
Abstract
The origin and transmission routes of atypical bovine spongiform encephalopathy (BSE) remain unclear. To assess whether the biological and biochemical characteristics of atypical L-type BSE detected in Japanese cattle (BSE/JP24) are conserved during serial passages within a single host, 3 calves were inoculated intracerebrally with a brain homogenate prepared from first-passaged BSE/JP24-affected cattle. Detailed immunohistochemical and neuropathologic analysis of the brains of second-passaged animals, which had developed the disease and survived for an average of 16 months after inoculation, revealed distribution of spongiform changes and disease-associated prion protein (PrPSc) throughout the brain. Although immunolabeled PrPSc obtained from brain tissue was characterized by the presence of PrP plaques and diffuse synaptic granular accumulations, no stellate-type deposits were detected. Western blot analysis suggested no obvious differences in PrPSc molecular mass or glycoform pattern in the brains of first- and second-passaged cattle. These findings suggest failures to identify differences in mean incubation period and biochemical and neuropathologic properties of the BSE/JP24 prion between the first and second passages in cattle.
Bovine spongiform encephalopathy (BSE) is a fatal neurodegenerative disorder manifested as one of a group of transmissible spongiform encephalopathies, including scrapie in sheep and goats, chronic wasting disease in deer, and Creutzfeldt-Jakob disease in humans. The infectious agent responsible for these diseases is an abnormal isoform prion protein (PrPSc), currently thought to be a posttranslationally modified form of a host-encoded membrane glycoprotein (PrPC). The uniform nature of the pathology and biochemical features of the proteinase K–resistant PrPSc (PrPres) profile obtained for cattle infected with BSE and mice in BSE transmission studies suggests that BSE is caused by a single prion strain.
Recently, variants of BSE (denoted atypical BSE) have been detected in cattle in Japan, Europe, and North America. 2 Currently, atypical BSE cases are classified into at least 2 groups, namely the L-type and the H-type, according to the molecular weight of the PrPres in the case compared to that of classical (C-type) BSE cases. Histopathologic as well as immunohistochemical analyses have confirmed that atypical H- and L-type forms of BSE are transmissible to mice and clearly differ from C-type BSE in terms of incubation period, PrPres profile, and pathology. 1 The L-type BSE recognized in Italy, which has been further characterized by the presence of PrPSc-positive amyloid plaques in the brain, has been termed bovine amyloidotic spongiform encephalopathy (BASE). 5
Western blot analysis of one 14-year-old Japanese black beef cow (BSE/JP24) 7 suggested a glycoform ratio similar to that of BASE-infected cattle, while PrPSc immunohistochemistry analysis suggested the presence of PrPSc-positive amyloid plaques in the brain similar to those found in BASE-infected cattle. 5 Additional studies have demonstrated that the BSE/JP24 isolate can be transmitted to cattle 6 and that BSE/JP 24 and BASE have nearly the same incubation period during the first passage in Holstein cattle. 6,8
Research has also suggested that during subsequent passages, the incubation period of the disease may be shorter or more stable than that of the first passage and that its characterization may vary. The primary objective of this study was to further investigate the characteristics of the second passage of the disease by clinicopathologic analysis of BSE/JP24-infected cattle. The secondary objective was to fill a significant research gap by further identifying and describing the topographic distribution and patterns of immunolabeled PrPSc, thereby redressing the paucity of immunohistochemical data that have been collected regarding atypical BSE cases.
All the experiments involving animals were performed in the biosafety level 3 areas with the approval of the Animal Ethical Committee and the Animal Care and Use Committee of the National Institute of Animal Health. Three Holstein female calves aged 2 to 3 months old were inoculated through the intracerebral route with 1 ml of 10% brain homogenate prepared from the brainstem of a first-passaged BSE/JP24-affected cow (case No. 1A; code 8515) used in a previous experiment. 6 The brain material was confirmed positive by Western blot and immunohistochemical analysis. Two uninoculated calves served as controls and were sacrificed 24 months after inoculation.
Between 12 and 13 months after inoculation, the animals began to exhibit signs of fear or anxiety, which are initial clinical signs of BSE. The animals were observed to exhibit ataxia of the hind limbs before astasia and were subsequently killed at 421 (case No. 2A), 504 (case No. 2B), and 559 (case No. 2C) days after inoculation. The results of unpaired t testing (InStat3, GraphPad Software, La Jolla, CA) failed to identify a significant difference in mean incubation period between the first- and second-passaged cattle (486 ± 11 days 6 and 495 ± 69 days, respectively).
After the brain was cut longitudinally at necropsy, the left half, including the brainstem and cerebellum, was fixed in 10% neutral buffered formalin containing 10% methanol, while the right half was frozen at −80°C for Western blot analysis. Formalin-fixed coronal slices of the brain and tissue samples from the entire body were trimmed into 3- to 4-mm thick sections, immersed in 98% formic acid for 60 minutes to reduce infectivity, rinsed, dehydrated, embedded in paraffin wax, sectioned at 4 μm, and stained with hematoxylin and eosin. A vacuolar lesion profile of the hematoxylin and eosin–stained sections was obtained using the method described by Simmons et al. 10 Selected sections were stained with phenol Congo red and examined under a polarizing microscope, and the presence of amyloid was confirmed by observation of its characteristic dichroism.
For the demonstration of immunolabeled PrPSc, dewaxed sections were placed on a silanated slide, treated with 3% hydrogen peroxide at room temperature for 10 minutes, incubated with 10 μg/ml of proteinase K (0.03 U/μg; Nakarai, Kyoto, Japan) in phosphate-buffered saline containing 0.1% Triton-X at room temperature for 10 minutes, and immersed in 150mM sodium hydroxide at 60°C for 10 minutes. 6,9 The sections were then incubated with monoclonal antibody F99/97.6.1 (1 μg/ml; VMRD Inc., Pullman, WA), which reacts with a conserved epitope located at amino acid residues 228–233(QYQRES) of the bovine prion protein when using a horseradish peroxidase–labeled polymer detection system (Nichirei Histofine Simple Stain MAX-PO (M), Nichirei, Tokyo, Japan). The sections were then visualized using 3′-3 diaminobenzidine tetrachloride as the chromogen in accordance with the manufacturer’s instructions and slightly counterstained with hematoxylin. The intensity and extent of PrPSc accumulation were subjectively scored from 0 to 4 (0, negative; 1, apparent at high magnification; 2, apparent at moderate magnification; 3, apparent at low magnification and moderate amounts of accumulation; 4, large amounts of accumulation). Negative controls were incubated with nonimmune mouse or rabbit IgG (1:30 dilution; Dako) and phosphate buffered saline instead of the primary antibody. Schematic representation of immunolabeled PrPSc distribution in the brain was drawn with Adobe Photoshop. Proteins were extracted from the frozen samples and subjected to Western blot analysis of PrPres according to a previously described method. 6 The blotted membrane was then incubated with anti-PrP-specific monoclonal antibody T2 and the signals detected using a chemiluminescent substrate (SuperSignal; Thermo Fisher Scientific Inc., Rockford, IL). The signal intensity and glycoform ratio of PrPSc were calculated with Fluorochem software (Alpha Innotech, San Leandro, CA). Brains of 2 sham-inoculated Holstein cattle served as controls for immunohistochemical and Western blot analyses.
The results of the analysis suggested that the neuropil but not the neuronal vacuolation was consistent in most nuclei of the thalamus and brainstem structures, including the midbrain, pons, and medulla oblongata, but not in the dorsal motor nucleus of the vagus nerve. The vacuolar lesion score in the second-passaged animals was similar to that in first-passaged cattle, as previously described (data not shown). 6 The variability in PrPSc immunolabeling intensity and distribution pattern was found to be nearly identical. Seven varieties of PrPSc immunostaining patterns—including intraneuronal, perineuronal, intraglial, linear, fine particulate, coarse granular, and plaques—were identified throughout the brain of first- and second-passaged animals. As can be observed in Figure 1, which shows the topographic distribution of the PrPSc identified, significant quantities of PrPSc were detected in the brainstem and thalamus. The most conspicuous pattern of PrPSc was that of fine particulate and coarse granular deposits in the neuropil of the thalamus and midbrain, particularly in the periaqueductal grey matter, and to some extent in the cerebral cortices and cerebellar nuclei (Fig. 2). The major feature of the plaques was a spherical structure with a unicentric core up to 20 μm in diameter. The plaques were sparsely detected in the thalamus, basal ganglia, midbrain, pons, deeper layers of the cerebral cortices, and subcortical white matter and were not detected to any extent in the olfactory bulb, medulla oblongata, cerebellum, or spinal cord. These plaques exhibited a variable morphology; although most plaques that had a uniformly dense core were noncongophilic and less than 15 μm in diameter, some that had a lucent core were congophilic, larger than 15 μm in diameter, irregularly outlined, and showed distinctly intense surrounding immunolabeling (Fig. 3). The stellate-type of PrPSc deposition usually detected in the cerebral and cerebellar cortices of C-type BSE-affected cattle was undetectable in any sections of any of the animals, while less intraneuronal deposition of PrPSc was found in the cerebral cortex compared to the thalamus and brainstem (Fig. 2). PrPSc immunolabeling was detected in extracerebral tissues, including the trigeminal and dorsal root ganglia (Fig. 4), neurohypophysis, and retina (Fig. 5). PrPSc accumulation was prominent in the ganglion cell layer as well as the inner and outer plexiform layers of the retina (Fig. 5). No histopathologic changes were present in the peripheral tissues, including the skeletal muscles.
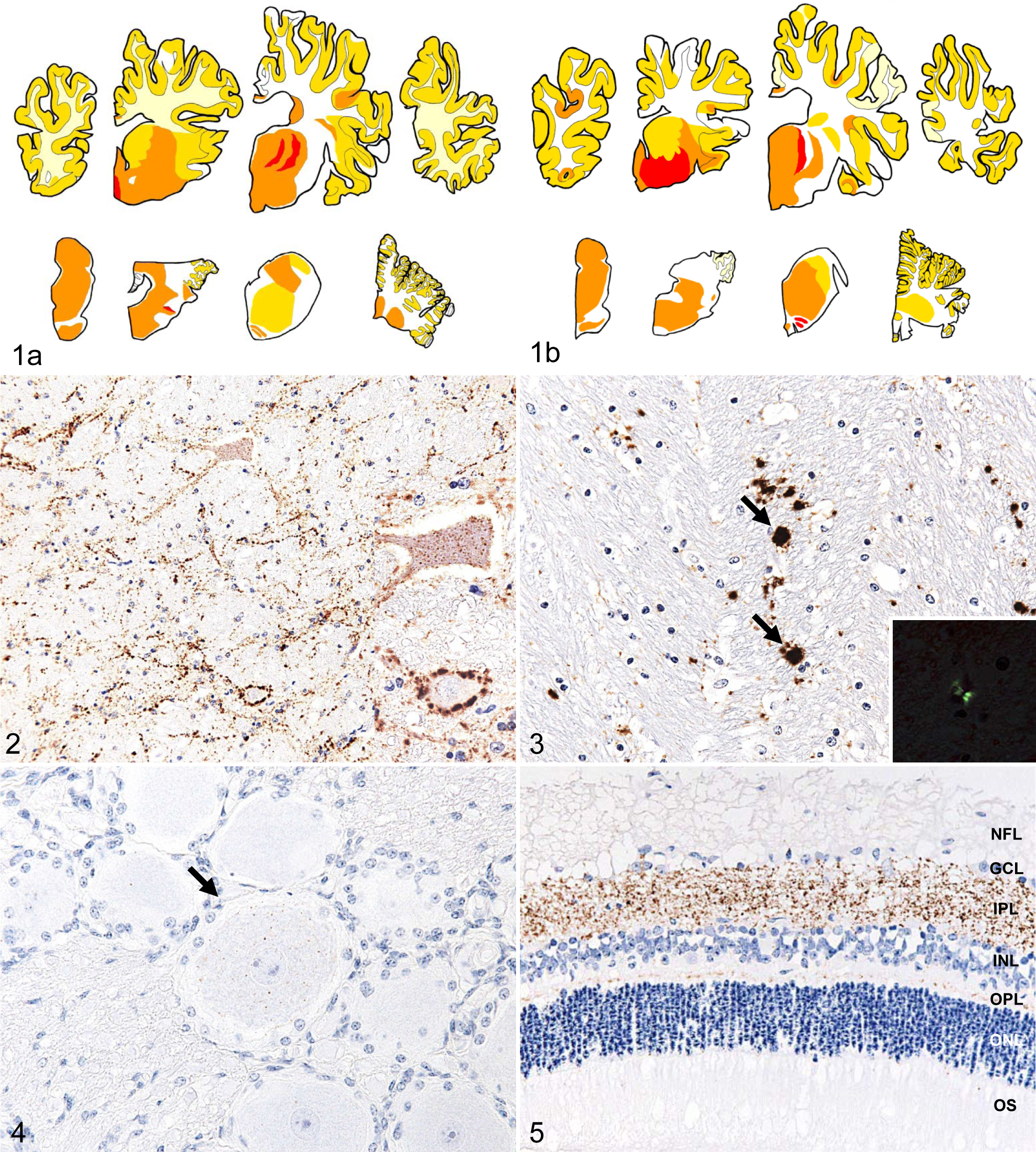
Immunolabeled PrPSc distribution in the central nervous system of (a) first- and (b) second-passaged cattle (respectively, case Nos. 1A and 2C). Immunolabeling intensity of PrPSc deposition was scored as none (white), scanty (pale yellow), mild (dark yellow), moderate (orange), and heavy (red). Eight brain areas can be observed, at the level of the frontal lobe, striatum, thalamus, occipital lobe, midbrain, pons, medulla oblongata at the obex, and the cerebellum, from the upper left to the lower right.
Western blot analysis of PrPres suggested that the brains of first- and second-passaged cattle had a similar molecular mass and exhibited similar glycoform patterns (Figs. 6, 7). The signal intensities in diglycosylated, monoglycosylated, and unglycosylated PrPres fragments were found to be similar in both the first- and second-passaged cattle. In addition, the unglycosylated PrPres fragment detected in both groups showed a lower molecular weight than that in cattle affected with a C-type BSE agent, suggesting that quantitative studies of PrPres molecular mass and glycoforms proportions in Western blots will be more useful to demonstrate the difference between the classical and different passages of atypical BSEs. The mean incubation period of the first- and second-passaged Holstein cattle challenged with the BSE/JP24 prion was approximately 16 months, the same as that of BASE-inoculated Holstein cattle. 8 In contrast, the mean incubation period of C-type BSE-challenged cattle ranges from 17 to 22 months after intracerebral inoculation. 8 In studies of bovine PrP-overexpressing transgenic mice, the incubation period was also shorter in L-type BSE-inoculated mice than in C-type. 3,4,9 These results suggest that in cattle, L-type BSE agents have a shorter incubation period compared to that of C-type BSE agents and that the biochemical properties of PrPSc from an original BSE/JP24 isolate have no evident differences in BSE/JP24-infected cattle. The incubation period and molecular profiles of PrPSc suggest that the isolate of BSE/JP24 is clearly distinct from that of C-type BSE while closely resembling that of BASE isolate. 5,6,8
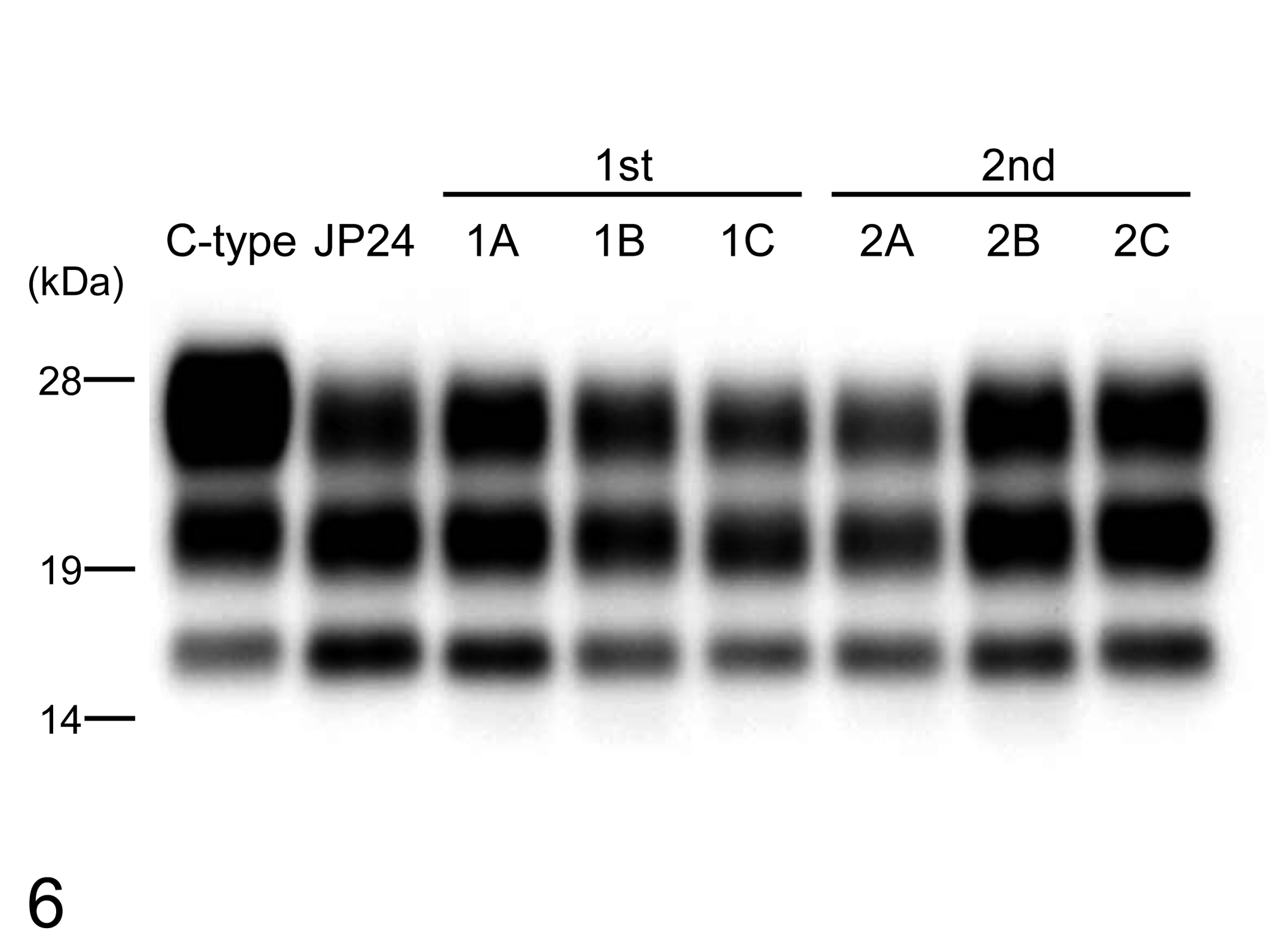
Western blot analysis of proteinase K–digested prion protein (PrPres) from the medulla oblongata at the obex level of C-type bovine spongiform encephalopathy (BSE) and BSE/JP24 first- and second-passaged cattle with monoclonal antibody T2. All samples were digested with 50 μg/ml of proteinase K at 37°C for 1 hour. Molecular markers are shown on the left.
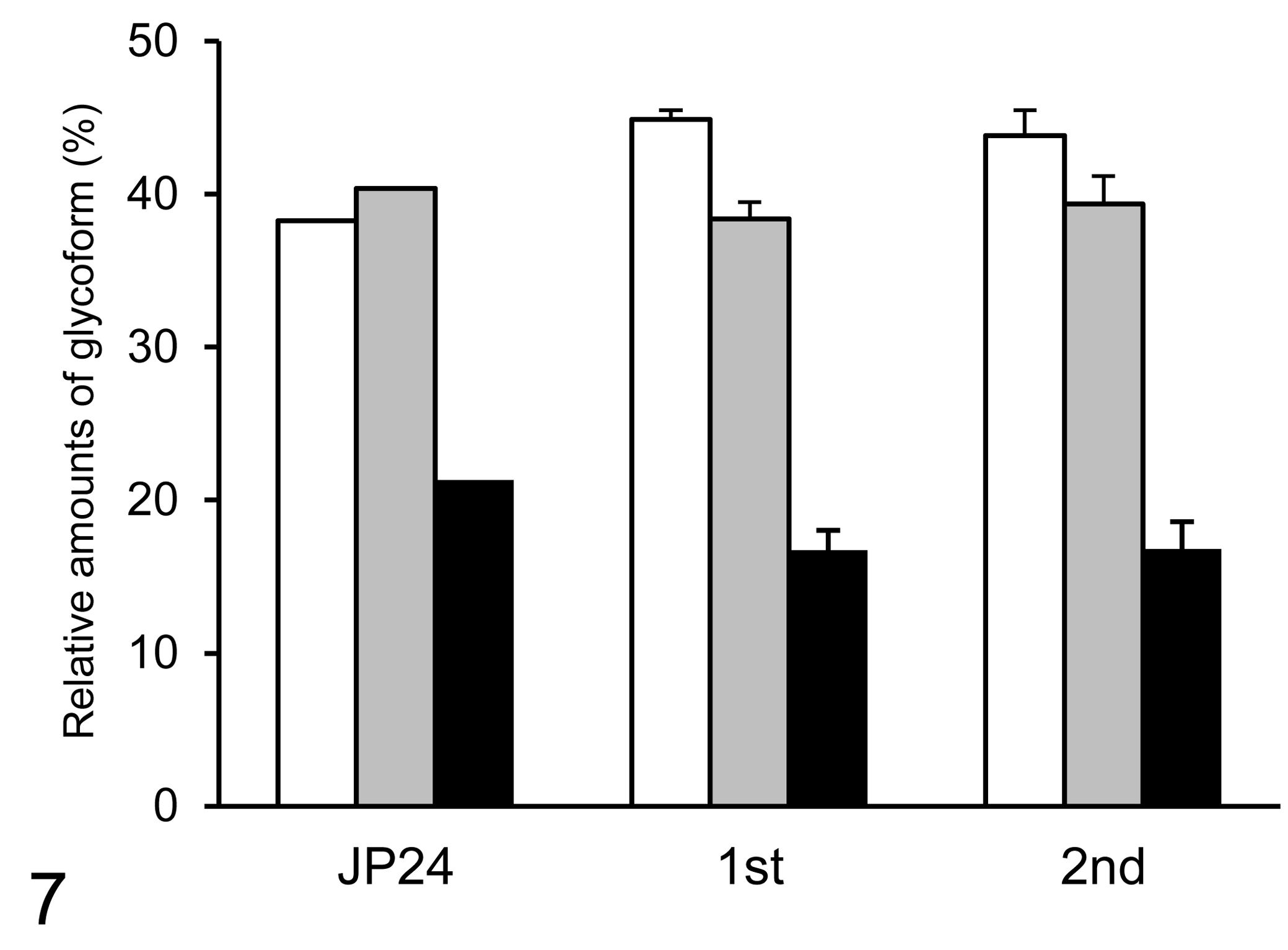
The relative amount of the diglycosylated, monoglycosylated, and unglycosylated form of PrPres in the BSE/JP24, first-passaged, and second-passaged prion-affected individual with monoclonal antibody T2. The results are shown as mean ± standard deviation in triplicate experiments. Bar diagram indicates diglycoslated form (black column), monoglycosylated form (gray column), and unglycosylated form (white column).
Immunohistochemical analysis revealed that the patterns of PrPSc deposition in BSE/JP24 prion-affected cattle were characterized by the presence of amyloid PrP plaques and the absence of stellate-type PrPSc deposits. However, except for the detection of higher quantities of PrPSc in the cerebral cortices of BSE/JP24-affected cattle, no striking differences were identified between the topographic distribution of PrPSc in the C-type BSE and BSE/JP24 prion-affected cattle. 6 The incubation periods and neuropathologic and immunohistochemical characteristics of the BSE/JP24-inoculated Holstein cattle closely resemble those of BASE-affected cattle, 8 indicating that both L-type BSE prions have similar biological properties.
Although atrophy of both type I and II muscle fibers had been detected in BASE cases in previous experiments, 8 evident changes were not detected in the skeletal muscle of the cases examined in this study. This discrepancy may have been due to the fiber-type grouping used in the previous studies, which had resulted in neurogenic atrophy of the innervated muscles, indicating that the muscular lesions in the BASE cases might not be associated with a neurogenic disorder. The discrepancy might also be due to the use of different technical protocols in tissue processing—specifically, freezing of sections versus formalin fixing and paraffin embedding of sections.
Although L-type BSE cases have been detected throughout Europe, it remains unclear whether the BSE/JP24 prion is identical to other L-type BSE prions. Although the results of this study may accord with those obtained in studies of cattle intracerebrally inoculated with BASE, further research is required to determine if they truly do so. In conclusion, the results of the detailed immunohistochemical and neuropathologic analysis conducted in this study suggest the failure to identify differences between the first- and second-passaged cattle regarding the variables of survival period, molecular properties of PrPres, and other immunohistochemical or neuropathologic features; this suggests that the BSE/JP24 isolate is a biologically and biochemically stable prion in cattle. The findings of the present study will aid future transmission experiments on different L-type BSE isolates.
Footnotes
Acknowledgements
Expert technical assistance was provided by Junko Endo, Mutsumi Sakurai, Noriko Amagai, Tomoko Murata, Naoko Tabeta, and the animal caretaker.
Declaration of Conflicting Interests
The authors declare that they have no conflicts of interests with respect to their authorship or the publication of this article.
Funding
This work was supported by grants from the other Prion Disease Control Project of the Ministry of Agriculture, Forestry, and Fisheries of Japan and the BSE Research Project of the Ministry of Health, Labor, and Welfare of Japan.