
Abstract
Avian metapneumovirus (aMPV) is a respiratory virus that infects a range of avian hosts, including chickens and turkeys. Migratory and local wild birds are implicated in aMPV spread among farms, countries, and seasonal outbreaks of the disease. A subtype B aMPV isolate from commercial chicken flocks suffering from respiratory disease was experimentally inoculated oculonasally into 7-week old chickens, young pigeons, and sparrows. Chickens showed minimal tracheal rales, whereas pigeons and sparrows were asymptomatic. Shedding of aMPV was detected by reverse transcription polymerase chain reaction on homogenates from nasal turbinates. At 5 days postinfection, 5 of 5 chickens, 2 of 5 pigeons, and 1 of 5 sparrows were positive; at 10 or 15 days, none were positive. At 2 and 5 days, aMPV antigens were localized at the ciliated boarder of respiratory epithelium in nasal cavity and trachea of chickens, as well as to the conjunctival epithelium. Pigeons had detectable viral antigens in only the trachea at 2 and 5 days; sparrow tissues did not show any positive staining. At the end of the experiment, at 21 days postinfection, 14 of 15 inoculated chickens seroconverted against aMPV, but none of the inoculated pigeons or sparrows did. The authors believe that pigeons and sparrows have the ability to transmit the virus between chicken farms, although they do not consider pigeons and sparrows as natural hosts for aMPV, given that they failed to seroconvert. In conclusion, pigeons and sparrows are partially susceptible to aMPV infection, probably acting more as mechanical vectors because infection is only temporary and short-lived.
Avian metapneumovirus (aMPV) is the etiologic agent of turkey rhinotracheitis and swollen head syndrome in chickens, and it refers to upper respiratory tract infection of turkeys and some other avian species. This virus was previously referred to as avian pneumovirus. 10
Migratory wild birds are implicated in transmitting aMPV over long distances,3,5,15 and local wild birds are implicated in transmitting aMPV among commercial poultry flocks in endemic areas because poultry houses have big attractions for local wild birds in terms of available feed and shelter. Sparrows and pigeons are the most common wild birds around and in poultry farms in many countries. Researchers have detected aMPV RNA in sparrows from areas where aMPV infected commercial poultry farms.3,15 However, their studies did not investigate whether sparrows were a natural host for aMPV and whether they were able to sustain replication and shedding. Other studies demonstrated that aMPV of turkey origin was not infectious to pigeons, with no reisolation or seroconversion to the virus after experimental infection. 9
Chickens experimentally infected with turkey-origin aMPV had mononuclear inflammatory cell infiltrates in nasal turbinates and airways in lungs. Based on immunohistochemical technique, viral antigens were present up to 5 days postinfection (DPI) and were associated with cilia of the epithelial cells of nasal turbinates and in tracheas when inoculated intratracheally. No other tissues contained detectable antigens (lung, conjunctiva, magnum, isthmus, infundibulum, uterus, liver, kidney, thymus, spleen, and central nervous system). 14 In a later study, a similar turkey-origin isolate caused severe respiratory disease in chickens, as evident by clinical signs and histopathologic lesions in the respiratory organs. Viral antigen distribution was similar and restricted to the same postinfection period and same organs as the previous study indicated. 4 Turkeys infected by dropping inoculums in the nasal cavity and the conjunctiva had aMPV antigens in sinus and turbinates up to 8 DPI and, occasionally, in some trachea sections at only 4 and 6 DPI. 12
We have documented the presence of aMPV subtype B in Jordanian chicken flocks. 8 In this study, we investigate the susceptibility of pigeons and sparrows to this subtype B aMPV and their possible role in transmission among chicken farms.
Materials and Methods
Virus Inoculum
A local isolate of aMPV subtype B 8 was used to generate infectious nasal turbinate homogenates from infected specific-pathogen-free chicks. The inoculum was bacteria-free and tested by reverse transcription polymerase chain reaction (RT-PCR) to be positive for aMPV subtype B and to be negative for Newcastle disease virus, avian influenza virus, and infectious bronchitis virus.
Birds and Housing
One-day-old Hubbard breed broiler chicks were obtained from a local commercial breeder farm known to be Mycoplasma gallisepticum-free and reared until 7 weeks of age. Young-adult pigeons (Columba livia) were purchased from a local bird market. Adult sparrows (Passer domesticus) were caught from local areas. All birds were raised in environment-controlled rooms in the Animal House Facility at Jordan University of Science and Technology.
Experimental Design
Two days before challenge, around 15 blood samples from each species were collected, and birds were tested to be free from aMPV infection by competitive ELISA. Four chickens, sparrows, and pigeons were euthanized and their nasal turbinate homogenates tested negative for aMPV by RT-PCR. Their tissues were used as negative control for immunohistochemical staining. Chickens, pigeons, and sparrows were separated into challenge (n = 40) and control (n = 17) groups. Birds in the challenge group were infected by dropping 50 μl of the aMPV inoculum in each eye and nostril, for a total of 200 μl for each bird. The 2 groups were housed in separate rooms. Postinfection, birds were observed daily for clinical signs. Five birds from each species from the infected group were euthanized on 5, 10, and 15 DPI and turbinates collected for aMPV RT-PCR. On 3 and 5 DPI, 2 and 3 birds, respectively, from each species of the infected group were randomly selected and euthanized to collect complete set of tissues for immunohistochemical staining. At 21 DPI, 15 blood samples from each species in each group were collected for serologic testing.
Serology
Blood samples were collected from each species and tested by competitive ELISA using the turkey rhinotracheitis–swollen head syndrome avian pneumovirus ELISA kit (Svanova Biotech, Lyon Cedex, France) according to the manufacturer’s instructions and similar to our previous study. 8
Antiserum (Primary Antibody) Production for Immunohistochemistry
Two young adult female rabbits about 2.7 to 3.0 kg (Animal House Facility, Yarmouk University, Irbid, Jordan) were injected subcutaneously with 4 ml of commercial avian pneumovirus vaccine Nobilis RT Inac (Intervet Int BV, Boxmer, Holland) into 4 sites. They received 2 additional 2-ml injections (1 injection every 4 weeks). Two weeks after the third injection, rabbits' sera were tested by competitive ELISA for aMPV antibodies. One rabbit’s serum had a postinfection value of 58.03 (strong positive).
Immunohistochemistry
After necropsy, tissues were immediately fixed in 4% phosphate-buffered formaldehyde for 24 hours, then routinely processed. The above primary antibody (1:2000 dilution) with Dako EnVision + Dual Link System-HRP system (DAKO Inc, Carpinteria, CA) was used according to the manufacturer’s instructions and our previous study. 7 Negative controls were performed using negative control tissues and negative control serum. Tissues with multifocal staining in less than 30% of susceptible cells were defined as having mild staining (+); in 30 to 80%, moderate staining (++); and more than 80%, intense staining (+++).
Avian Metapneumovirus Shedding
Virus shedding was determined by testing nasal turbinate homogenates for aMPV by RT-PCR. RNA was extracted from nasal turbinate homogenate supernatant with an RNA purification kit (Total RNA Isolation System, Promega Corp) according to the manufacturer’s procedure. One-step RT-PCR was performed with the Access RT-PCR System (Promega Corp) according to the manufacturer’s procedure and a previously described primers and procedure for subtype B detection. 2 An aMPV subtype B live vaccine (Hipraviar SHS, HIPRA, Girona, Spain) was used as a positive control for RNA extraction and RT-PCR.
Results
Clinical and Pathologic Observations
Minimal tracheal rales were the only respiratory sign observed in inoculated chickens 2 to 5 DPI. Yet, sparrows and pigeons from the inoculated group both appeared normal. No clinical signs were observed in the noninoculated group. On necropsy of birds for collection of tissue for immunohistochemical and virus shedding, no significant gross lesions were observed in the inoculated chickens, sparrows, or pigeons.
Serology
At 21 DPI, 14 of 15 chickens seroconverted positively, and 1 chicken had a doubtful result (Table 1 ), but all inoculated pigeons and sparrows and the uninoculated control group remained negative.
ELISA Results and Percent Inhibition for Avian Metapneumovirus in 15 Chicken Sera at 21 Days Postinfection
a[(optical density of the negative control − optical density of the sample) × 100] / optical density of the negative control
b < 30%, negative (–); 30–40%, doubtful (±); > 40%, positive (+).
Immunohistochemistry
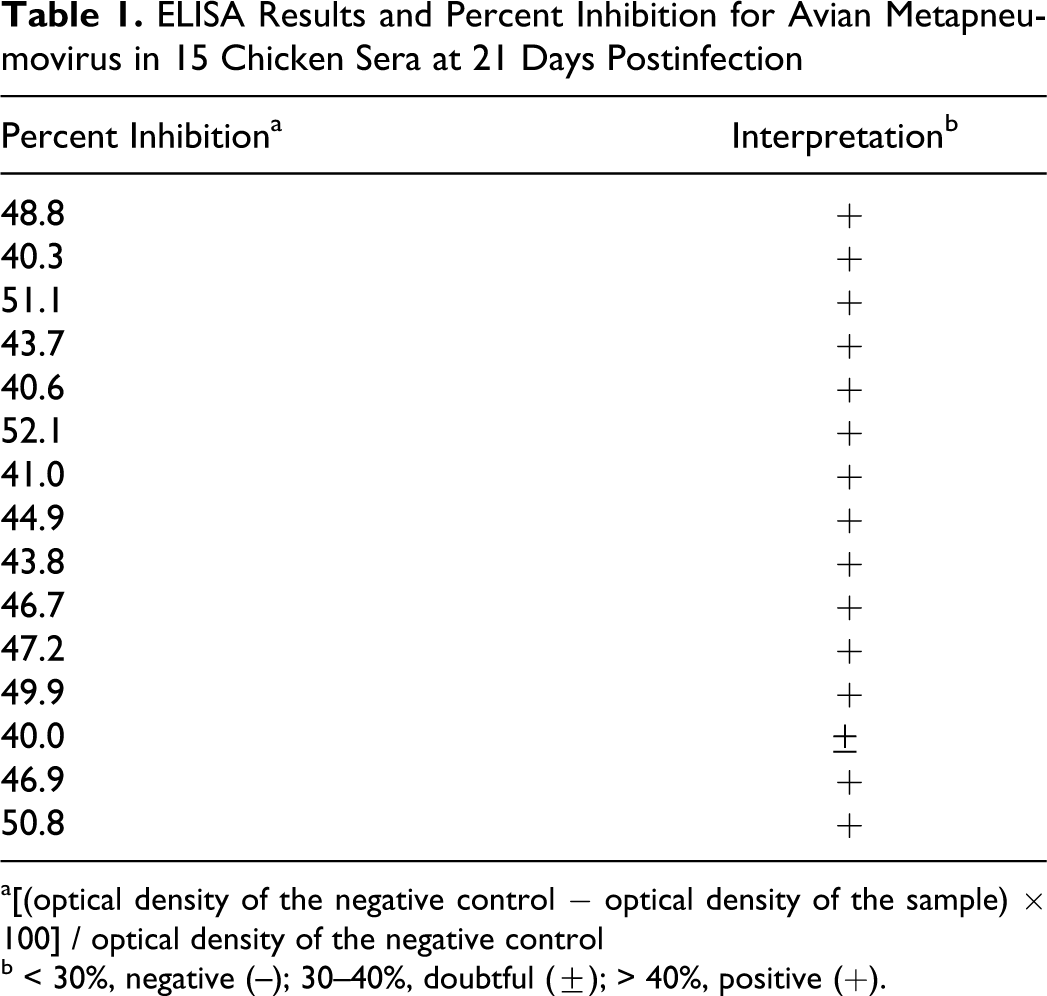
Avian metapneumovirus antigens were detected by localization of granular brown pigment of different intensities on the apical portion of the ciliated epithelium of the respiratory organs and were closely associated with the cilia. Chicken tissues had the most intense staining in the apical border of conjunctival (Fig.1 ) and nasal epithelium (Fig. 2) at 3 and 5 DPI. The trachea had moderate staining in chickens (Fig. 3) and mild staining in pigeons at 3 and 5 DPI (Fig. 4). Sparrow tissues did not show any positive staining. Other tissues examined did not have any detectable antigen (airsac, brain, heart, liver, spleen, kidney, duodenum, pancreas, proventriculus, gizzard, thymus, cecal tonsil, bursa of Fabricius, testes and ovary).
Virus Shedding
Table 2 summarizes aMPV shedding in nasal turbinates as detected by RT-PCR. There was shedding at only 5 DPI. Viral RNA was detected in nasal turbinate homogenate in 5 of 5 chickens, 2 of 5 pigeons, and 1 of 5 sparrows (Fig. 5 ). No virus RNA was detected from the 3 species at 10 or 15 DPI.
Avian Metapneumovirus Shedding in Nasal Turbinate Homogenate Detected by Reverse Transcription Polymerase Chain Reaction a
a number of birds in which avian metapneumovirus was detected / total number of birds examined.
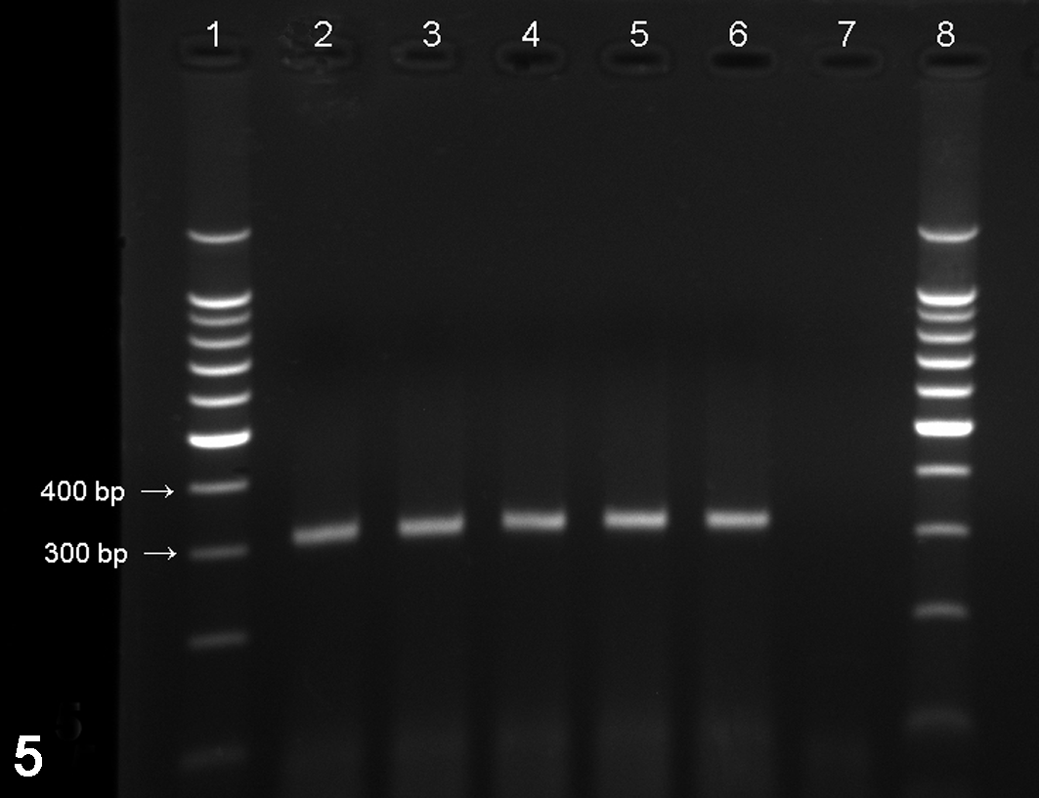
Reverse transcription polymerase chain reaction for aMPV subtype B. Positive reaction will show 312–base pair amplicon on agarose gel. Lanes 1 and 8, 100–base pair marker; lane 2, positive control (aMPV vaccine); lanes 3–6, examples of positive samples; lane 7, negative control.
Discussion
The clinical signs observed in these 7-week-old broiler chickens attributed to aMPV infection in this experiment are minimal compared to the ones seen in field cases, where they are more severe and where some chickens exhibit swelling of the head. Research has shown that experimental aMPV infection does not always produce appreciable clinical signs. For example, aMPV did not cause any respiratory signs in 27-week-old broiler breeder chickens, but it did cause mild respiratory signs in 3-week-old layer chickens. 14 When severe respiratory signs are observed in association with aMPV in chickens in the field, the typical belief is that they are a result of coinfection with other respiratory or secondary pathogens. 10 Producing clinical signs similar to those seen in the field cases of aMPV was possible by coinfecting turkeys with aMPV and Newcastle disease virus. 16 Another factor potentially contributing to the mild clinical signs is the age of the chickens at the time of infection; increasing the dose of challenging virus with increasing age is necessary to reproduce clinical signs. 17 The chickens in this experiment were 7 weeks old at the time of infection.
We detected seroconversion for aMPV infection in chickens by competitive ELISA. At 21 DPI, almost 100% of the chickens seroconverted (Table 1). However, pigeons did not seroconvert, which agrees with previous studies. 9 To the best of our knowledge, no previous studies have reported serologic testing of sparrows, and the current study is the first to report that aMPV-inoculated sparrows did not seroconvert.
In our experiment, aMPV shedding was detected by RT-PCR testing on nasal turbinate tissue homogenates, because turbinates are the most reliable site for detecting aMPV infection in turkeys. 11 In chickens, aMPV RNA was detected in all tested 5 chickens at 5 DPI; the virus was not detected at 10 or 15 DPI, which agrees with previous studies regarding aMPV shedding in chickens1,4,14 and which correlates with its shedding in turkeys.6,13 In 2 of 5 pigeons, we detected aMPV in turbinate homogenates at 5 DPI. Previous studies did not detect aMPV in inoculated pigeons with a turkey strain. 9 This could be due to differences in either the virus (ie, strain and origin) or the pigeon (ie, breed and susceptibility to aMPV). Also, it is possible that we detected RNA from only inactive virus still present in the inoculation site, whereas the previous study detected viable virus by isolation. Sparrows can harbor aMPV: Its RNA in this study was detected in nasal turbinate tissues after infection, which agrees with previous investigations of sparrows' susceptibility to infection from infected turkey farms with subtype C .3,15 Failure of detecting aMPV after 5 DPI in any of the 3 species emphasizes the transient short-lived infection of the virus.1,4
In chickens, we detected only aMPV antigens by immunohistochemistry in some respiratory tissues. When tissues were positive, they were positive at 3 and 5 DPI, and the stain was detected in the apical portion of the ciliated respiratory epithelium and closely associated with the cilia, in addition to the conjunctival epithelium. Immunohistochemical results in chicken tissues of this study are consistent with those of previous studies in regard to tissue distribution,4,11,12,14 intensity of stain,11,12,14 and time of detection4,14 in either chickens or turkeys. In a previous study infecting chickens with subtype B aMPV, the intensity of stain was described to be extensive in the respiratory epithelium, unlike what we describe in this study. 4 However, in that study, the authors describe other dark-staining areas in the lamina propria of the mucosa, and they considered it nonspecific because this dark staining was also present in tissues from negative chickens. In contrast with previous studies, chicken conjunctiva had strong positive staining (Fig. 1). However, in previous studies infecting chickens 14 or turkeys 12 with a turkey-origin subtype A aMPV, the conjunctiva tested negative. This difference in tissue distribution might be due to differences in aMPV subtype and origin. The isolate in this study was from chickens, and it was classified as subtype B; the isolate that they used was turkey-origin subtype A. A more recent study showed that subtype B has more pathogenicity and wider tissue distribution in chickens, including the lacrimal gland, when compared to subtype A. 1 Unfortunately, the conjunctival tissue was not examined in that study to compare it to our results. In addition, it is possible that the sensitivity of the immunohistochemical kit used in this study is superior compared to kits used in previous studies.
Pigeons had only mild immunohistochemical staining in their tracheas but not in the conjunctiva or the nasal turbinates, and none of sparrow tissues tested positive. However, we detected the virus RNA in the turbinate homogenate from 2 of 5 pigeons and 1 of 5 sparrows at 5 DPI by RT-PCR, so the sensitivity of immunohistochemical technique was probably not high enough when compared to that of RT-PCR to detect the small amounts of antigen in the other tissues of pigeons or sparrows.
Similar to chickens, Pigeons and sparrows did not seroconvert after aMPV infection. However, a few of the inoculated birds shed the virus by 5 DPI, compared to all the tested chickens at that time. Neither the chickens nor the pigeons and sparrows shed virus by 10 DPI. It appears that the virus does not replicate to high titers in pigeon or sparrow tissues and that the small quantities of virus present or produced are not enough to stimulate a detectable immune response in pigeons or sparrows. We believe that pigeons and sparrows have the ability to transmit the virus between chicken farms. However, we do not consider pigeons and sparrows as natural hosts for aMPV, because they failed to seroconvert. In conclusion, pigeons and sparrows are partially susceptible to aMPV infection, and they probably act more as mechanical vectors because infection is only temporary and short-lived.
Footnotes
Acknowledgments
We would like to thank the Deanship of Research at Jordan University of Science and Technology and the Scientific Research Fund of the Ministry of Higher Education and Scientific Research for supporting this work. We also thank Dr Kamel Mahmoud for his comments and help with the statistical analysis.
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
The authors declared that they received no financial support for their research and/or authorship of this article.