
Abstract
A novel swine-origin H1N1 influenza A virus has been identified as the cause of the 2009 influenza pandemic in humans. Since then, infections with the pandemic (H1N1) 2009 influenza virus have been documented in a number of animal species. The first known cases of lethal respiratory disease associated with pandemic (H1N1) 2009 influenza virus infection in house pets occurred in domestic cats in Oregon. A 10-year-old neutered domestic shorthair and an 8-year-old spayed domestic shorthair died shortly after developing severe respiratory disease. Grossly, lung lobes of both cats were diffusely firm and incompletely collapsed. Histologically, moderate to severe necrotizing to pyonecrotizing bronchointerstitial pneumonia was accompanied by serofibrinous exudation and hyaline membranes in the alveolar spaces. Influenza A virus was isolated from nasal secretions of the male cat and from lung homogenate of the female cat. Both isolates were confirmed as pandemic (H1N1) 2009 influenza virus by real-time reverse transcriptase polymerase chain reaction. With immunohistochemistry, influenza A viral antigen was demonstrated in bronchiolar epithelial cells, pneumocytes, and alveolar macrophages in pneumonic areas. The most likely sources of infection were people in the household with influenza-like illness or confirmed pandemic (H1N1) 2009 influenza. The 2 cases reported here provide, to the best of the authors’ knowledge, the first description of the pathology and viral antigen distribution of lethal respiratory disease in domestic cats after natural pandemic (H1N1) 2009 influenza virus infection, probably transmitted from humans.
The first reports of a new swine-origin influenza virus came from unrelated human cases in California in March and April 2009 and were subsequently linked to an outbreak in Mexico. 4,6,7 The isolated virus was identified as a novel quadruple reassortant swine-origin influenza A virus (SOIV), or pandemic (H1N1) 2009 influenza virus. 8,19 Genetic analysis of the novel virus and recent influenza A virus isolates from humans, birds, and pigs revealed that genome sequences are traceable to well-established swine influenza lineages. 8,27
Influenza A viruses have only recently been recognized as possible causes of respiratory disease in cats. 12,14,26,28 Highly pathogenic avian influenza virus (HPAIV) H5N1 has been documented to successfully infect felids. 14,31 Natural and experimental infections of domestic cats with HPAIV H5N1 lead to bronchointerstitial pneumonia with serofibrinous to neutrophilic alveolar exudate and can result in death. 12,14,26,28
Identification of pandemic (H1N1) 2009 influenza virus in animals—namely, pigs—was first reported in May 2009 from Canada 20,34 and since then from other continents. The host species spectrum has expanded to include diverse animals, from birds (turkeys) to exotic mammals (cheetah). 24 The first pandemic (H1N1) 2009 influenza virus infection in house pets was diagnosed in ferrets in Oregon and Nebraska, followed by domestic cats in Iowa, Utah, and Oregon. 24,30 The first known reported pandemic (H1N1) 2009 influenza virus-associated mortalities of house pets occurred in 2 domestic cats in Oregon and are the subject of this report.
Materials and Methods
Case Histories and Samples
Case No. 1, a 10-year-old neutered domestic shorthair from Oregon, was examined November 4, 2009 (day 1), by a veterinarian for labored breathing of 1 day’s duration and for vomiting the morning of presentation. The cat was an indoor/outdoor cat with an unknown vaccination status and without previous health problems. On physical examination, the cat had a body temperature of 101.7°F and a rapid respiratory rate, with shallow breathing and inspiratory and expiratory noises. No coughing or sneezing was reported or present during the examination. Thoracic radiographic findings were consistent with pneumonia with caudal lobe air bronchograms and mixed interstitial alveolar pattern. The cat was treated with amoxicillin trihydrate/clavulanate potassium, 15.6 mg/kg per os (PO) twice daily, and sent home. The cat was presented again the following day (day 2) because of worsening of dyspnea and was subsequently placed on oxygen therapy, with oxygen running through a distilled water tank and then into a cage (28 × 28 × 28 in. [71 × 71 × 71 cm]) at a flow rate of 0.5 liters per minute. The cat was also treated with terbutaline sulfate (0.01 mg/kg, subcutaneously [SQ]) and enrofloxacin (5 mg/kg, intramuscularly [IM]). At the end of the day, the cat had an improved respiratory rate and appetite and was released with enrofloxacin (2 mg/kg PO twice daily) and terbutaline sulfate (0.07 mg/kg PO twice daily). On day 3, the cat was again presented for increased dyspnea and cyanosis. The cat was hospitalized and received oxygen therapy, terbutaline sulfate (0.01 mg/kg SQ), and amoxicillin trihydrate/clavulanate potassium (15.6 mg/kg PO) along with enrofloxacin (2 mg/kg PO). A second radiograph depicted consolidation of the ventral aspect of the cranial lung lobes. Owing to the considerable worsening of radiographic changes, dexamethasone sodium phosphate (0.4 mg/kg IM), terbutaline sulfate (0.01 mg/kg IM), enrofloxacin (5 mg/kg IM), and ampicillin (6 mg/kg SQ) were added, and oxygen therapy was continued. The cat died the next day (day 4).
A child in the household had been sick with severe influenza-like illness, suspected but not confirmed to be an H1N1 infection, approximately 1 week before the cat’s illness. Four other cats in the household became ill with variably severe clinical signs of respiratory disease but recovered. Gross examination of the thoracic cavity of cat No. 1 by the practitioner confirmed pneumonia affecting about 75% of the lung field, with no gross lesions in other organs. Formalin-fixed lung tissue from cat No. 1 plus nasal swabs from cat No. 1 and 2 of the 4 cats in the household with previous clinical signs of respiratory disease were submitted to the Veterinary Diagnostic Laboratory (VDL) at Oregon State University. A deep bronchial swab taken at necropsy of cat No. 1 was submitted to a commercial diagnostic laboratory and did not yield any bacterial growth. One cat in the household never developed influenza-like illness. Other than the 5 cats, no other pets lived in the household.
Case No. 2, an 8-year-old spayed domestic shorthair, presented November 24, 2009, as an emergency to a veterinary clinic with a complaint of severe weakness, painfulness, and inappetence. The cat had a history of chronic upper respiratory disease with sneezing, nasal discharge, and chronic sinusitis, attributed by the caretaker to allergies. Vaccination status was unknown, and the cat had not been tested for feline immunodeficiency virus or feline leukemia virus infection. Physical examination findings included rectal temperature < 96°F, 7% dehydration, recumbency, nasal and ocular discharge, sneezing, cyanotic mucous membranes, dental tartar and gingivitis, and occasional vocalization. Consolidation of cranial lung lobes, more severe in the left lung, and pleural effusion were evident in thoracic radiographs. A nasal swab in saline was frozen at –20°C for influenza viral testing. The cat was treated with subcutaneous saline (80 ml/kg, 0.9% NaCl) and oseltamivir phosphate (4.8 mg/kg PO) but died the same day. Upon gross examination of the thoracic and abdominal cavity, the practitioner found no lesions except pneumonia and swollen bronchial lymph nodes. Samples of fresh lung and formalin-fixed lung and bronchial lymph nodes were submitted to the VDL at Oregon State University.
The owner of cat No. 2 had a 10-day history of severe respiratory disease, during which time she had handled and cared for the cat at home. By the time cat No. 2 was presented to the veterinary clinic, the owner had been hospitalized, and an infection with pandemic (H1N1) 2009 virus had been confirmed. The only other pets on the premises were llamas.
Histopathology and Immunohistochemistry
Two pieces of lung from cat No. 1 were received in formalin; 3 sections were submitted for histologic analysis. Three pieces of lung and a bronchial lymph node from cat No. 2, received in formalin, were trimmed into 5 sections of lung and 2 sections of lymph node for histologic analysis. Tissues were routinely processed for light microscopy. Briefly, sections of formalin-fixed, paraffin-embedded tissues were deparaffinized and stained with HE, Brown–Hopps Gram stain, and Warthin–Starry silver impregnation (3 μm) or processed for immunohistochemistry (5 μm).
Immunohistochemistry was performed on an autostainer (Dako Autostainer Universal Staining System, Dako, Carpinteria, CA) with Nova Red chromogen (Vector Laboratories, Burlingame, CA) and Mayer’s hematoxylin counterstain (Sigma, St. Louis, MO). Histologic sections were high-temperature antigen retrieved with BD Retrieval A solution (Dako). To detect viral antigen, a goat anti-influenza A polyclonal antibody (ab20841 against strain USSR-H1N1, Abcam Inc, Cambridge, MA; diluted 1:300) and rabbit anti-goat secondary antibody (Vector; diluted 1:1000) were used. Sections of a formalin-fixed, paraffin-embedded pellet of Madin–Darby canine kidney (MDCK) cells infected with a pandemic (H1N1) 2009 influenza virus isolate served as positive control. Sections from a pellet of uninfected MDCK cells, normal cat lung, and serial sections of lung from cat Nos. 1 and 2 incubated with nonimmune serum served as negative controls. The primary antibody for cytokeratins were the monoclonal mouse antibody cocktail AE1/AE3 (Dako; diluted 1:200) and a polyclonal rabbit broad-spectrum cytokeratin antiserum (Dako; diluted 1:500). Sections of normal cat lung and skin served as positive control tissues; serial sections of these tissues incubated with nonimmune serum served as negative controls.
Virology
The postmortem nasal swabs from cat No. 1 and antemortem swabs from 2 cats in the same household that recovered from respiratory disease were submitted for polymerase chain reaction (PCR) analysis and virus isolation. Fresh lung tissue was not submitted from this case. A nasal swab in saline (stored at –20°C and submitted frozen) and a piece of fresh lung from case No. 2 were submitted for PCR analysis and virus isolation. For virus isolation, saline containing nasal swabs was diluted with an equal volume of Dulbecco’s minimal essential medium (MEM) supplemented with penicillin (100 U/ml), streptomycin (100 μg/ml), gentamicin (50 μg/ml), and amphotericin-B (2.5 μg/ml) and stored at 4°C until inoculated. The homogenate of lung from cat No. 2 was prepared as an approximately 10% suspension in similar Dulbecco’s MEM. Specimens were centrifuged to pellet the cell debris, and supernatants were stored at 4°C until inoculated. For real-time reverse transcriptase PCR (rRT-PCR), saline containing nasal swabs and lung homogenates in MEM plus antibiotics were extracted using commercially available extraction kits (Ambion MagMax AI/ND Viral RNA Isolation Kit, Ambion Inc, Austin, TX).
For initial screening for influenza virus, rRT-PCR assays designed to detect the influenza A matrix gene 29 and the neuraminidase 1 gene specific for the 2009 pandemic H1N1 influenza A virus were done at the VDL at Oregon State University, which is a member of the National Animal Health Laboratory Network and performed the assays according to the standardized protocol provided by the National Veterinary Services Laboratories (NVSL). 30 Virus isolation was performed at the VDL in MDCK cell cultures with procedures specified by the OIE (World Organisation for Animal Health) for swine influenza virus. 21
For confirmation of virus identification by rRT-PCR and sequencing, the nasal secretions from cat No. 1 were submitted to the NVSL in Ames, Iowa. 30 The lung homogenate from cat No. 2 was submitted to the Center for Advanced Host Defense Immunobiotics and Translational Comparative Medicine (CAHDIT) and the VDL at Iowa State University for confirmatory testing by molecular screening and typing for influenza A and the pandemic (H1N1) 2009 influenza A virus. 30
To detect other feline respiratory viruses, virus isolation was performed on the nasal swab from cat No. 1 and the nasal and lung samples from Cat No. 2. Crandell feline kidney cell cultures (CRFK) were prepared in 12.5-cm2 tissue culture flasks in MEM containing 10% gamma-irradiated fetal bovine serum (FBS). Media from cell cultures at 90% to 100% confluency were removed, and cells were inoculated with 0.5 ml prepared supernatant and adsorbed for 60 minutes at 37°C. Following adsorption, the cells were rinsed once with serum-free MEM, then incubated in MEM containing 5% gamma-irradiated FBS plus penicillin, streptomycin, gentamicin, and amphotericin-B. Cultures were observed for 5 to 7 days for visible cytopathic effect and, if negative, were blind-passaged for an additional 5 to 6 days on fresh CRFK cell cultures. Following 2 passages, cells were scraped and cell smears examined for feline herpesvirus 1 and feline calicivirus by direct fluorescent antibody technique. 1,32
Results
Gross Pathology
The 2 formalin-fixed pieces of lung from cat No. 1 were diffusely firm, dark, and incompletely collapsed. One piece had a focus (1.0 × 0.5 cm) of pleural thickening. On cut section, one piece had an enhanced peribronchiolar pattern (Fig. 1 ).
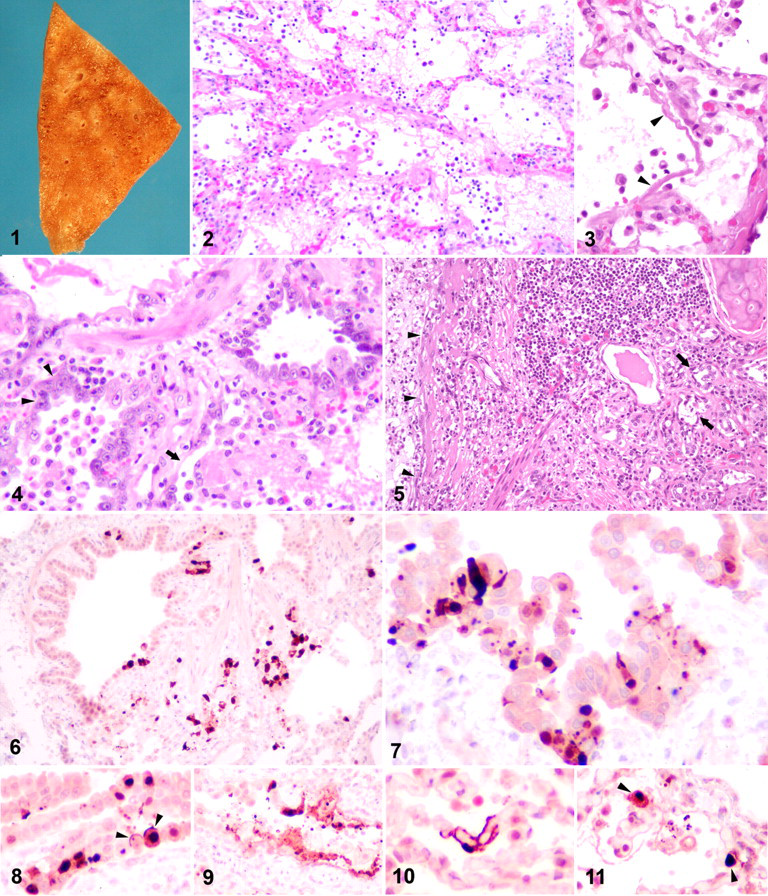
Three pieces of formalin-fixed lung and a lymph node were received from cat No. 2. Two pieces of lung were diffusely firm, dark, and incompletely collapsed: One piece had multiple pinpoint off-white subpleural foci (interpreted as alveolar histiocytosis); the other piece had multifocal pleural pallor and thickening (pleural fibrosis). The lymph node was enlarged with a fine multinodular pattern on capsular and cut surfaces, interpreted as lymphoid hyperplasia.
Histopathology and Immunohistochemistry
Cat No. 1 had severe, diffuse, acute to subacute bronchointerstitial fibrinonecrotizing pneumonia (all sections) and multifocal, severe type II pneumocyte hyperplasia of peribronchiolar alveoli (1 section). Lesions in all sections were centered on terminal airways. Most bronchioles had complete or segmental epithelial necrosis and rare multinucleated cells. Denuded areas were covered with fibrin, and bronchiolar lumens contained a few macrophages, sloughed epithelial cells, and scanty cellular debris (Fig. 2 ). Remaining bronchiolar epithelium was attenuated or appeared normal on light microscopy. Diffusely alveolar spaces were flooded with wispy to dense protein-rich material (Fig. 2), hyaline membrane formation (Fig. 3 ), a variable number of large round cells with moderate amounts of cytoplasm (macrophages and desquamated type II pneumocytes), scattered neutrophils, and rare multinucleated giant cells. In 1 section, most peribronchiolar alveoli were lined with cuboidal epithelium (type II pneumocyte hyperplasia) or large, sometimes multinucleated cells, and they had segmental epithelial necrosis and dense fibrin accumulations (Fig. 4 ). Bronchi had multifocal, mild lymphoplasmacytic to histiocytic infiltrates in the lamina propria and epithelial degeneration in rare bronchial glands. Diffuse, mild to moderate perivascular edema and mild, multifocal pleural mesothelial hypertrophy and hyperplasia were also present.
Cat No. 2 had moderate to severe, multifocal, acute necrotizing to mucopurulent bronchointerstitial pneumonia superimposed on severe, multifocal chronic bronchitis and focally extensive bronchiolitis obliterans. The bronchial lymph node had marked lymphoid hyperplasia with focal, mild pyonecrotizing lymphadenitis. The lesions in airways included segmental to diffuse epithelial necrosis in rare bronchi and many bronchioles, loose luminal accumulation of mucus with scattered neutrophils, and mild to severe lymphoplasmacytic infiltration of bronchial submucosa (Fig. 5 ). Multifocally, epithelial necrosis and neutrophils extended into bronchial glands (Fig. 5). There was a mild to moderate, patchy accumulation of macrophages and neutrophils with mild fibrin exudation in alveolar spaces. Lesions restricted to 1 or 2 samples included multifocal, severe chronic bronchitis and bronchiolitis with massive peribronchiolar fibrosis, atrophy of bronchial glands, squamous epithelial metaplasia, and scattered granulomas with rare acicular clefts (presumptive cholesterol clefts) obliterating lumens; focally extensive, suppurative alveolitis and bronchiolitis; multifocal, severe alveolar edema and moderate emphysema; and multifocal, moderate pleural and interstitial fibrosis. Two sections of lymph node had prominent lymphoid hyperplasia with secondary follicles. Medullary and subcapsular sinuses were filled with moderate numbers of macrophages and scattered intact and degenerated neutrophils and, focally, with degenerated neutrophils, cellular debris, and fibrin. Bacteria were not detected with HE, Gram stain, or Warthin–Starry silver impregnation in serial sections of lung or lymph node.
Immunohistochemistry for influenza viral antigen resulted in a homogeneous to granular, red to dark red, nuclear and cytoplasmic signal that often obscured cellular detail. In both cats, a few of the bronchioles with intact epithelium had individual or small groups of immunoreactive, intact or necrotic, nonciliated epithelial cells flanked by negative, histologically normal epithelial cells (Figs. 6, 7 ). Intense immunoreactivity occasionally labeled the apical surface of nonciliated bronchiolar epithelial cells (Fig. 8 ). Bronchioles that lacked epithelial lining had segmental, irregular, linear to clumped immunoreactivity along the denuded surface (Fig. 9 ). Cellular debris in some bronchioles and alveolar spaces contained dark red to almost brown globules. Peribronchiolar alveoli with type II pneumocyte hyperplasia had individual to small groups of immunoreactive cells interpreted as type II pneumocytes. In areas with acute alveolar damage, intense immunoreactivity labeled the surface of short segments of interalveolar septa, filled the nucleus and cytoplasm of flat lining cells (Fig. 10 ), or obscured cytologic detail of large round cells in alveolar spaces or interalveolar septa (Fig. 11 ). Based on their distribution and shape, the immunoreactive elongated flat cells lining alveolar spaces were interpreted as type I pneumocytes. The round cells in alveolar spaces that were immunohistochemically positive for influenza viral antigen did not react with antibodies against low– or high–molecular weight cytokeratins in serial sections and were interpreted as alveolar macrophages. Nonetheless, the possibility cannot be ruled out that some of the cells may have been infected and desquamated pneumocytes. Rarely, the epithelium of bronchial glands had a few cytoplasmic dark red granules. This pattern was more abundant in the hyperplastic glands of cat No. 2. Viral antigen was not detected in bronchial epithelium of either cat.
Virology
Nasal secretions from cat No. 1 and lung homogenate from cat No. 2 tested positive for the influenza A matrix gene 21 and neuraminidase 1 gene of pandemic (H1N1) 2009 influenza virus by rRT-PCR at the VDL at Oregon State University. 30 Cycle thresholds for the matrix and neuraminidase 1 genes were 29.0 and 30.74 for case No. 1 and 20.0 and 22.22 for case No. 2, respectively. Nasal swabs from the other 2 cats (recovered) from the household of cat No. 1 and the nasal swab from cat No. 2 tested negative.
Virus isolation from the nasal swab of cat No. 1 and lung homogenate from cat No. 2 yielded an influenza virus. The positive samples were confirmed to contain pandemic (H1N1) 2009 influenza virus by the NVSL in Ames, Iowa, or the VDL and CAHDIT at Iowa State University. 30 No other viruses were isolated. Neither feline herpesviral nor feline caliciviral antigen was detected by indirect immunofluorescence in smears from scraped cell cultures.
Discussion
Historically, cats have been considered relatively resistant to influenza virus infections. 9 This view has been modified by recent reports of respiratory and systemic disease in domestic cats after natural and experimental infections with HPAIV H5N1. 12,14,26,28 Cats infected with HPAIV H5N1 develop bronchointerstitial pneumonia with epithelial necrosis and fibrin exudation in bronchioles and alveoli. 12,14,26,28 Necrotizing bronchointerstitial pneumonia is also a consistent finding in cynomolgus monkeys, BALB/c mice, miniature pigs, and ferrets after experimental and natural infections with pandemic (H1N1) 2009 influenza virus 10,17 and in human patients with lethal SOIV infections. 15,23 Both cats in this report had necrotizing bronchointerstitial pneumonia. Feline herpesvirus and calicivirus are the main infectious causes for acute bronchointerstitial pneumonia in cats 2 ; neither virus was detected from the 2 cases presented here.
Extension of viral infections and lesions to alveoli is characterized by alveolitis and alveolar flooding with serofibrinous to neutrophilic exudate. These inconsistent but clinically important findings in influenza vary with the host species, specific virus involved, and route of infection. 10,12,15 –17 Domestic cats naturally infected with HPAIV H5N1, 12 experimental animals infected with pandemic (H1N1) 2009 influenza virus, 17 and human patients with lethal pneumonia based on pandemic (H1N1) 2009 influenza virus all develop severe alveolar lesions. Both cats of this report developed alveolitis, albeit with variable severity and histologic features. Cat No. 1 had severe, diffuse alveolitis dominated by abundant fibrin exudation, occasional hyaline membranes, numerous macrophages, and fewer desquamated pneumocytes, whereas cat No. 2 had a more restricted distribution of alveolar lesions with less serofibrinous exudation that included neutrophils. Aside from viral infections, gastric acid, highly concentrated oxygen (85 to 100%), and hyperventilation cause severe acute alveolitis secondary to damage of capillary endothelium and/or pneumocytes. Interestingly, cat No.1, but not cat No. 2, received oxygen therapy at a routine flow rate. Neither cat had a history of vomiting. It is conceivable that the virus-induced alveolar damage in cat No. 1 was exacerbated by increased alveolar partial oxygen pressure and/or hyperventilation. Alternatively, differences in host or pathogen-dependent factors could have contributed to the difference in alveolar damage between the 2 cats. Both cats had an unknown feline immunodeficiency virus and feline leukemia virus infection status. A more detailed analysis of the pandemic (H1N1) 2009 influenza virus isolated from the 2 cats is pending.
Demonstration of viral antigen within lesions supports a causal association between pathogen and lesion. In both cases presented here, immunohistochemistry with an antibody specific for influenza A viral antigen resulted in strong labeling of bronchiolar epithelium, with focal reaction of alveolar epithelium, rounds cells in alveolar spaces, and, rarely, bronchial glands. Consistent with the histologic lesions, alveolar immunoreactivity was more abundant and widely distributed in cat No. 1 than in cat No. 2. Based on distribution, shape, and immunohistochemical expression of cytokeratins, the influenza virus–immunoreactive flat, elongated cells lining alveoli and the round cells in alveolar spaces were interpreted as type I pneumocytes and macrophages, respectively. However, it is possible that some of the luminal cells immunoreactive for influenza viral antigen were desquamated type II pneumocytes. The immunohistochemical labeling of the surface of bronchiolar and alveolar epithelium with antibody to influenza A virus is reminiscent of the labeling pattern with lectins for detection of influenza virus receptors 13,35 and might reflect virus bound to or virus budding from the surface of the airway and alveolar epithelium. 18,25 In the case of alveolar epithelial cells, it cannot be completely excluded that this represents cytoplasmic immunoreactivity of portions of pneumocyte cell bodies. Immunoelectron microscopy would be required to confirm the location of the signal at a subcellular level; this was not done, because only formalin-fixed, paraffin-embedded tissue was available. The coating of denuded surfaces of bronchioles and alveoli with small amounts of debris heavily laden with viral antigen was observed in both cats and could have served as a source of virus for intrapulmonary spread of the infection.
Interestingly, neither prominent necrosis nor viral antigen was detected in bronchial epithelium in either cat. This is in contrast to the necrotizing bronchitis with intralesional viral antigen seen in some human patients with lethal pandemic (H1N1) 2009 influenza virus–associated pneumonia 15 and in ferrets experimentally infected with pandemic (H1N1) 2009 influenza virus. 17 The distribution of lesions and influenza viral antigen throughout the respiratory tract correlates with the distribution pattern of sialic acid molecules on cell surfaces of the host. 33 For example, human influenza viruses preferentially bind to alpha-2,6-linked sialic acids, which are abundant in the human trachea, whereas avian influenza viruses bind preferentially to alpha-2,3-linked sialic acids, which are present mainly on pneumocytes and only sparsely in the tracheal and bronchial epithelium of humans. 11,35 Interestingly, expression of sialic acid molecules can be altered in influenza virus–infected cells and in metaplastic cells of the respiratory tract. 35 The distribution pattern of viral antigen in the cats presented here resembles the in vitro binding pattern of avian influenza A viral particles to tissues from healthy cats and differs from that of seasonal H1N1 influenza A viral particles to tissues from healthy cats and healthy humans. 33
The type II pneumocyte hyperplasia in cat No. 1 was superimposed with acute epithelial necrosis and fibrin exudation in many areas. Prominent type II pneumocyte hyperplasia occurs as early as 2 to 3 days after damage to the respiratory epithelium with ongoing acute alveolar damage. 2 The course of clinical disease in cat No. 1 was protracted enough that type II pneumocyte hyperplasia could have been a response to the ongoing viral damage rather than a preexisting condition. The focal extensive chronic bronchitis in cat No. 2 is consistent with the cat’s history of chronic respiratory disease and preexisting pulmonary damage. In children, preexisting conditions, especially of the respiratory tract and nervous system, dramatically increase the risk of lethal outcome after infection with pandemic (H1N1) 2009 influenza virus. 5 The situation may be similar in cats.
Bacterial coinfections are identified in less than one-third of lung samples from lethal pandemic (H1N1) 2009 influenza virus infections in humans. 3 Nonetheless, predominance of neutrophils in the bronchioloalveolar exudate in cat No. 2 cannot be taken as clear evidence of a bacterial secondary infection. Even under experimental conditions, a predominantly neutrophilic bronchiolitis and alveolitis develops in the absence of bacterial pathogens in animals infected with influenza A viruses. 16,26 Nonetheless, failure to demonstrate bacteria in Gram-stained and Warthin–Starry silver-impregnated serial lung sections from either cat and from lymph node of cat No. 2 and failure to grow bacteria from the postmortem bronchial swab of cat No. 2 do not rule out secondary bacterial infection.
The negative PCR result for influenza virus on the nasal secretions from cat No. 2 could have multiple causes. The nasal swab was held for 1 week at –20°C before shipment to the VDL, whereas the positive lung sample from the same cat was refrigerated at 4°C. Degradation of RNA during storage could have resulted in false-negative PCR results from nasal secretions. Alternatively, viral concentration in the swab sample may have been too low for detection. Virus replication was limited to the lower respiratory tract in natural HPAIV H5N1 infections in cats. 12 It is also possible that the sample was simply of poor quality.
Transmission of pandemic 2009 (H1N1) influenza virus from humans to cats was recently implicated in the case of a domestic cat that recovered from clinical respiratory disease. 30 A member of the owner’s family in the household of cat No. 1 of this report had symptoms consistent with severe influenza-like illness. The owner of cat No. 2 was hospitalized with pneumonia and confirmed positive for pandemic (H1N1) 2009 influenza virus. Probable horizontal transmission of influenza A virus between felids has been reported in domestic cats and tigers. 26,31 The cats in this report had no known contact with other animals with respiratory disease owing to pandemic (H1N1) 2009 influenza virus, and they lived 99 km apart. Household members were therefore the most likely source of infection. Transmission of an influenza A virus from a human patient to 2 domestic cats has been documented for the Hong Kong pandemic influenza virus (H3N2). 22 A detailed genetic analysis comparing the feline pandemic (H1N1) 2009 isolates of this report with that from the owner of cat No. 2 would be required to prove this point.
In summary, this report provides evidence that domestic cats are at risk of severe respiratory disease and death attributed to pandemic (H1N1) 2009 influenza virus infection. The close spatiotemporal correlation of the cats in this report with severe human influenza-like illness or confirmed human pandemic (H1N1) 2009 influenza virus infection, in the absence of an obvious alternative source of infection, is consistent with human-to-cat transmission of pandemic (H1N1) 2009 influenza virus. Host adaptations of the pandemic (H1N1) 2009 influenza virus isolates from the cases reported here and possible associated changes in virulence have not been characterized.
Footnotes
Acknowledgements
We thank the following institutions and individuals for confirmation of pandemic (H1N1) 2009 influenza virus isolates: for case No. 1, S. Swenson, L. Koster, M. Jenkins-Moore, and M. Killian, US Department of Agriculture, Animal and Plant Health Inspection Service, National Veterinary Services Laboratories, Ames, Iowa; for case No. 2, J. Trujillo at the Center for Advanced Host Defense Immunobiotics and Translational Comparative Medicine and K.-J. Yoon, E. Strait, K. Harman, and L. Bower at the Veterinary Diagnostic Laboratory, College of Veterinary Medicine, Iowa State University (case No. 2). We thank B. Valentine for review of a draft of the article.
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
The authors declared that they received no financial support for their research and/or authorship of this article.