
Abstract
In dogs, there are several idiopathic meningoencephalitides, such as necrotizing meningoencephalitis (NME), necrotizing leukoencephalitis (NLE), and granulomatous meningoencephalomyelitis (GME). Although they are often assumed to be immune mediated, the etiology of these diseases remains elusive. In this study, the histopathology of the lesions caused by these conditions and the inflammatory cell populations produced in response to them were examined among dogs affected with GME, NME, or NLE to understand their pathogeneses. The brain tissues of dogs with NME (n = 25), NLE (n = 5), or GME (n = 9) were used. The inflammatory cells were identified by immunohistochemistry using antibodies against CD3, IgG, CD20, CD79acy, and CD163. In NME and NLE, malacic changes were located in the cerebral cortex, as well as the cerebral white matter and thalamus, respectively. The distribution of the brain lesions in NME and NLE was breed specific. In GME, granulomatous lesions that were mostly composed of epithelioid macrophages were observed in the cerebral white matter, cerebellum, and brainstem. Although the proportions of IgG-, CD20-, and CD79acy-positive cells (B cells) were not significantly different among the GME, NME, and NLE lesions, that of CD3-positive cells (T cells) was increased in GME. In NME and NLE, CD163-positive cells (macrophages) had diffusely infiltrated the cerebral cortex and white matter, respectively. However, in GME, CD163-positive cells accumulated around the blood vessels in the cerebral and cerebellar white matter. The distributions of these lesions were quite different among GME, NME, and NLE, whereas there were no marked differences in the proportions of inflammatory cells.
Keywords
Necrotizing nonsuppurative meningoencephalitis (NME) is an idiopathic inflammatory disease of the canine central nervous system (CNS) that is characterized by prominent necrosis and the infiltration of inflammatory cells, including lymphocytes, plasma cells, and monocytes or histiocytes into the cerebral cortex and/or white matter, hippocampus, thalamus, and leptomeninges. NME has been reported in various canine breeds, including the Pug, 4,9,27,32 Maltese, 14,41,49 Shih-tzu, 51 Papillon, 51 Chihuahua, 20 Pekingese, 8 Yorkshire Terrier, 2,13,29,34,54,56 and French Bulldog. 48 The areas of necrosis and inflammatory cell infiltration are localized in the cerebral cortex and subcortical region in most NME cases, whereas in a few breeds, such as the Yorkshire Terrier and French Bulldog, the lesions are predominately observed in the white matter, and the disease is called necrotizing leukoencephalitis (NLE).
Granulomatous meningoencephalomyelitis (GME) is another idiopathic canine CNS disorder that is characterized by perivascular cuffs composed of lymphocytes, plasma cells, macrophages, and some neutrophils, as well as granulomatous lesions containing epithelioid cells, mainly in the cerebellum and brainstem. 1,15,16,24 –26,51 –53 However, we have observed a few GME cases that did not display prominent granulomatous lesions, perivascular cuffs, or inflammatory cell infiltration, and rare cases involved the cerebral cortex and white matter. Differential diagnoses for NME should therefore be considered.
The etiopathogeneses of NME, NLE, and GME remain unclear, although many previous reports have tried to identify their causes. 2, 17, 36, 39, 42, 45, 47, 51, 52, 55 In a few cases of GME, the involvement of Borna virus 39,57 or West Nile virus 53 was suspected. In NME and NLE cases, the influence of RNA viruses, such as canine distemper virus, herpes virus, and rabies virus, has also been examined.4, 7, 8, 20, 27, 34, 45, 48, 51, 54 However, no viruses were detected by polymerase chain reaction (PCR) in paraffin-embedded brain tissues from dogs with NME, NLE, or GME. 45 Some reports have suggested that these conditions are autoantibody mediated 47,53,55 or T cell mediated. 24,51,52 Although autoantibodies against glial fibrillary acid protein (GFAP) have been detected in the cerebrospinal fluid (CSF) of many dogs with NME, they were not specific to NME and were absent in some cases. 36,47 It is therefore disputed whether autoantibodies against GFAP are a cause or secondary products of NME. Suzuki et al 51,52 reported that CD3-positive T cells predominate in NME and GME lesions and suggested that GME is a form of delayed-type hypersensitivity based on an autoimmune response. 24 Thus, the T cell response may play a key role in NME, NLE, and GME.
NME, NLE, and GME are considered different diseases because of their unique characteristic lesions, localization in the brain, and breed specificities. Therefore, to confirm whether they are different, the dominant inflammatory cell populations of the diseases should be compared. Thus, in the present study, we compared the lesions, lesion locations, and inflammatory cell populations of NME, NLE, and GME.
Material and Methods
Dog brains
The brain tissues of dogs with NME, NLE, and/or GME were obtained during necropsies performed between 1990 and 2009 at our laboratory. The brains from 25 NME, 5 NLE, and 9 GME cases were examined (Suppl. Table S1).
Pathological examinations
Brain tissues were fixed in 10% neutral buffered formalin, and selected tissues were subsequently routinely embedded in paraffin, sectioned at 4 μm, and stained with hematoxylin and eosin (HE), HE–Luxol fast blue (HE-LFB), and toluidine blue (TB) for microscopic examinations. Other fresh brain tissues from 2 dogs with NME and 1 dog with GME were frozen in Tissue-Tek O.C.T. Compound (Sakura Finetek Japan, Tokyo, Japan) and subsequently stored at –80°C.
Immunohistochemistry
For antigen retrieval, deparaffinized sections were autoclaved at 121°C for 15 minutes or digested with proteinase K (1:400; Wako, Osaka, Japan) at 37°C for 30 minutes. In addition, for the CD79acy immunostaining, sections were treated with 1% sodium dodecyl sulfate (SDS) at room temperature for 5 minutes and then treated with hyaluronidase (4000 U/ml; Sigma, St. Louis, MO) at 37°C for 30 minutes. Endogenous peroxidase activity was blocked by incubating the sections with 3% hydrogen peroxide in methanol at room temperature for 5 minutes. For the tissue blocking, the sections were further treated with skimmed milk at 37°C for 40 minutes. The sections were then incubated at 37°C for 1 hour or at 4°C overnight with primary antibodies. The primary antibodies employed are listed in Supplementary Table S2. Thereafter, the sections were incubated with the Envision polymer reagent (DAKO Japan, Kyoto, Japan) or the LSAB reagent (DAKO Japan) at room temperature for 40 minutes. Visualization was performed with 3,3′-diaminobenzidine tetrahydrochloride (Wako), and counterstaining was performed with hematoxylin.
Quantitative data analysis
The numbers of CD3-, IgG-, CD20-, CD79acy-, and CD163-positive cells per 200 mononuclear cells were counted under ×400 magnification in 10 selected fields of perivascular cuff lesions and areas displaying parenchymal infiltration. Then, mean cell percentage and standard deviation values were calculated. Mean values were compared using the Student’s t-test.
Double-labeling immunofluorescence
Six-μm-thick paraffin sections were autoclaved at 121°C for 15 minutes for antigen retrieval and treated with skimmed milk at 37°C for 40 minutes to block nonspecific reactions. The first primary antibodies were employed at 4°C overnight. The first primary antibodies were rabbit polyclonal antibodies against human CD3 (DAKO, Japan) and biotinylated sheep antiserum against canine IgG (American Qualex, San Clemente, CA). The sections were then incubated with the second primary antibody, mouse monoclonal against human glial filament protein (GFP, prediluted; PROGEN, Heidelberg, Germany), at 37°C for 1 hour. The secondary antibody cocktail was employed at room temperature for 1 hour. The cocktail included fluorescein isothiocyanate–labeled goat anti-rabbit IgG (1:200; Vector Laboratories, Burlingame, CA), fluorescein streptavidin (1:200; Vector Laboratories), and Alexa Fluor 594–labeled goat anti-mouse IgG (1:200; Invitrogen, Eugene, OR). Fluorescein streptavidin was used to visualize the first biotinylated canine IgG antibody. The sections were finally counterstained with TO-PRO-3 iodide (1:1000; Invitrogen, Camarillo, CA) at room temperature for 30 minutes, mounted with Vectashield HardSet (Vector Laboratories), and observed using the Zeiss LSM 510 confocal laser scanning microscope.
Results
Breed Specificities
Different from GME, NME and NLE tended to be breed specific and usually occurred in small breeds. Pug, Chihuahua, Maltese, and Papillon dogs were often affected by NME (Suppl. Table S1, Fig. 1). Yorkshire Terriers and some Pugs were more likely to be affected by NLE (Suppl. Table S1, Fig. 3). One Papillon with NME developed malacic lesions in both the cerebral cortex and white matter.
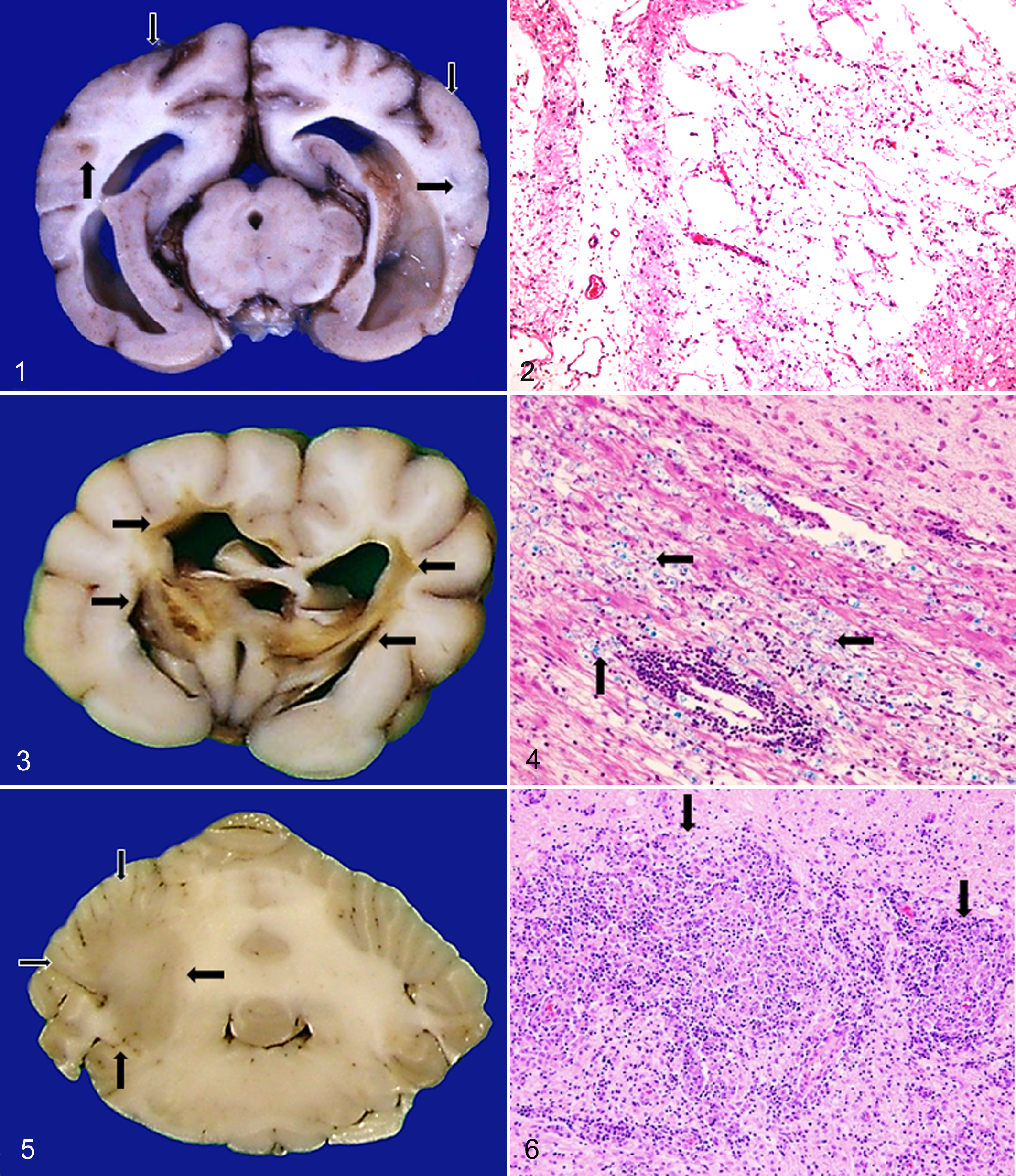
Pathological Changes
NME
Among the microscopic findings of NME, malacic changes and inflammatory lesions were most prominent in the cerebral cortex, hippocampus, and thalamus (Table 1, Suppl. Table S3, Fig. 2). In addition, endothelial hyperplasia or microgliosis was also observed in the capsula interna, nucleus geniculatus medialis, nucleus caudatus, crus cerebri, pons, nuclei habenulares, nucleus pretectalis, and the cerebral cortex in several cases.
Distribution of Lesions in NME, NLE, and GME
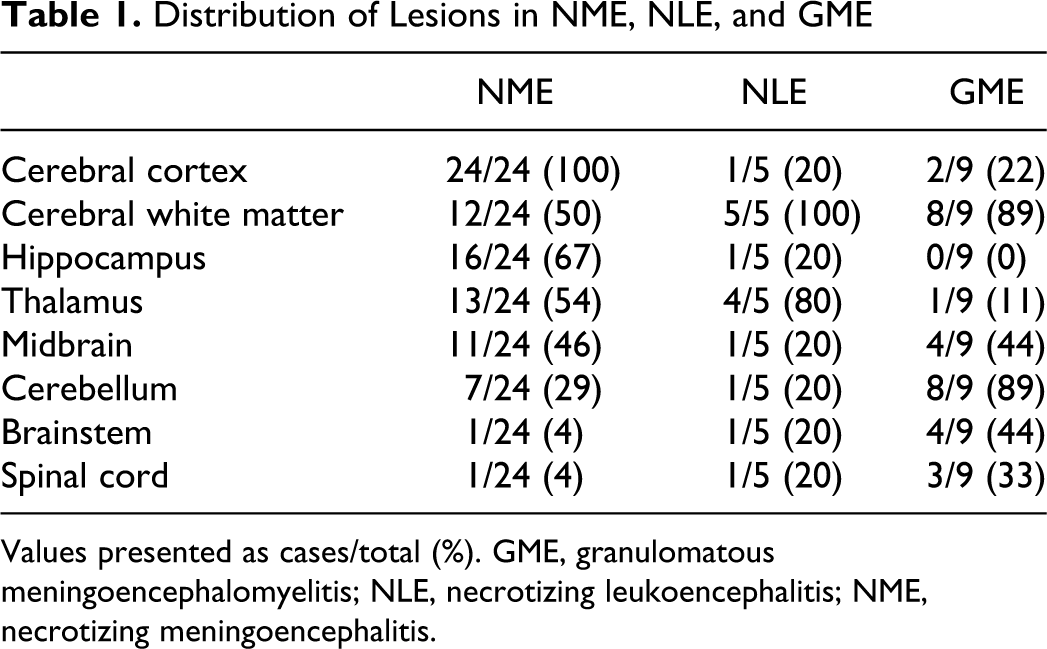
Values presented as cases/total (%). GME, granulomatous meningoencephalomyelitis; NLE, necrotizing leukoencephalitis; NME, necrotizing meningoencephalitis.
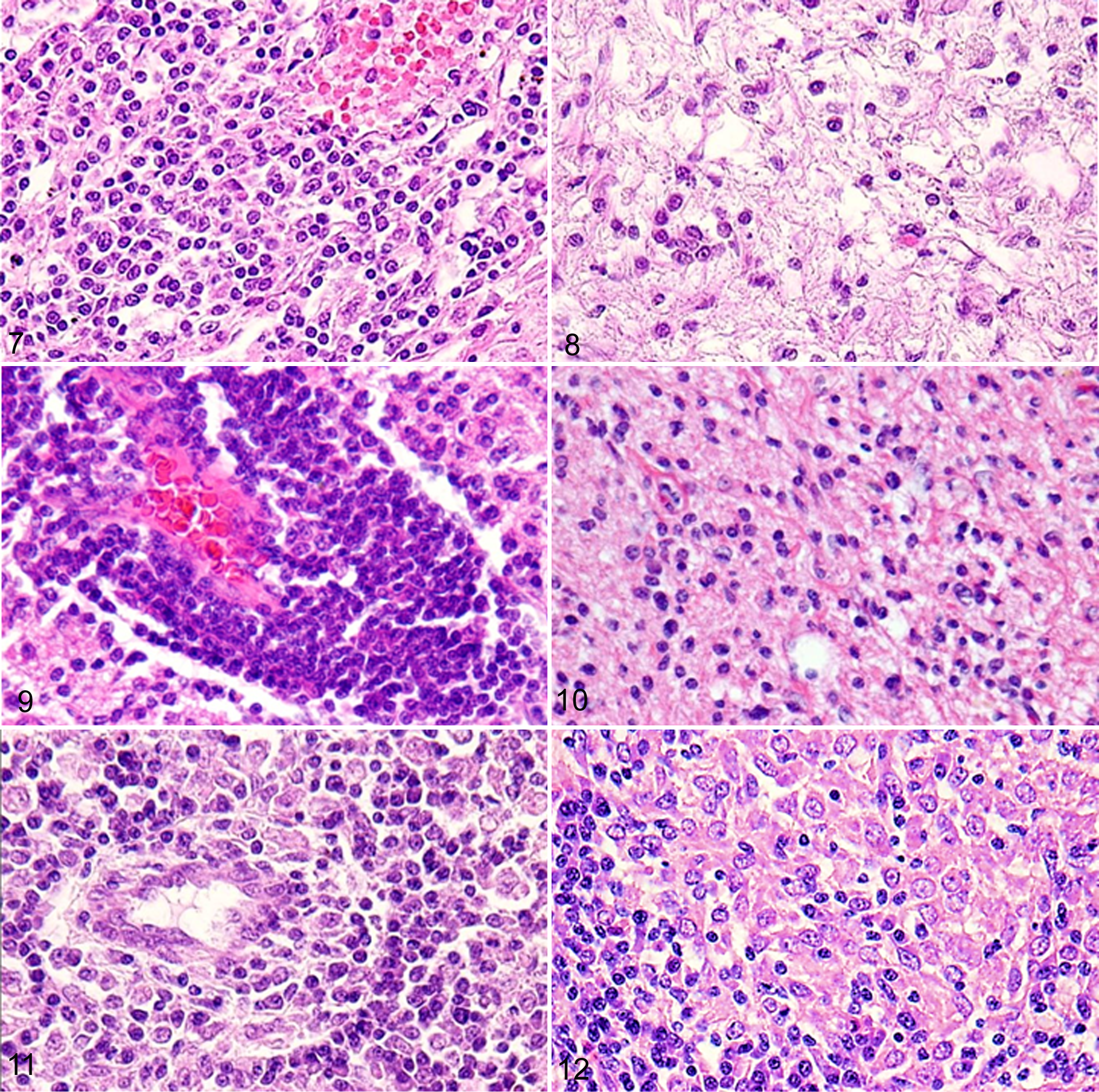
The histopathological lesions of the NME dogs examined were divided into 3 phases: acute, subacute, and chronic. The acute-phase lesions were mainly composed of regions of mild inflammatory cell infiltration. Moderate malacic changes and intense inflammatory reactions were observed in the subacute phase (Figs. 7, 8). In the chronic phase, extensive malacic changes predominated. Areas of leptomeningeal infiltration and perivascular cuffs were most commonly observed in the subacute phase, but they were also found in the chronic phase accompanied with extensive necrosis. The inflammatory reactions progressed continuously throughout the progression of the disease.
NLE
Lesions were found in the thalamus of the majority of NLE cases and in the hippocampus, mesencephalon, cerebellum, and spinal cord in a few cases (Table 1, Suppl. Table S3). The capsula interna was affected in 1 case.
Malacic changes were specifically found in the cerebral white matter and subcortical region (Fig. 3), and inflammatory lesions (Figs. 4, 9, 10) were located in similar regions to those in which they were found in NME. Different from NME, the subleptomeningeal lesions and astrogliosis were mild to moderate. Inflammatory cell infiltration was prominent in the ependyma and mild in the choroid plexus. Vacuolar changes, swollen or fragmented myelin fibers, and spheroid formation were found in the peripheral malacic regions.
GME
Characteristically angiocentric or nodular granulomatous lesions (Figs. 6, 11, 12) composed of macrophages, epithelioid cells, lymphocytes, plasma cells, and neutrophils were observed in the cerebellum, medulla oblongata, and spinal cord (Table 1, Suppl. Table S3). Lesions were detected in the cerebral white matter, especially in subcortical regions, such as the mesencephalon and thalamus (Fig. 5).
Perivascular cuff lesions as well as those involving leptomeningeal infiltration, astrogliosis, and microgliosis were multifocal and asymmetric. The perivascular cuffs consisted of lymphocytes, plasma cells, macrophages, neutrophils, and hypertrophic endothelial cells. Areas of leptomeningeal infiltration were prominent in the cerebellum, brainstem, and spinal cord, especially in the sulci, but mild in the cerebrum. Astrogliosis and microgliosis were frequently detected in the active and severe inflammatory lesions. Malacic lesions were observed in a few cases, which were milder than those found in NME.
Quantification of Inflammatory Cells
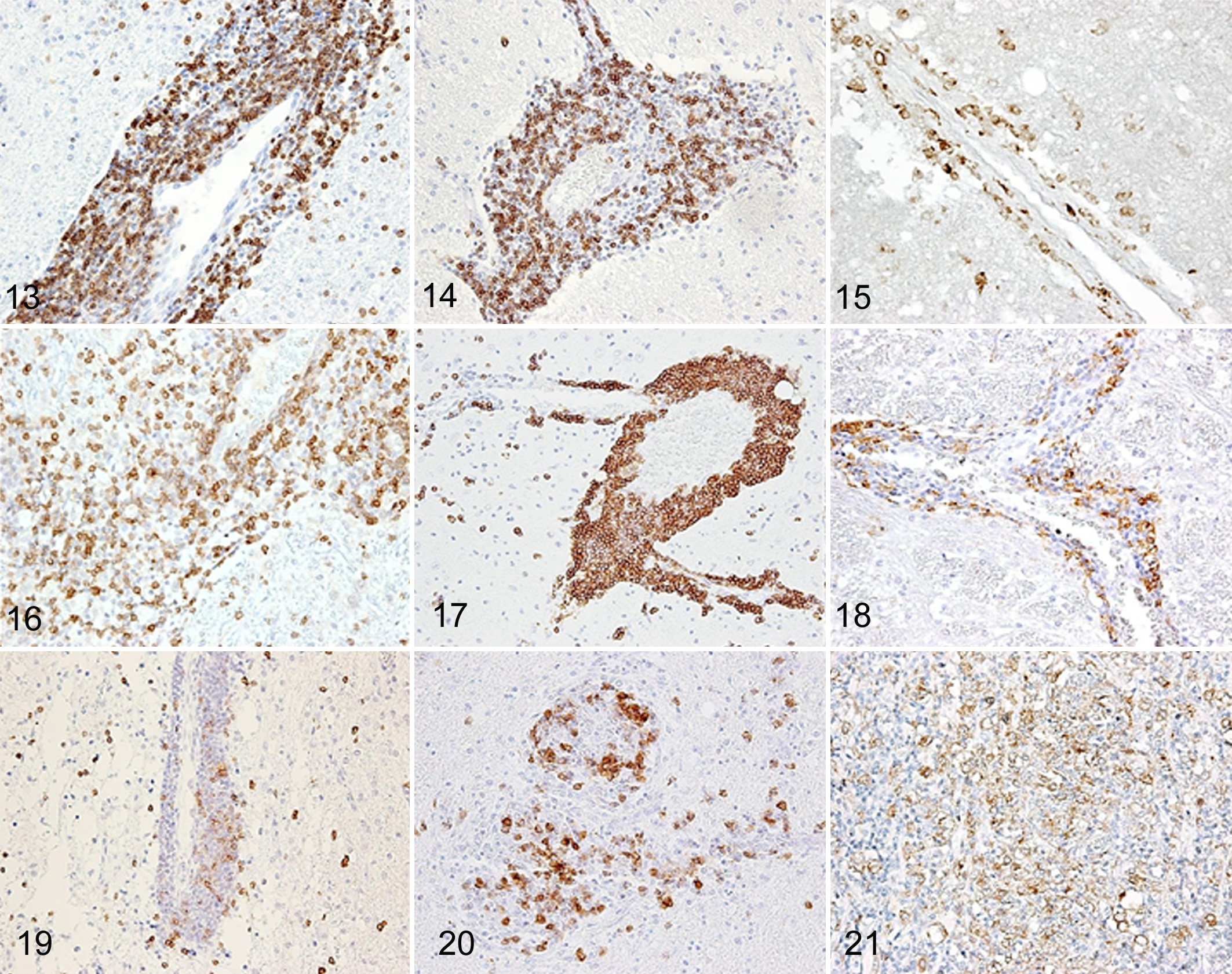
Immunohistochemistry was used to quantify the number of inflammatory cells as illustrated in Figs. 13 to 22. CD3-positive T cells in the perivascular cuffs, neuroparenchyma, and leptomeningeal lesions were predominant among the inflammatory cell populations in the order of GME, NLE, and NME, and the proportions of these cells were significantly different between the diseases (Figs. 13, 16, 19, 22). However, there were few CD3-positive T cells in the malacic regions of NME and NLE.
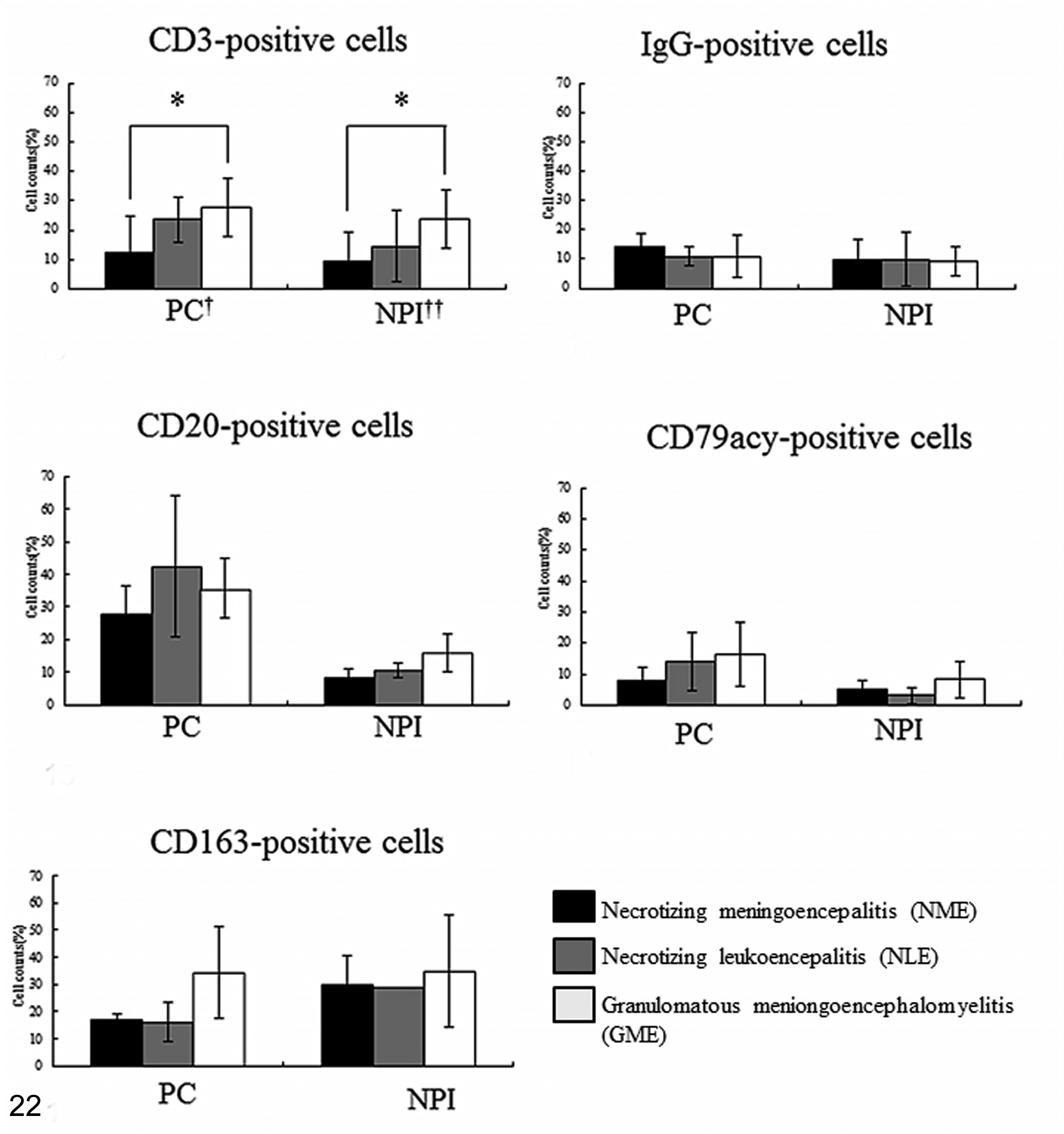
The percentages of CD3-, IgG-, CD20-, CD79acy-, and CD163-positive cells (mean ± SEM, %) in the perivascular cuffs (PC) and neuroparenchymal infiltration (NPI) in animals with NME, NLE, and GME (*P < .01).
In this study, IgG, CD20, and CD79acy were used as B cell markers. CD20 is present in the cell membrane of late pro–B cells through memory cells, but not plasma cells, whereas IgG is present in the cytoplasm of plasma cells. CD79acy is expressed in the cell membrane throughout the B cell life cycle, as well as active plasma cells. The proportion of IgG-positive plasma cells and CD20- and CD79acy-positive B cells was not significantly different (Figs. 14, 17, 20, 22). CD20-positive B cells were more frequently observed in the perivascular cuffs than in the neuroparenchyma, whereas IgG-positive plasma cells and CD79acy-positive B cells infiltrated similarly in the perivascular cuffs and in the neuroparenchyma (Figs. 14, 17, 20, 22).
We then used CD163 as a macrophage and epithelioid cell marker as it is slightly more specific than other antimacrophage antibodies such as antimyeloid/histiocyte antigen (MAC387), lysozyme, Iba-1, and macrophage (HAM56). CD163-positive macrophages were observed more frequently in the GME lesions, particularly in the perivascular cuffs, than in those observed in NME and NLE (Figs. 15, 18, 21, 22). In NME, NLE, and GME, CD163-positive macrophages were localized in the active inflammatory lesions such as the perivascular cuffs and areas of neuroparenchymal infiltration (Figs. 15, 18, 21). In NME and NLE, they were mainly found in the cerebral cortex and white matter, particularly in necrotic lesions, and rarely in the cerebellum, brainstem, and spinal cord. On the other hand, in GME, they were mostly observed in the cerebellum, brainstem, spinal cord, and cerebral white matter. CD163-positive epithelioid cells were specifically detected in the granulomatous lesions of GME but not NME or NLE (Fig. 21).
The proportion of myeloperoxidase-positive neutrophils was approximately 5% to 10%, although no suppurative lesions were found in NME, NLE, or GME (data not shown). Neutrophils were mainly located in the necrotic, perivascular cuff, neuroparenchymal, and leptomeningeal lesions, and they were also frequently observed in the granulomatous GME lesions.
Double-Labeling Immunofluorescence
CD3 and GFAP
In the NME lesions, many CD3-positive T cells (green) adhered to GFAP-positive astrocytes (red) in the peripheral malacic regions of the cortex (Fig. 23). However, in the NLE and GME lesions, CD3-positive T cells had infiltrated among the fibrous processes of GFAP-positive astrocytes in the white matter (Fig. 25).
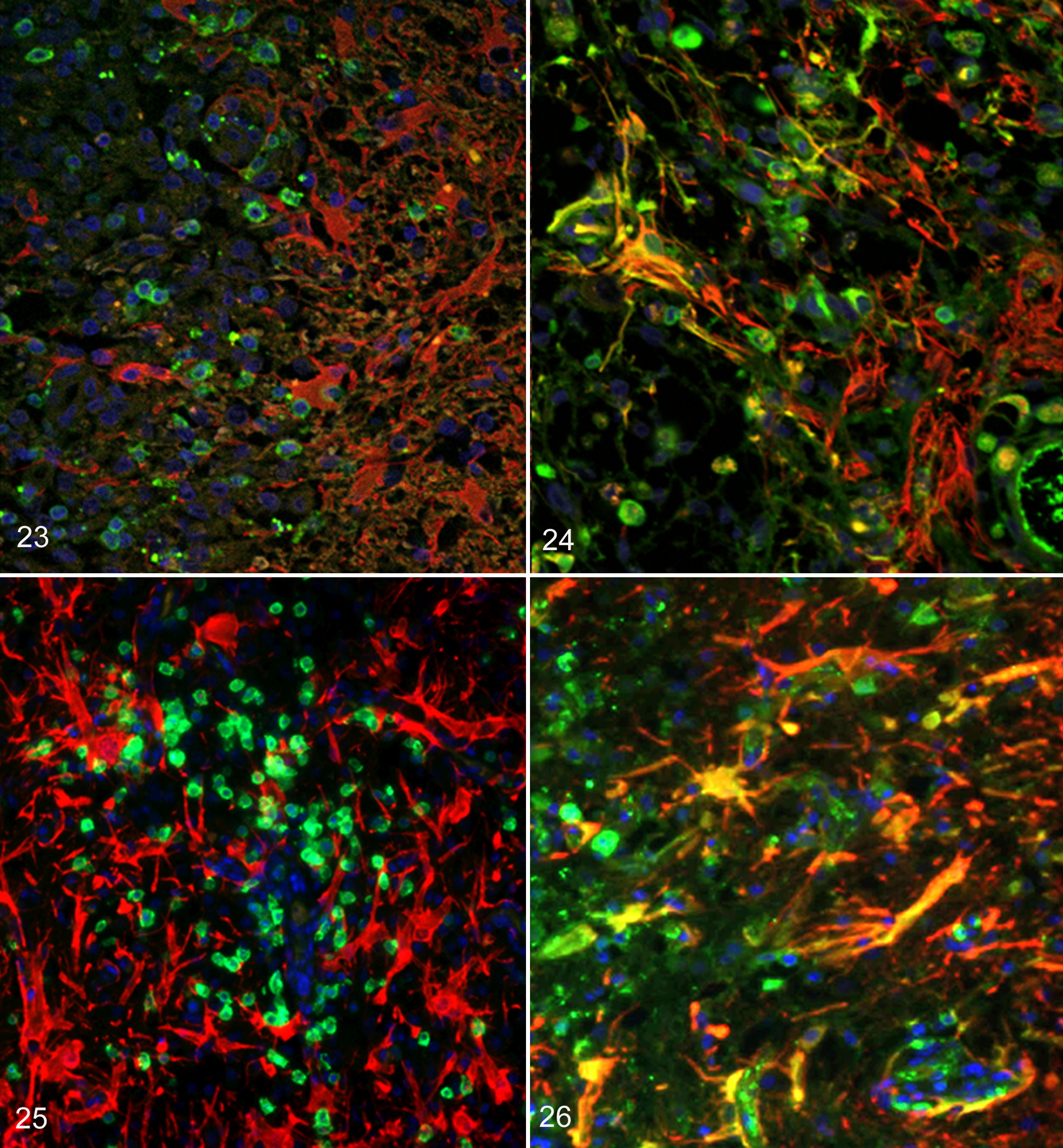
IgG and GFAP
The cytoplasm and/or processes of some astrocytes were labeled with both IgG and GFAP (yellow) in both NME (Fig. 24) and NLE (Fig. 26) but not in GME. These double-positive astrocytes were more prominent in NME than NLE and were frequently observed in the leptomeninges and the periphery of the lesions displaying necrosis and astrogliosis.
Discussion
In the present study, the brain lesions observed in Pug, Papillon, Chihuahua, and Maltese dogs with NME were mainly characterized by cerebral cortical lesions such as malacia, perivascular cuffs, and the neuroparenchymal infiltration of inflammatory cells. The lesions were located in the white matter as well as the cerebral cortex in a few Pug, Papillon, and Chihuahua dogs. In contrast, similar brain lesions were localized in the white matter of Yorkshire Terriers with NLE. Thus, the distribution of NME and NLE lesions seems to be breed specific. On the contrary, in dogs with GME, the characteristic granulomatous lesions were mostly located in the cerebellum, brainstem, and spinal cord, and additional nongranulomatous inflammatory lesions were found in the cerebral white matter and mesencephalon. Consequently, in NME and NLE, the histopathological lesions are malacic or inflammatory and are found in the prosencephalon and parts of the mesencephalon, whereas those found in GME are granulomatous or inflammatory and are usually located in the rhombencephalon. The distribution of the lesions in these canine brain inflammatory diseases was consistent with that described in previous reports.1, 2, 4, 8, 9, 12–16, 20, 24–27, 29, 32, 34, 41, 42, 48, 49, 51–54, 56
In a comparison of the inflammatory cell populations among NME, NLE, and GME, CD3-positive T cells were found to be the most prominent inflammatory cells in these diseases and were most often observed in the GME lesions, followed by those of NLE and NME, confirming the results of previous reports. 16,20,24,26,48,51 –53 CD163-positive macrophages were more predominant in GME than in NME or NLE, although the difference was not significant. It is plausible that the macrophages that infiltrate into the malacic lesions in the brains of dogs with NME or NLE act as scavenger cells that remove malacic or inflammatory products. 35 In contrast, the macrophages in GME lesions are thought to form granulomatous lesions as part of an immune response. 24,32,35,51,53 In this study, the proportion of IgG-positive plasma cells and CD20- and/or CD79acy-positive B cells was not significantly different among NME, NLE, and GME. B cells seemed to infiltrate into the neuroparenchyma less than plasma cells because CD20-positive B cells were observed more in the perivascular cuffs than in the neuroparenchyma. The proportion of inflammatory cells did not differ significantly among NME, NLE, and GME, although there were marked differences in the histopathology of the lesions and their locations. Taken together, these results suggest that these diseases share a common pathogenesis in dogs.
This retrospective study has several limitations such as variability in disease duration and treatment. The majority of cases had been diagnosed presumptively as NME, NLE, or GME through clinical signs, autoantibody test, computed tomography (CT), or magnetic resonance imaging (MRI). The dogs were treated with immunosuppressive drugs, such as corticosteroids (prednisone, prednisolone, and dexamethasone) and anticonvulsants such as phenobarbital and pentobarbital. They died or were euthanized due to poor response to the treatment, and a definitive diagnosis was made with NME, NLE, and GME through the histopathological examination. Corticosteroids such as dexamethasone and prednisone inhibit T cell– or macrophage-mediated immunity, production of cytokines, and partial B cell–mediated immunity. 6,10,11,22,33,40,50 This study was performed with the cases that progressed acutely or chronically despite continuous treatment and maintenance. Certainly, some degree of inflammatory reactions could have been affected by corticosteroids. However, inflammatory reactions of cases in this study were active, and characteristic lesions such as malacic or granulomatous changes were observed. This suggests that the nature of the inflammatory reactions was not significantly distorted by treatment.
Double-labeling immunofluorescence antibody (IFA) was performed to identify the targets of inflammatory cells. CD3-positive T cells were found in close proximity or attached to astrocytes, and the cytoplasm and astrocytic processes were positive for IgG in NME and NLE lesions, although the distribution of the lesions differed among the diseases. These IgG-positive astrocytes were more prominent in NME than in NLE. Because NME prominently affects the cerebral gray matter, which is composed of protoplasmic astrocytes, and NLE affects the white matter, which is composed of fibrous astrocytes, the difference in the lesion distribution between NME and NLE can be attributed to the targets of autoantibodies against protoplasmic astrocytes or fibrous astrocytes. 37,58 In addition, because astrogliosis was more significant in NME than in NLE and the proportion of autoantibodies in dogs with NLE was low compared with those described for NME in previous reports, 36,53 it could be considered that these differences are related to the etiologies of the various conditions. In fact, in some previous reports, 36,47,53,55 the involvement of an autoantibody to astrocytes in the etiology of NME was indicated. However, double-labeling IFA could not confirm whether these autoreactive inflammatory cells against GFAP are a primary cause or a secondary consequence of NME.
In some cases, malacic changes and inflammatory lesions were observed both in the gray matter and white matter in NME and NLE, although the distribution of lesions was different. The proportion of inflammatory cells was not significantly different. The presence of IgG in GFAP-positive astrocytes was observed in both NME and NLE. Thus, NME and NLE may be considered the same disease entity, necrotizing encephalitis, and divided into subtypes, the gray matter form and the white matter form, according to the distribution of lesions.
Although there have been many attempts to detect specific pathogens or antigens that induce NME, NLE, or GME, all have failed except for the reports detailing the involvement of GFAP.4, 8, 9, 20, 32, 34, 36, 39, 42, 45–49, 51, 53, 54, 55–57 Similarly, no specific pathogens or antigens have been detected in human multiple sclerosis (MS). 3,23,30,38 Multiple sclerosis is an autoimmune-mediated, inflammatory, and demyelinating inflammatory disorder affecting the CNS white matter, which is divided into several subtypes according to the lesions produced and the clinical course of the disease. It is disputed whether MS is mediated by major histocompatibility complex (MHC) class II–restricted CD4+ T cells, MHC class I–restricted CD8+ T cells, or antibody-complement complex. 5,21,23,28,30,31,38,43 However, other evidence suggests that genetic factors such as HLA-DR2 play a crucial role in susceptibility to the familial type of MS. 18,19,31,38 Chromosomes 17q22 and 6p21 (MHC) may also be associated with MS. 18,38,44 Taken together, multiple etiological factors, including both environmental and genetic factors, are likely to correlate with the onset of MS. In fact, it was reported that DLA-DR class II antigen may be associated with the cause of NME in Pug dogs 17 and that the mutation of mitochondrial DNA is related to NLE in Yorkshire Terriers. 2 Canine brain diseases might be caused by genetic and environmental factors as well as multiple immune-mediated mechanisms.
Footnotes
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The authors received no financial support for the research, authorship, and/or publication of this article.