
Abstract
An H3N8 influenza virus closely related to equine influenza virus was identified in racing greyhound dogs with respiratory disease in 2004 and subsequently identified in shelter and pet dogs. Pathologic findings in dogs spontaneously infected with canine influenza virus were compared with lesions induced in beagle and mongrel dogs following experimental inoculation with influenza A/canine/Florida/43/2004. BALB/c mice were inoculated with canine influenza virus to assess their suitability as an experimental model for viral pathogenesis studies. All dogs inoculated with virus developed necrotizing and hyperplastic tracheitis and bronchitis with involvement of submucosal glands as well as mild bronchiolitis and pneumonia. Viral antigen was identified in bronchial and tracheal epithelial cells of all dogs and in alveolar macrophages of several dogs. Many dogs that were spontaneously infected with virus also developed bacterial pneumonia, and greyhound dogs with fatal spontaneous infection developed severe pulmonary hemorrhage with hemothorax. Virus-inoculated BALB/c mice developed tracheitis, bronchitis, bronchiolitis, and mild pneumonia in association with viral antigen in airway epithelial cells and in type 2 alveolar epithelial cells. Virus was not detected in extrarespiratory sites in any animals. The results indicate that canine influenza virus infection consistently induces acute tracheitis and bronchitis in dogs. Mice may be a useful model for some pathogenesis studies on canine influenza virus infection.
The genes for all influenza A viruses are thought to have originated in avian influenza viruses of shorebirds and wild waterfowl. 32 Influenza A viruses naturally infect an array of birds and mammals but exhibit varying degrees of host adaptation. Although influenza becomes strongly host adapted, interspecies transmission appears to be a common event. 24,32 There has been an emergence of a highly pathogenic avian H5N1 influenza virus with a transmission from birds to humans, as well as a novel swine-origin influenza (H1N1) that is transmitted human to human; both of which have heightened the biomedical community’s sensitivity to the dangers of interspecies influenza transmission and stimulated increased interest in factors that regulate interspecies transmissibility. 21,24
Influenza virus is an enveloped RNA virus with an eight-segmented, single-stranded, negative-sense genome belonging to the family Orthomyxoviridae. Major viral factors that influence pathogenicity and interspecies transmission include hemagglutinin (HA) receptor specificity and cleavability, 12,17,24 polymerase-mediated (PB1, PB2, PA) control of replication efficiency, neuraminidase efficiency in cleaving binding receptors to release the virus, 20 and nonstructural protein-mediated (NS1) control of innate cell defense mechanisms and hyperinduction of cytokines. 4,20,24 However, other genetic factors of the virus and host that control viral transmissibility remain poorly characterized. 15
In 2004, a H3N8 canine influenza virus was isolated from racing Greyhound dogs with severe respiratory disease, and it had greater than 96% nucleotide sequence identity to current circulating strains of equine influenza virus. 7 The virus was subsequently identified as being associated with outbreaks of severe respiratory disease in racing Greyhound dogs throughout the United States as well as in shelter and pet dogs. 10,23 The virus is highly contagious and has a high attack rate. Virus infection appears to be rarely fatal unless there is intercurrent disease such as bacterial pneumonia, except in racing greyhound dogs that appear to be more susceptible to fatal pneumonia with pulmonary and thoracic hemorrhage. 7,10,23,33 Sporadic cases of avian H5N1 and H3N2 influenza virus infection have also been associated with fatal disease in dogs. 28,29
The objectives of this study were (1) to characterize sites of virus replication and pathologic changes induced in dogs by experimental inoculation with canine H3N8 influenza virus and (2) to compare the experimentally induced lesions with those found in dogs following spontaneous infection. Finally, we inoculated BALB/c mice with canine influenza virus to determine whether mice were a suitable animal to use for molecular pathogenesis studies. A previous study indicated that mice were not susceptible to the H3N8 influenza virus circulating in horses in the 1980s. 13 We compared infection and disease in mice induced by canine influenza virus and a genetically more distant equine influenza virus isolated from horses in 1991. 19 Results are presented showing that canine influenza viral replication predominates in tracheal and bronchial epithelial cells to induce inflammatory lesions and that experimental infection in mice may be a suitable small animal model for some virus pathogenesis studies.
Materials and Methods
Animals
Racing Greyhound dogs with spontaneous infection
Information on the initial outbreak of respiratory disease in racing Greyhound dogs in north Florida, as well as references to other outbreaks, has been described. 7 Of the 22 clinically affected adult Greyhound dogs that died or were euthanatized with severe clinical disease, 5 were subjected to complete necropsies with histopathology at the University of Florida Veterinary Medical Center (UFVMC) by board-certified veterinary pathologists. One of the dogs was examined at necropsy by a local veterinary practitioner, and samples of lung were submitted to the UFVMC. Lung samples were taken at necropsy and submitted to the UFVMC clinical microbiology service. Samples of lung were also submitted for viral isolation to Cornell University Animal Health Diagnostic Laboratory.
A separate respiratory disease outbreak occurred in June 2006 at a racetrack kennel in Naples, Florida, during which 16 adult Greyhound dogs died (not previously described). Seven dogs (2.0 to 2.5 years old, male and female) were examined at necropsy and by extensive histopathology at the UFVMC. Lung samples were cultured for bacteria and mycoplasma. Lung samples were positive by polymerase chain reaction (PCR) for H3 sequences or positive by virus isolation.
Shelter and pet dogs with spontaneous influenza virus infection
Spontaneous disease outbreaks of canine influenza virus were identified throughout spring and summer of 2005 in Florida as previously described, with diagnosis based on detection of seroconversion (fourfold increase in hemagglutination inhibition titer to canine influenza virus) and PCR. 23 Eight dogs were submitted for necropsy at the UFVMC, for which 8 of 8 were positive by PCR testing of lung samples; in addition, canine influenza virus was isolated from 2 of 8. Six of the dogs originated from a north Florida animal shelter where an outbreak of respiratory disease occurred and included male and female dogs from 4 months to unknown adult age. They were mixed-breed dogs except for one 6-month-old female Australian shepherd dog. One of the shelter dogs died. The other 5 were euthanatized because of fever and chronic cough. The nonshelter dogs were pet dogs that died with signs of respiratory disease and were being temporarily housed with other dogs. One was a 9-year-old male Yorkshire terrier. The other pet dog was a 4-year-old male Pekinese dog.
Experimentally inoculated dogs
All animals were studied under experimental protocol approved by the University of Florida Institutional Animal Care and Use Committee. Twelve specific-pathogen-free Beagle dogs and mongrel dogs (16 to 24 months old, male and female) were purchased (Liberty Research Inc, Waverly, New York). Ten dogs were conditioned and inoculated with influenza virus A/canine/Florida/43/2004 (H3N8; canine/FL/04) that was produced and assayed in Madin–Darby canine kidney (MDCK) cells as described. 7 Beagle dogs (2 males and 2 females) received 106.6 50% tissue culture infectious dose (TCID50) virus with dogs under propofol anesthesia. Dose was split, with half given intranasally and half given intratracheally. Six mongrel dogs (3 male and 3 female) received 10 7 TCID50 virus under anesthesia by intratracheal injection. Nasal and oropharyngeal swabs were collected and assayed for virus on MDCK cells or processed for influenza virus matrix gene by real-time PCR as described. 7 Two Beagle dogs each were killed at 5 and 14 days after inoculation. Mongrel dogs were killed at each of the following days after inoculation: 1, 2, 3, 4, 5, 6. One Beagle dog was inoculated with 106.3 TCID50 influenza virus A/equine/Kentucky/1/1991 (H3N8; equine/KY/91) in 3 ml via intratracheal route and euthanatized at 3 days after inoculation. One uninoculated Beagle dog served as a control. Dogs were killed by intravenous injection with sodium pentobarbital and sodium phenytoin (Beuthanasia-D, Schering-Plough Animal Health Corp, 1 ml per 5 kg body weight).
BALB/c mice
Male BALB/c mice (22 to 24 g, 50 to 70 days old) were purchased from Charles River Laboratories (Charles River Laboratories International Inc, Wilmington, Massachusetts) and maintained in microisolator cages under biosafety level 2–plus conditions before inoculation with virus or tissue culture media. Mice were anesthetized with isoflurane and inoculated intranasally with 100 μL containing either 106.48 TCID50 canine/FL/04 or 106.48 TCID50 equine/KY/91 virus. Controls received saline intranasally. BALB/c mice were chosen because they are widely used in influenza A virus pathogenesis studies. 5,13,22 Mice were weighed and monitored daily. Mice were euthanatized by anesthesia with urethane and exsanguination and by pneumothorax before tissue collection. In studies with canine/FL/04 virus, 3 to 6 mice were studied per time point at 0, 1, 2, 3, 5, 7, 10, and 14 days after inoculation. Control mice (sham inoculation with saline; n, 3 to 6) were studied at 0, 7, and 14 days after inoculation. For parallel studies with equine/KY/91 virus, 3 to 6 mice were studied at 3, 5, and 7 days after inoculation, with 3 control mice studied at 3 days after sham inoculation.
Necropsy and Microbiological Procedures
Tissues—including lung, trachea, nasal mucosa, as well as brain, heart, liver, right lung, intestine, and kidney—were collected at necropsy for virology and morphologic studies. Segments from the right lung were fixed by immersion in 10% buffered formalin. The left lung was fixed by airway installation at 30 cm water pressure with dilute Karnovsky’s fixative for light and transmission electron microscopy as described. 3
Virus and Viral Assays
Canine/FL/04 virus for inoculation was a third-passage isolate produced by one passage in MDCK cells, as described. 7 Equine/KY/91 was produced by inoculating 9-day-old embryonated chicken eggs, which were harvested 2 days after inoculation because virus titers produced in MDCK were lower than that needed for a matching inoculum titer. Virus titer was determined by TCID50 assays as described below. Equine/KY/91 was selected because it has been sequenced and is genetically closer to the equine H3N8 viruses used by Kawaoka 13 in his studies with mice (ie, A/equine/Tennessee/5/1986) than the more recently circulating viruses thought to have been transmitted directly to dogs (eg, A/equine/Ohio/1/2003). 7,19 We expected to see differences in lesions induced by canine/FL/04 versus equine/KY/91 because of genetic difference in one or more of the 8 gene segments between the 2 viruses. Differences in lesions induced by the 2 viruses with known genetic difference could be used in reverse-genetic studies to identify genetic sequences important in disease pathogenesis.
Nasal and lung samples from dogs were assayed for infectious virus, and lung samples were analyzed for influenza matrix gene RNA using real-time quantitative PCR as described. 7,23 Samples from mice were assayed for infectious influenza virus by plaque assay, and lung, brain, and spleen samples were processed for influenza matrix gene RNA by qualitative PCR.
TCID50 assays were run after 10-fold serial dilutions of samples and inoculation onto confluent MDCK cells that had been grown in 24-well plates with Eagle’s minimum essential medium (EMEM) containing 10% fetal bovine serum (Sigma-Aldrich, St Louis, Missouri) and antibiotic/antimycotic (100 U penicillin, 100 μg streptomycin, and 250 ng Amphotericin B per ml media). Cells were rinsed twice with saline before inoculation. After virus adsorption for 60 minutes, cells were covered with EMEM containing antibiotic/antimycotic and 1 μg per ml porcine pancreatic trypsin. Wells were examined for cytopathic effect after 2 days of incubation. TCID50 titers were calculated by the Reed-Muench method 27 with 4 wells per dilution. Virus-specific cytopathic effect was confirmed by immunocytochemistry with antibody to H3 HA influenza A antigen.
Plaque assays were run similarly to TCID50 titer assays, as described previously, 7 except that 12-well plates were used and overlay contained 0.5% gelatin and 0.5% agarose. At 2 days after well inoculation in triplicate per dilution, cells were fixed with 4% neutral phosphate-buffered formaldehyde. The overlay was removed, and cell layers were stained with crystal violet before plaques were read to calculate average plaque number per well to calculate the titer.
Qualitative PCR for matrix gene expression was run on mouse lung, spleen, and brain samples following RNA extraction with a kit (RNeasy mini kit, QIAGEN Inc, Valencia, California), in a thermocycler using primers for the matrix gene: forward sequence, TGATCTTCTTGAAAATTTGCAG; reverse sequence, CCGTAGCAGGCCCTCTTTTCA. This was followed by separation of reactants on 2% agarose minigels and by identification of the 200–base pair product. For some paraffin-embedded tissue samples, RNA was extracted for qualitative PCR with an Ambion RecoverAll Total Nucleic Acid Isolation Kit (Applied Biosystems/Ambion, Austin, Texas).
Immunohistochemistry
Viral antigen was detected in paraffin sections by immunohistochemistry following deparaffinization and rehydration of sections. Sections were treated with a primary mouse monoclonal antibody to H3 HA influenza A antigen (MAB8254, Chemicon-Millipore, Billerica, Massachusetts). Primary antibody was detected using a STAT-Q three-step peroxidase staining system (Innovex Biosciences, Richmond, California), and slides were counterstained with hematoxylin. Limited immunohistochemistry was also run, with primary antibodies to influenza A nucleoprotein from the Centers for Disease Control and Prevention and with 2 primary antibodies from commercial suppliers. Because tested nucleoprotein antibodies yielded high background staining, all results are reported with the H3 antibody.
Transmission Electron Microscopy
Lungs were fixed by perfusion via the trachea at 30 cm water pressure with dilute Karnovsky’s fixative and processed as described. 3 Tracheal, bronchial, and pulmonary parenchymal blocks were embedded in Embed12-Araldite in JB-4 molds. Large sections were cut with Ralph knives and stained with toluidine blue. Areas of interest were cut out of the larger blocks and mounted on smaller blocks for ultrathin sectioning. Ultrathin sections were stained with lead citrate and uranyl acetate before examination by transmission electron microscopy. Lung samples were also processed from one Greyhound dog that was euthanized in moribund condition.
Results
Lesions and Distribution of Viral Antigen in Racing Greyhound Dogs and in Pet/Shelter Dogs Spontaneously Infected With Canine Influenza Virus
Racing Greyhound dogs
All 13 racing Greyhound dogs that died or were euthanatized with severe clinical disease had extensive areas of grossly detectable pulmonary hemorrhage (Fig. 1 ), often involving entire lobes or lung; the hemorrhage was confirmed through histologic examination. These dogs also had hemorrhage into the mediastinum and hemothorax with as much as 100 to 200 ml of free blood in the pleural cavity.
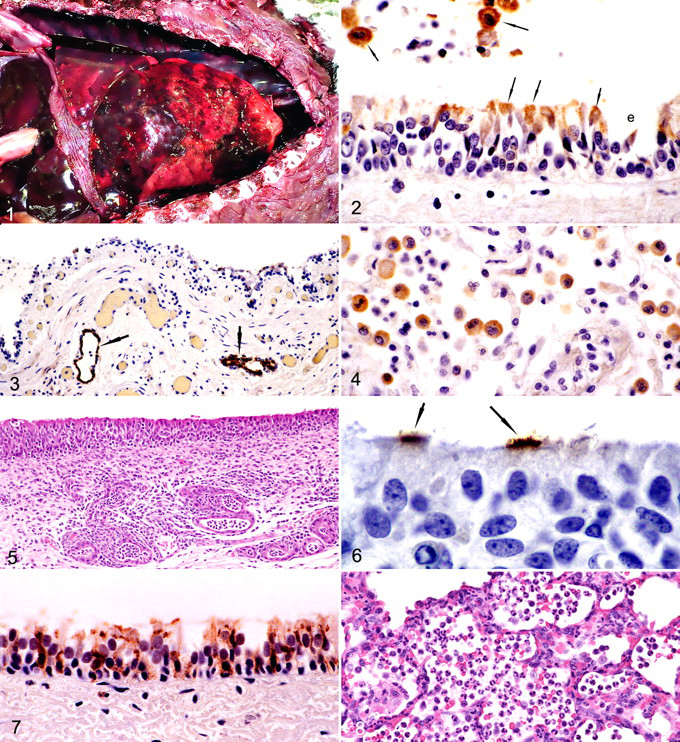
All 13 dogs had histologic evidence of tracheitis and bronchitis, characterized by (1) necrosis and erosion of surface epithelium or (2) hyperplasia of lining epithelial cells with intercurrent infiltration by neutrophils and small mononuclear inflammatory cells consistent with lymphocytes and macrophages (Figs. 2, 3). There was often either necrosis or hyperplasia of epithelial cells lining tracheal and bronchial glands. Transmission electron microscopy on bronchial samples from one dog confirmed necrosis of ciliated epithelial cells and inflammatory changes. Viral structures were not found. Of the 13 dogs, 2 had bronchiolitis, characterized by (1) epithelial necrosis and erosion or (2) epithelial hyperplasia with inflammatory cell infiltrates dominated by neutrophils and macrophages with fewer lymphocytes in bronchiolar walls and lumens. Nasal turbinates were examined from 11 of 13 Greyhound dogs, 9 or which had mild rhinitis characterized by epithelial necrosis or erosion and lymphocytic and macrophage infiltration of mucosa.
Transmission electron micrograph of bronchus of dog 5 days after inoculation with canine influenza virus. Cilia are clumped together (arrows) in an area where viral particles are budding from the membranes of ciliary shafts. Bar = 1 µm.
Transmission electron micrograph of the bronchus of dog 5 days after inoculation with canine influenza virus. Virion budding from cilia (arrow). Bar = 250 nm.
Mild to severe suppurative bronchopneumonia was present in 12 of 13 dogs. Lesions of bronchopneumonia were predominant in cranioventral locations, and the inflammatory cell infiltrates were airway oriented or diffuse lobule and dominated by neutrophils and macrophages. Alveolar exudates included abundant fibrin and red blood cells. In other areas of the lung, pulmonary hemorrhage was present without inflammatory cell infiltrates.
Influenza virus was identified in all 13 Greyhound dogs by viral isolation, PCR, and/or immunohistochemistry for viral antigen. Immunohistochemical localization of H3 antigen was identified in tracheal and bronchial surface epithelial cells (Fig. 2) as well as in epithelium of submucosal glands (Fig. 3). H3 antigen was multifocally found in bronchiolar epithelium. It was also identified in alveolar macrophages and in airway macrophages of one dog (Figs. 2, 4). No other viruses were isolated.
Transmission electron micrograph of bronchus of dog 5 days after inoculation with canine influenza virus. Cilia with 3 axonemes in an area where viral particles are budding from the ciliated cell (not shown). Bar = 250 nm.
Bacteria were identified in the respiratory tract by Brown and Brenn Gram stain and/or isolated by bacterial culture in all 13 dogs. The most common bacterial isolates were as follows: Streptococcus equi subsp zooepidemicus (7 dogs), Escherichia coli (5 dogs), Pasteurella multocida (2 dogs), and Mycoplasma sp (concurrent with other isolates in 2 dogs).
All dogs examined had mild to moderate lymphoid hyperplasia in tracheobronchial lymph nodes. Other notable concurrent nonrespiratory lesions were found in 2 dogs. One Greyhound dog had acute multifocal arterial fibrinoid necrosis in heart and lung and multifocal necrotizing encephalitis and myocardial necrosis. Immunohistochemistry failed to reveal influenza viral antigen in association with lesions in the arteries, brain, or heart. Qualitative PCR on RNA extracted from paraffin sections of heart and brain were negative for matrix RNA. The second dog had mild multifocal renal tubular necrosis.
Shelter and pet dogs
All 8 shelter and pet dogs had tracheitis and bronchitis, comparable to that observed in Greyhound dogs, and they were positive for virus by PCR. Two virus isolates recovered from these dogs (A/canine/Jacksonville/2005 and A/canine/Miami/2005) have been described in detail. 23 None of the dogs had severe pulmonary and thoracic hemorrhage that occurred in Greyhound dogs. Of 8 dogs, 3 had moderate to severe suppurative to histiocytic bronchopneumonia associated with bacterial infection (dog No. 1, S equi subsp zooepidemicus; dog No. 2, Enterococcus faecium; dog No. 3, E coli and Enterococcus sp). Three dogs had died spontaneously; 2 of these had bronchopneumonia and one had infestation with Dirofilaria immitis. Five dogs were euthanatized primarily because of chronic cough that was unresponsive to antibiotics; bronchitis and tracheitis were the only significant findings at necropsy. In one of the dogs with tracheitis, bronchitis, and bronchopneumonia, bronchiolitis and rhinitis were found and viral antigen was localized in tracheal, bronchial, and nasal epithelium.
Experimental Canine Influenza Virus Infection in Dogs
The most common lesions in experimentally inoculated Beagle and mongrel dogs were tracheitis and bronchitis with accompanying inflammation of underlying submucosal glands (Figs. 5-7). Bronchiolitis was less commonly observed and mild. Pneumonia in inoculated dogs was limited in distribution and duration. Dogs showed minimal or no clinical signs and did not develop coughs or nasal discharges. Mongrel dogs had transient fevers from 24 to 48 hours after inoculation (39.3 to 39.5°C). Beagle dogs did not develop fevers.
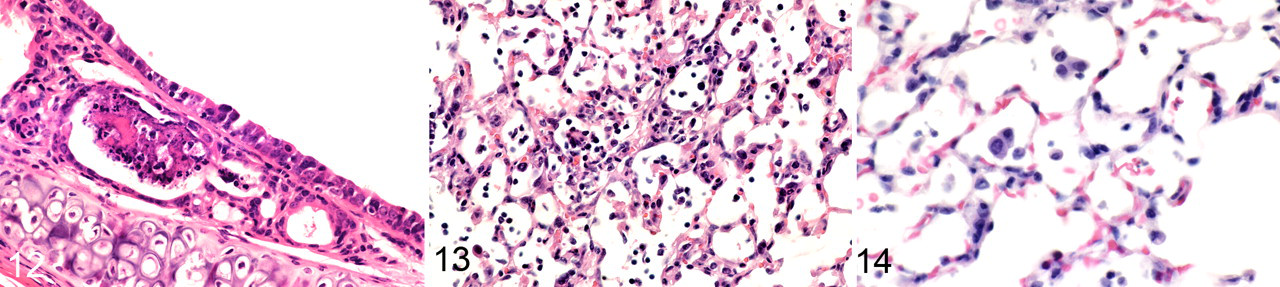
Tracheitis/bronchitis
As early as 1 day after inoculation, lesions were present multifocally throughout trachea and bronchi, as characterized by individual epithelial cell necrosis or rare apoptosis with intercurrent infiltration by neutrophils among epithelial cells and in the lamina propria and submucosa. Larger areas of epithelial cell necrosis and erosions were observed at 2, 3, 4, and 5 days after inoculation in the surface epithelium. Only minimal multifocal epithelial necrosis was observed at 6 days after inoculation. Epithelial hypertrophy and hyperplasia were observed intermixed with epithelial necrosis at 4, 5, and 6 days after inoculation (Fig. 5). Lymphocytes and macrophages were increasing components of the inflammatory cell infiltrates observed at 2, 3, 4, and 5 days after inoculation and were associated with edema of the lamina propria and submucosa.
Inflammation of the submucosal glands in bronchi and bronchioles was characterized by epithelial cell necrosis or loss of normal cuboidal glandular cells, replacement by flattened low cuboidal cells, and infiltration of the epithelium and surrounding submucosa by neutrophils and lymphocytes (Fig. 5). Glandular epithelial hyperplasia was observed at 4, 5, and 6 days after inoculation. Tracheal and bronchial inflammatory lesions were almost entirely resolved at 14 days after inoculation, with only multifocal lymphoid aggregates of low cellular density being present in lamina propria.
Bronchiolitis
At 1 and 2 days after inoculation, low-density aggregates of neutrophils and macrophages were present in bronchiolar lumens and occasionally in lamina propria. Multifocal small erosions of bronchiolar epithelium were found at 4 and 5 days after inoculation and were associated with small numbers of lymphocytes and macrophages and with mild multifocal edema in the bronchiolar wall.
Pneumonia
Pneumonia was present at 1, 2, and 3 days after intratracheal inoculation in cranioventral lung areas and was characterized by inflammatory cell infiltrates dominated by neutrophils in alveolar spaces and interalveolar septa around terminal and respiratory bronchioles (Fig. 8). Gross lesions were visible at 2 and 3 days after inoculation when approximately 0.5 and 8.0% of the lung, respectively, had reddened areas corresponding to pneumonia.
At 4 days after inoculation, a similar cranioventral and proximal acinar distribution of lesions was found histologically. However, the density of inflammatory cells was much lower at this later time. Macrophages were the predominant inflammatory cell type in alveoli, and there was thickening of interalveolar septa and mild hyperplasia of alveolar type 2 epithelial cells. At 5 and 6 days after inoculation, mild multifocal thickening of interalveolar septa in proximal acinar areas was observed with hyperplasia of type 2 alveolar epithelial cells and multifocal aggregates of macrophages in alveolar spaces.
Rhinitis
Beagle and mongrel dogs had mild rhinitis of comparable severity. These lesions were characterized by mild multifocal epithelial cell necrosis and infiltration of mucosal epithelium and lamina propria by low-density aggregates of neutrophils and lymphocytes from 2 to 6 days after inoculation.
Viral isolation/viral PCR
Virus was recovered from lung and trachea of all virus-inoculated dogs that were necropsied at 1, 2, 3, 4, and 5 days after inoculation. Virus was not recovered from any of the dogs that were necropsied at 6 and 14 days after virus inoculation. All lung samples were positive for matrix gene expression by PCR from virus-inoculated dogs necropsied at 1, 2, 3, 5, and 6 days after inoculation.
Immunohistochemistry for viral antigen and transmission electron microscopy
Viral antigen was multifocally present in bronchial and tracheal ciliated epithelial cells at 1 and 2 days after inoculation (Fig. 6). At 3, 4, 5, and 6 days after inoculation, viral antigen was present in surface ciliated, nonciliated, or mucous cells, as well as basal epithelial cells (Fig. 7). Submucosal gland epithelial cells were also antigen positive. Antigen was found in occasional bronchiolar epithelial cells at 1 day after virus inoculation but not at later times. Viral antigen was present in widely scattered alveolar macrophages in proximal acinar areas in dogs at 1 to 3 days after inoculation.
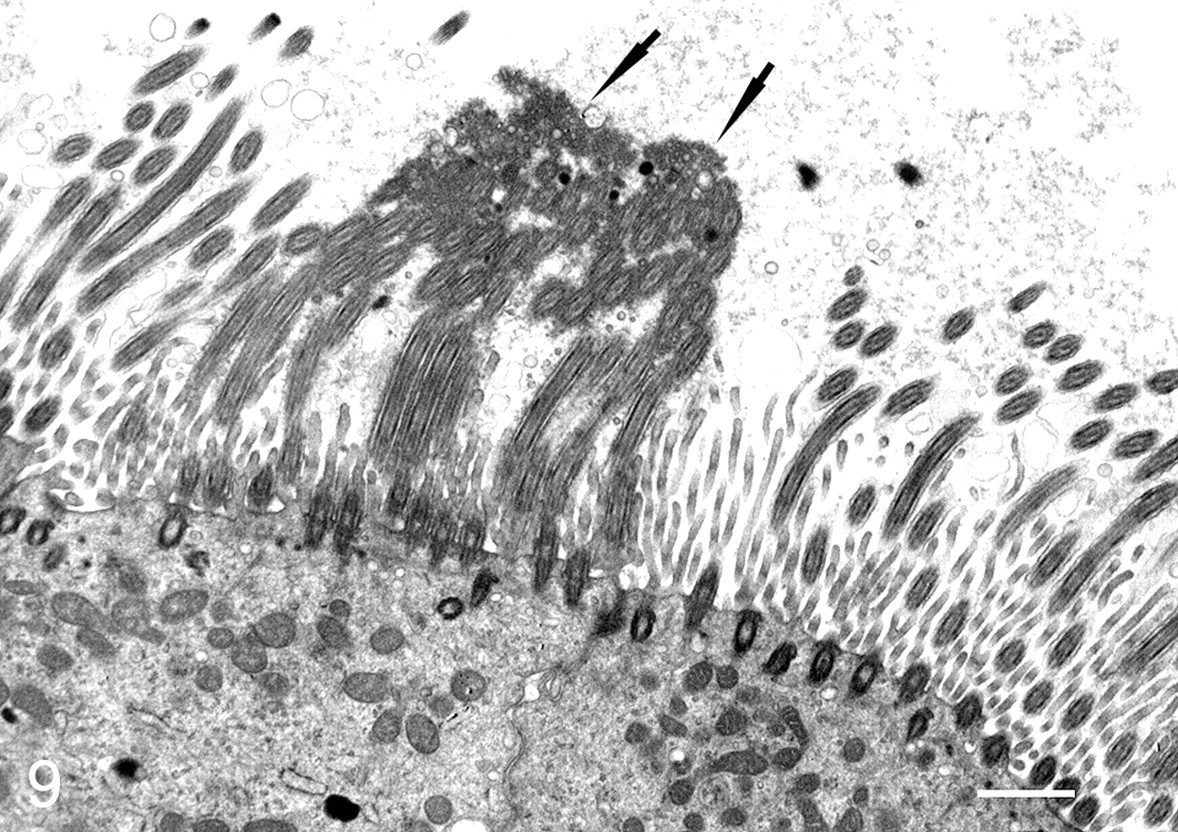
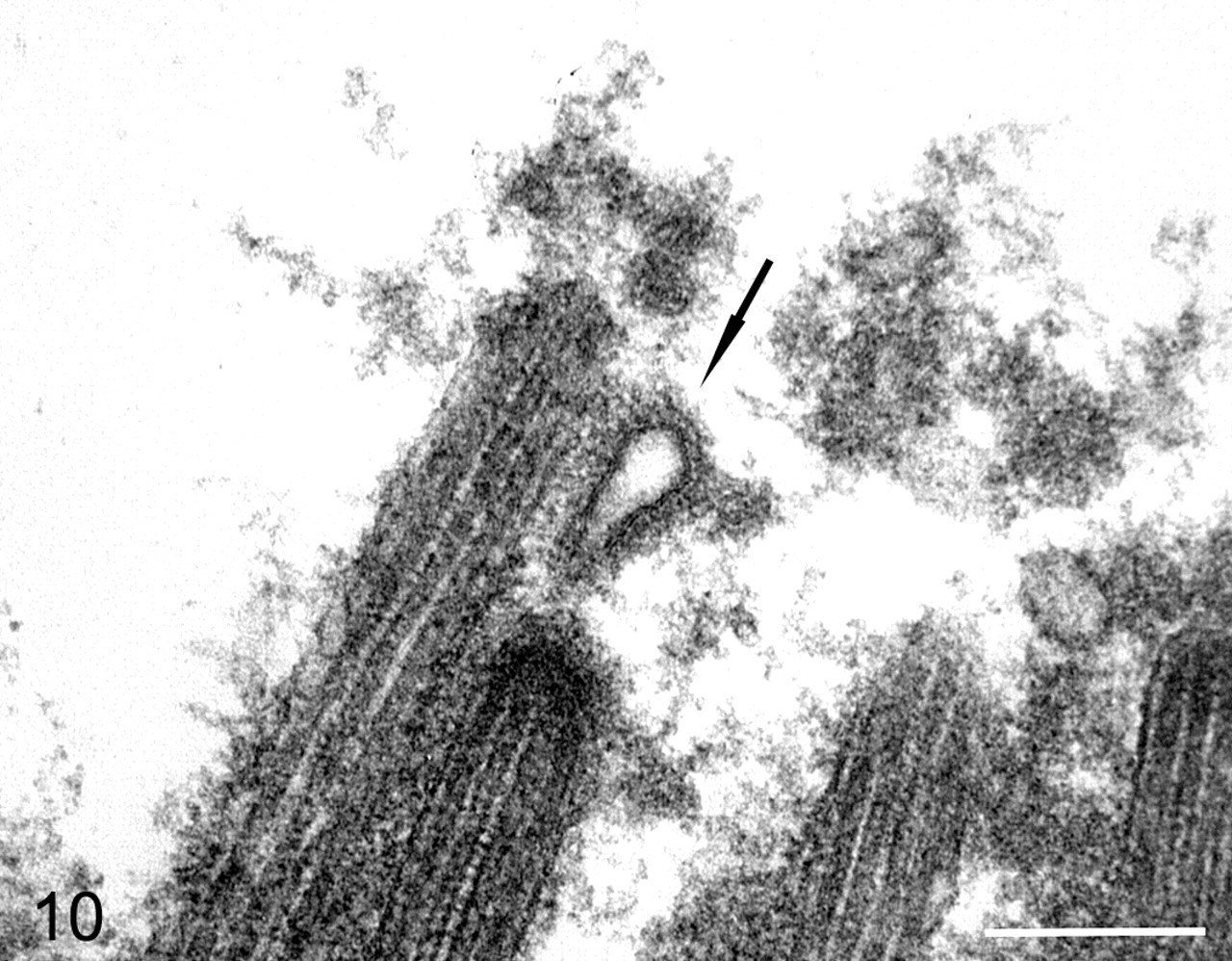
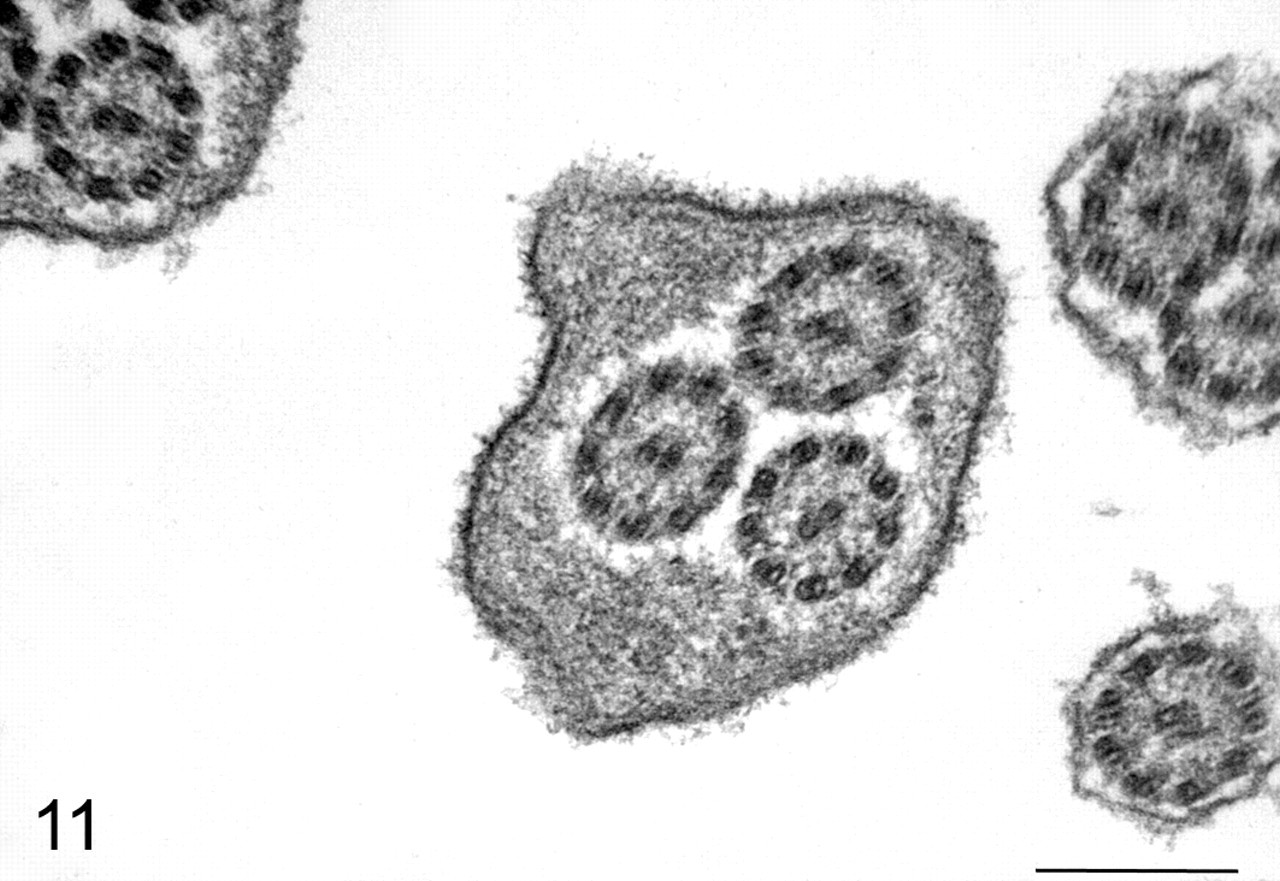
By transmission electron microscopy, viral structures were identified budding from ciliary shafts in both tracheal and bronchial ciliated epithelial cells (Figs. 9, 10). In many ciliated cells in areas where budding virions were found, there was fusion of ciliary shafts and formation of compound cilia with multiple axonemes (Figs. 9, 11). These lesions were not identified in control dogs or in virus-inoculated dogs where budding virions were not found. Other ultrastructural findings were confirmatory of the cell injury, and inflammatory changes were noted in the histologic studies.
Only one Beagle dog was inoculated with equine/KY/91 influenza virus. Neither gross nor histologic lesions were present at necropsy 3 days after inoculation. No infectious virus was recovered from trachea samples.
Experimental Inoculation of BALB/c Mice
Clinical signs
Mice inoculated with canine/FL/04 showed few clinical signs other than slight ruffling of fur from 2 to 5 days after inoculation. They lost an average of 5% body weight by 5 days after inoculation but recovered and had body weight gains comparable with control mice by 14 days after inoculation. Mice inoculated with equine/KY/91 showed no clinical signs and had no significant loss of body weight compared with control mice.
Lesions after canine/FL/04 virus inoculation
Mice inoculated with canine/FL/04 influenza virus developed tracheitis, bronchitis, and bronchiolitis as early as 2 days after inoculation. Lesions at this time were characterized by mild to moderate multifocal necrosis of surface epithelial cells and tracheal submucosal gland epithelial cells (Fig. 12), accompanied by infiltration of lymphocytes, plasma cells, and neutrophils in the epithelium and lamina propria. Bronchiolar epithelial necrosis with erosion of epithelium and intercurrent epithelial hyperplasia was observed primarily in proximal large-caliber bronchioles with lesser involvement of terminal bronchioles. Inflammatory changes and epithelial necrosis were most severe from 2 to 5 days after inoculation and less severe at 7 days after inoculation when airway epithelial hyperplasia was prominent. By 10 days after inoculation, airway reparative changes were largely complete, with resolution of inflammation and redifferentiation of ciliated cells in the mucosal surface. Mild multifocal pneumonia in proximal acinar areas was observed from 2 to 5 days after inoculation and was characterized by thickening of interalveolar septa with edema and hyperplasia of type 2 alveolar epithelial cells, as well as by low-density alveolar aggregates of macrophages, lymphocytes, and few neutrophils (Fig. 13). Virus-inoculated mice had multifocal to locally extensive rhinitis from 1 to 5 days after inoculation characterized by epithelial necrosis and erosion and by infiltration of epithelium, lamina propria, and lumens by neutrophil- and lymphocyte-dominated inflammatory cell infiltrates. Multifocal low-density lymphocyte aggregates were present in the mucosa at 7, 10, and 14 days after inoculation.
Immunohistochemistry
Viral antigen was identified by immunohistochemistry in ciliated and nonciliated surface epithelial cells of trachea, bronchi, and bronchioles. In terminal bronchioles, viral antigen was infrequently found in nonciliated bronchiolar epithelial cells. Viral antigen was identified with immunohistochemistry in occasional type 2 alveolar epithelial cells. Alveolar macrophages were not positive for virus antigen.
Lesions after equine/KY/91 virus inoculation
Mice inoculated with equine/KY/91 influenza virus developed tracheitis, bronchitis, and bronchiolitis that was similar to that in mice inoculated with canine/FL/04. However, erosions were less severe, and inflammatory cell infiltrates were generally lower in density. Mice inoculated with equine/KY/91 virus developed mild multifocal pneumonia at 3 and 5 days after inoculation, as characterized by lower-density aggregates of inflammatory cells in alveolar spaces and milder thickening of interalveolar septa (Fig. 14) than that seen in mice inoculated with canine/FL/04. Inflammatory lesions were largely resolved by 7 days after inoculation.
Virus isolation
Infectious virus was recovered and assayed by infectious plaque assay from mice inoculated with canine/FL/04 influenza from 1 through 5 days after inoculation (Fig. 15). Infectious virus was not detected by infectious plaque assay from lungs of mice inoculated with equine/KY/91. However, lung samples were positive for matrix gene RNA by PCR at 1 through 5 days after inoculation with canine/FL/04 and at 3 and 5 days after inoculation with equine/KY/91.
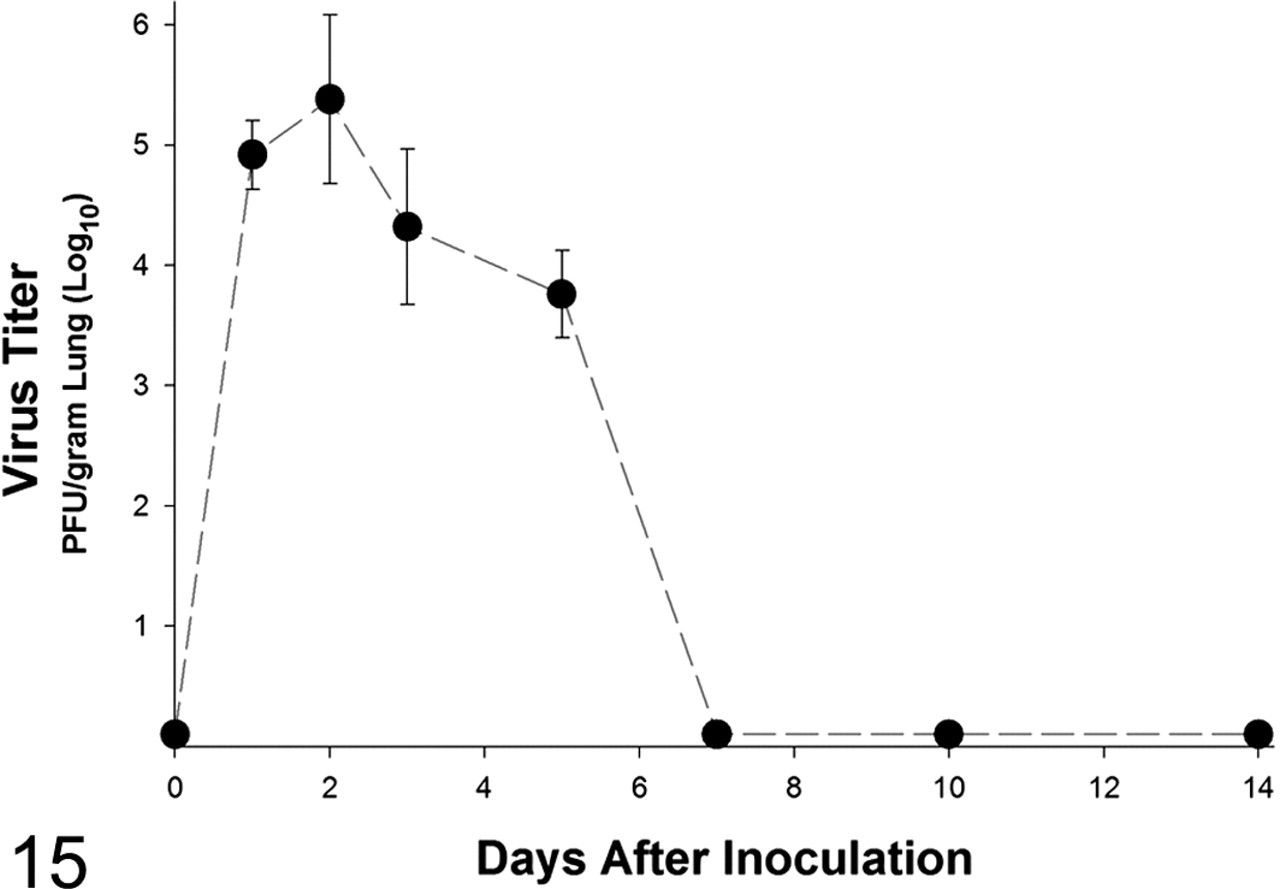
Lung virus titer from BALB/c mice inoculated with canine influenza virus. n = 3 at each point.
Discussion
Major objectives of this study were (1) to characterize sites of virus replication and pathologic changes induced in dogs by experimental inoculation with canine H3N8 influenza virus and (2) to compare the experimentally induced lesions with those found in Greyhound and pet/shelter dogs following spontaneous infection. We also wanted to determine whether mice developed lesions comparable with those in dogs following experimental infection and if mice might be a suitable animal to use for molecular pathogenesis studies with recombinant virus in reverse-genetic studies.
Experimental inoculation with canine/FL/04 influenza virus induced consistent tracheitis and bronchitis with involvement of surface epithelium as well as submucosal gland epithelium. Experimentally induced tracheal and bronchial lesions were comparable with those observed in spontaneously infected racing Greyhound dogs and shelter/pet dogs. Only mild bronchiolitis and pneumonia were induced in dogs by experimental virus inoculation in this study, whereas pneumonia was a common finding in spontaneously infected Greyhound dogs and in shelter/pet dogs.
The findings of consistently induced tracheitis and bronchitis were in agreement with immunohistochemical localization of influenza HA antigen in surface ciliated and nonciliated epithelial cells, as well as in epithelial cells lining submucosal glands in trachea and bronchi. Virions were also observed ultrastructurally budding from ciliary shafts in bronchi and trachea and were associated with ciliary structural abnormalities.
Viral antigen was infrequently found in bronchiolar epithelial cells and in alveolar macrophages of experimentally infected dogs. The low amount of detectable viral antigen in the lower respiratory tract was in agreement with the findings of induction of only mild bronchiolitis and pneumonia. Minimal viral antigen was observed in bronchiolar and alveolar areas in the experimentally inoculated dogs and the spontaneous canine influenza cases.
The discrepancy between commonly finding pneumonia in dogs spontaneously infected with canine H3N8 influenza virus and minimal induction of pneumonia in experimentally inoculated dogs can be explained by at least 2 mechanisms: First, canine influenza virus inefficiently replicates in alveolar tissue compared with airway epithelial cells, and other nonviral factors are responsible for pneumonic changes in dogs with spontaneous virus infection; second, the virus isolate we chose for experimental inoculation has attenuated virulence compared with wild virus.
The simplest explanation for the observed sites of viral antigen localization and lesion distribution in experimentally inoculated dogs is that canine influenza virus replicates to highest levels in tracheal and bronchial epithelium and that it replicates poorly in bronchioles and in alveolar tissue. Among the most important factors regulating sites of viral replication, levels of viral replication, and viral pathogenesis for type A influenza viruses in mammals are viral HA sequence, host cell receptor type, viral polymerases (PB1, PB2, PA), viral neuraminidase, and viral nonstructural proteins (NS1). 1,22
Viral HA binds to host receptors with oligosaccharides ending in sialic-acid α-2,6-galactose (SAα-2,6gal) or sialic-acid α-2,3-galactose (SAα-2,3gal). There are species and topographical cell differences in density of SAα-2,6gal and SAα-2,3gal cell receptors in the respiratory tract. Influenza viral HAs may bind predominantly to either SAα-2,6gal or SAα-2,3gal cell receptors, although some viruses can bind to both. 31 Studies with human and mouse cells indicate that tracheal ciliated cells and type 2 alveolar epithelial cells both express SAα-2,3gal receptors, although human ciliated cells also express SAα-2,6gal receptors. 12 Nonciliated tracheal epithelial cells express SAα-2,6gal receptors. 12 Initial studies indicate that dogs have both SAα-2,6gal and SAα-2,3gal receptors on their tracheal epithelial cells. 8 Binding studies with avian H5N1 virus and lectin-blocking studies indicate that dogs have SAα-2,3gal receptors in bronchiolar and alveolar epithelial cells. 14 Mutations in viral HA resulting in changes as few as two amino acids can abolish the transmission of highly infectious 1918 H1N1 influenza among ferrets. 31
Viral neuraminidase is important as a sialidase to promote viral release from host cells and spread; it also promotes HA cleavage. 1,32 Neuraminidase has demonstrated important virulence-enhancing properties in studies with 1918 influenza reassortants. 1,22
Influenza virus RNA synthesis is mediated by a heterotrimeric RNA-dependent RNA polymerase composed of PA, PB1, and PB2 proteins. Polymerase complex components are linked with the capacity of influenza virus to replicate efficiently in cells in a host-specific manner, and virulence of the 1918 pandemic influenza virus has been linked with the PB1 gene. 1 Single mutations in PB1-F2 resulting in a single amino acid substitution have been linked with increased pathogenicity of 1918 influenza virus and avian H5N1 influenza in a mouse model. 5
Influenza NS1 gene plays several roles in viral pathogenesis. The most widely recognized effects are to antagonize host cell interferon-α/β responses and to inhibit activation of host innate antiviral responses. 1 NS1 variants may also play a role in downregulating antiviral actions of interferon and TNF-α in infected host cells. 1
Although our studies indicate that canine influenza virus has limited ability to replicate in alveolar tissue and induce pneumonia in adult dogs, Deshpande et al 9 showed that pneumonia can be reliably induced using 20-fold higher concentrations of virus in young dogs (14- to 15-week-old puppies).
It is well recognized that passage of influenza virus in tissue culture and eggs can induce mutations that might reduce virulence of the virus when reintroduced into animals. 2,11 However, this appears to be an unlikely explanation for the observed induction of only mild pneumonia in dogs experimentally inoculated with canine influenza virus. The virus isolates we used had only four passages in MDCK cells. Furthermore, the spontaneous cases of canine influenza virus infection we reviewed were sources of 3 canine influenza virus isolates described (A/canine/Florida/43/2004, A/canine/Jacksonville/2005, and A/canine/Miami/2005). 7,23 In none of these spontaneously infected dogs was viral antigen localized in alveolar tissue other than in low numbers of macrophages.
Additional factors might be acting to promote pneumonia in dogs with canine influenza virus infection, when only small amounts of viral antigen can be detected in macrophages at the alveolar level. Such factors may include secondary bacterial infection and increased expression of proinflammatory molecules at the alveolar level in response to low levels of virus infection.
Influenza virus infection has been long recognized as predisposing to secondary bacterial infection. Secondary bacterial pneumonia was probably a major contributing factor in most of the 40 to 50 million human deaths in the 1918 influenza pandemic. 18,30 Influenza virus infection is thought to predispose to secondary bacterial infection through mechanisms that include epithelial damage and decreased mucociliary clearance, alterations of the innate and acquired immune responses, and induction of macrophage death. 16,18 In our spontaneous disease cases, bacteria were often recovered from lungs or identified in pneumonic lungs by special stains. No single bacterial agent was consistently associated with pneumonia in this study, and gram-negative and gram-positive organisms were both recovered, with S equi subsp zooepidemicus and E coli being the 2 most common isolates. There is evidence from the ultrastructural findings that influenza virus induces temporary disruption of ciliated cells through necrosis and erosion, as well as ciliary structural abnormalities that would be expected to disrupt mucociliary clearance. There is evidence from our immunohistochemical studies as well as in vitro inoculation studies that canine influenza virus replicates to a limited extent in macrophages 25 and could be interfering with bacterial defense by those cells at the alveolar level.
Severity of pneumonia and other disease manifestations in influenza virus infection in humans and other animals is at least in part mediated by induced or accentuated expression of proinflammatory cytokines. 4,16,22 Studies with dog alveolar macrophages have demonstrated that canine influenza virus replicates to a low level in macrophages, that the virus is a potent inducer of TNF-α, and that bacterial components further upregulate cytokine release after viral infection. 25,26
Studies on racing Greyhound dogs spontaneously infected with canine H3N8 influenza virus indicate that these dogs are highly susceptible to pulmonary and other thoracic hemorrhage following influenza virus infection. 7,33 As such, racing Greyhound dogs may have a greater propensity for hemorrhage in this inflammatory condition owing to systemic factors such as breed-associated hypertension, 6 increased virus replication in the respiratory system, or enhanced local cytokine release from infected macrophages and other cells. 16 Experimental studies with Greyhound dogs or their cells are needed to establish which, if any, of the preceding mechanisms are responsible for their apparent high susceptibility to influenza virus-induced pulmonary and thoracic hemorrhage. Pulmonary hemorrhage was a frequent finding in fatal influenza virus infection in humans during the 1918 pandemic, and it is found in fatal seasonal influenza infection. 30
Events and mechanisms responsible for the successful adaptation and transmission of equine influenza virus to dogs and subsequent transmission between dogs are poorly understood. 7,23 Analysis of important viral genetic mutations that allowed the virus of equine origin to adapt to dogs will eventually require a reverse-genetic approach and experimental infection of animals. Researchers have amply demonstrated the utility of the mouse model for viral genetics and pathogenesis studies with influenza viruses that infect humans. 1,16 We wanted to determine whether inoculation of BALB/c mice with canine H3N8 influenza virus would be a potentially useful model for canine influenza virus studies. Lesions in trachea, bronchi, and large-caliber bronchioles induced in mice by canine influenza virus were comparable with those in dogs with involvement of surface epithelium and submucosal glands. Only mild pneumonia was induced in mice. We can reasonably conclude that the overall pattern of virus injury in the respiratory tract induced by canine influenza virus is generally similar between dogs and mice and that the mouse may be a useful experimental model for some canine influenza virus studies.
Canine influenza virus originated from a direct transmission of equine virus to dogs, and recently circulating viruses in horses (eg, equine/Ohio/1/2003) have nucleotide homology that is 99% or greater with canine/FL/04 used in this study. 7,19 We wanted to use an equine virus in these studies that was more genetically distant, and we selected equine/KY/91 with only 97 to 98% nucleotide homology to canine/FL/04. Equine/KY/91 has homology that is comparable with the virus used by Kawoaka in his 1991 study (equine/Tennessee/5/1986). 13,19
Only mild tracheitis, bronchitis, and bronchiolitis were induced by equine/KY/91 virus, and no weight loss was observed following infection. In contrast, more severe airway lesions, as well as pneumonia, were observed in mice inoculated with canine/FL/04; mice also lost 5% body weight after inoculation. It is interesting that the one dog inoculated with equine/KY/91 failed to develop airway lesions or pneumonia.
Our studies with equine influenza virus in mice and one dog are preliminary and should be interpreted with caution. The lower pathogenicity of equine/KY/91 in mice and possibly dogs could be related to important genetic differences between the older equine virus and the 2004 canine virus, and such differences could influence interspecies transmission and adaptation to a new host. These genetic variations would be most optimally investigated with reverse-genetic studies. 5,22,31 Alternatively, the differing responses in mice and dogs may be more a reflection of differences in pathogenicity between a low-passage pathogenic isolate from dogs and a laboratory reference strain of influenza virus that has become adapted to eggs and lost important pathogenesis gene sequences through mutation and adaptation by serial passage in eggs. 2,11 More studies with different equine viruses—both old and contemporary circulating viruses with closer sequence to the canine virus—need to be made to interpret the findings and better understand genetic determinants of transmission and new host adaptation.
In conclusion, our studies indicate that the newly emerged canine influenza virus consistently induces acute tracheitis and bronchitis in dogs. Mice may be a useful model for some genetic and pathogenesis studies on canine influenza virus.
Footnotes
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
The authors declared the following financial support for the research and/or authorship of this article: Supported in part by a State of Florida Pari-Mutuel Wagering Trust Fund research grant and by College of Veterinary Medicine Consolidated Research Funds.