overcome the differences in disease outcomes among different subtypes of the virus, which is visible in lower doses as demonstrated by Svitek et al. 13
The viral titers in the brain of few ferrets, albeit low, did not correlate with the results of histopathology and immunohistochemistry. Although we did find a slightly increased number of lymphocytes in the meninges and choroid plexus of few ferrets, there was no viral antigen expression by immunohistochemistry as a demonstration of virus-infected cells. Therefore, we cannot confirm virus replication in the brain. One possible explanation for the presence of virus in the brain could be that it is present in the sample (eg, in the blood or cerebrospinal fluid) and not replicating in the neuronal tissue. Another possible explanation is that virus isolation is more sensitive than immunohistochemistry.
In conclusion, we here demonstrate an animal model for pneumonia from seasonal influenza A virus infection in humans. Regardless of the virus strain used, the results of clinical, pathologic, and virologic analyses were consistent and comparable. This model appears to be suitable for recent and older strains of H1N1 and H3N2 subtypes and should be useful to test vaccines and antiviral agents against pneumonia from these virus infections.
Footnotes
Acknowledgements
We thank C. van Hagen, R. van Lavieren, L. Leijten, P. van Run, and T. Bestebroer for technical assistance; A. Gomersbach and W. Vos for biotechnical assistance; D. van de Vijver and L. Reperant for statistical assistance; and F. van der Panne for figure preparation.
K.J.S. and J.H.S. are employed full-time by, and G.v.A. and A.D.M.E.O. are partly employed by, ViroClinics Biosciences BV.
This study was supported financially by TI Pharma (grant T4-214) and ERC project FLUPLAN (250136).