
Abstract
Mesenchymal stem cells (MSCs) hold great potential as a regenerative therapy for stroke, leading to increased repair and functional recovery in animal models of cerebral ischaemia. While it was initially hypothesised that cell replacement was an important mechanism of action of MSCs, focus has shifted to their paracrine actions or the so called “bystander” effect. MSCs secrete a wide array of growth factors, chemokines, cytokines and extracellular vesicles, commonly referred to as the MSC secretome. There is evidence suggesting the MSC secretome can promote repair through a number of mechanisms including preventing cell apoptosis, modulating the inflammatory response and promoting endogenous repair mechanisms such as angiogenesis and neurogenesis. In this review, we will discuss the in vitro approaches currently being employed to drive the MSC secretome towards a more anti-inflammatory and regenerative phenotype. We will then examine the role of the secretome in promoting repair and improving recovery in preclinical models of cerebral ischaemia.
Introduction
Stroke is a major global health problem with limited treatment options which leads to around 6.7 million deaths annually. 1 For the 33 million people living with stroke, a significant proportion have some disability. 2 Current treatments for acute ischaemic stroke are based on reperfusion through thrombolysis or endovascular therapy. Both approaches are very effective and have led to significant re-organisation of acute stroke services to allow greater access to these treatments. However, due to the narrow therapeutic window for administration of tPA (< 4.5 h of symptom onset), only 5% of patients in the UK receive thrombolysis 3 and an estimated 10% would be eligible for endovascular clot retrieval assuming national coverage, 4 which is still not the case. Therefore, there is much interest in developing regenerative therapies to alleviate the disability caused by stroke.
One promising candidate being widely investigated as a cell therapy for ischaemic stroke is mesenchymal stem/stromal cells (MSCs), multipotent cells first described by Friedenstein and colleagues in the 1960s and 1970s. 5 While initially found in bone marrow, MSCs have since been isolated from most postnatal organs 6 including adipose tissue, 7 dental pulp, 8 lungs, liver, spleen and brain.9,10 MSCs are also present in foetal tissues such as placenta, umbilical cord 11 and Wharton’s jelly. 12 The International Society for Cellular Therapy (ISCT) has defined the minimum criteria for MSCs as: adherence to tissue culture plastic; multipotency as demonstrated by in vitro differentiation into osteoclasts, adipocytes and chondroblasts; expression of surface markers CD73, CD90 and CD105; and negative for CD34, CD45, CD14 or CD11b, C79α or CD19 and HLA-DR. 13
A large number of clinical trials (794 as of January 2018) have been conducted or are ongoing to investigate MSCs as a potential therapy for a wide range of diseases including graft versus host disease, haematological malignancies, diabetes, and neurological diseases such as Alzheimer’s disease and amyotrophic lateral sclerosis.14,15 More specifically, a number of phase I/II clinical trials have suggested MSCs are a safe and feasible therapy for stroke.16–21 MSCs are immune evasive 22 and less immunogenic than many other cell types due to low expression of majority histocompatibility complex class I molecules. 23 In support of this, a meta-analysis conducted by Lalu et al. 14 found no association between acute infusional toxicity and MSC treatment overall and no adverse events in the 13 studies that used allogeneic cells. Thus, allogeneic transplantation without immunosuppressive therapy appears to be safe which has numerous advantages over autologous therapies including decreased cost and time to administration. 23
Numerous preclinical studies have demonstrated that treatment with stem cells, including MSCs, promotes functional recovery in rodent models of cerebral ischaemia. Although it was thought initially that the principle mechanism of therapeutic action of stem cells was direct replacement of dead and injured cells, this has been largely disregarded as very few cells reach the site of injury, engraft and survive long term.24,25 Following administration by intravenous (IV) or intra-arterial (IA) injection, the vast majority of MSCs become entrapped in the lungs within 48 h.26,27 Li et al. 28 reported that around 4% of cells were present in the ischaemic brain of rats 14 days after tail vein injection. Additionally, only a small percentage (<10%) of transplanted MSCs differentiate and express neuronal markers such as NeuN and MAP-2.29–32 To further disregard the cell replacement hypothesis, MSCs lack expression of the voltage-gated ion channels required for generating action potentials. 33 Despite this, MSC treatment leads to significant improvements in functional outcomes and can occur independently of cell migration to the ischaemic brain.28,34 There is growing evidence to support the paracrine actions of MSCs, also known as the bystander effect, in improving outcome in preclinical models of stroke. MSCs secrete a wide range of chemokines, cytokines, growth factors and extracellular vesicles (EVs) collectively termed the secretome.
In this review, we will firstly discuss in vitro approaches to modifying the MSC secretome to enhance a more anti-inflammatory and regenerative phenotype. We will then look at the involvement of the MSC secretome in promoting repair mechanisms, modulating inflammation and improving functional outcomes in preclinical models of cerebral ischaemia.
Approaches to enhancing the MSC secretome
MSCs secrete numerous growth factors, chemokines and cytokines including vascular endothelial growth factor (VEGF), insulin-like growth factor 1 (IGF-1), basic fibroblast growth factor (bFGF), transforming growth factor beta-1 (TGF-β1), nerve growth factor (NGF), placental growth factor (PGF), stromal-derived growth factor (SDF-1/CXCL12), monocyte chemoattractant protein-1 (MCP-1/CCL2), interleukin-6 (IL-6), IL-8, IL-10 and IL-13.35–38 There is some heterogeneity in the secretome of different populations of MSCs. Adipose-derived MSCs were reported to have higher mRNA expression of VEGF-D, IGF-1 and IL-8, while dermal sheath and dermal papilla-derived cells secreted higher concentrations of CCL2 and leptin than other populations.
39
Additionally, Du et al.
40
found increased expression levels of HGF (hepatocyte growth factor), bFGF, IL-6, IL-8, IL-1α and IL-1β in placenta-derived MSCs and in bone marrow-derived populations, VEGF-A, NGF and angiogenin were higher. Thus, MSCs secrete a number of factors that could promote angiogenesis and neurogenesis, prevent apoptosis and modulate inflammatory responses. The MSC secretome therefore has great potential as a regenerative therapy for stroke and a number of strategies have been employed to further enhance this reparative capacity (Figure 1).
Summary of in vitro approaches that have been utilised to enhance the therapeutic potential of mesenchymal stem cell secretome. BDNF: brain-derived neurotrophic factor; FGF: fibroblast growth factor; G-CSF: granulocyte-colony stimulating factor; HGF: hepatocyte growth factor; IFN-γ: interferon gamma; IGF: insulin-like growth factor; IL: interleukin; MMPs: matrix metalloproteinases; NGF: nerve growth factor; PGE2: prostaglandin E2; TGF-β: transforming growth factor beta; TNF-α: tissue necrosis factor alpha; TSG-6: TNF-α–stimulated gene 6 protein; VEGF: vascular endothelial growth factor.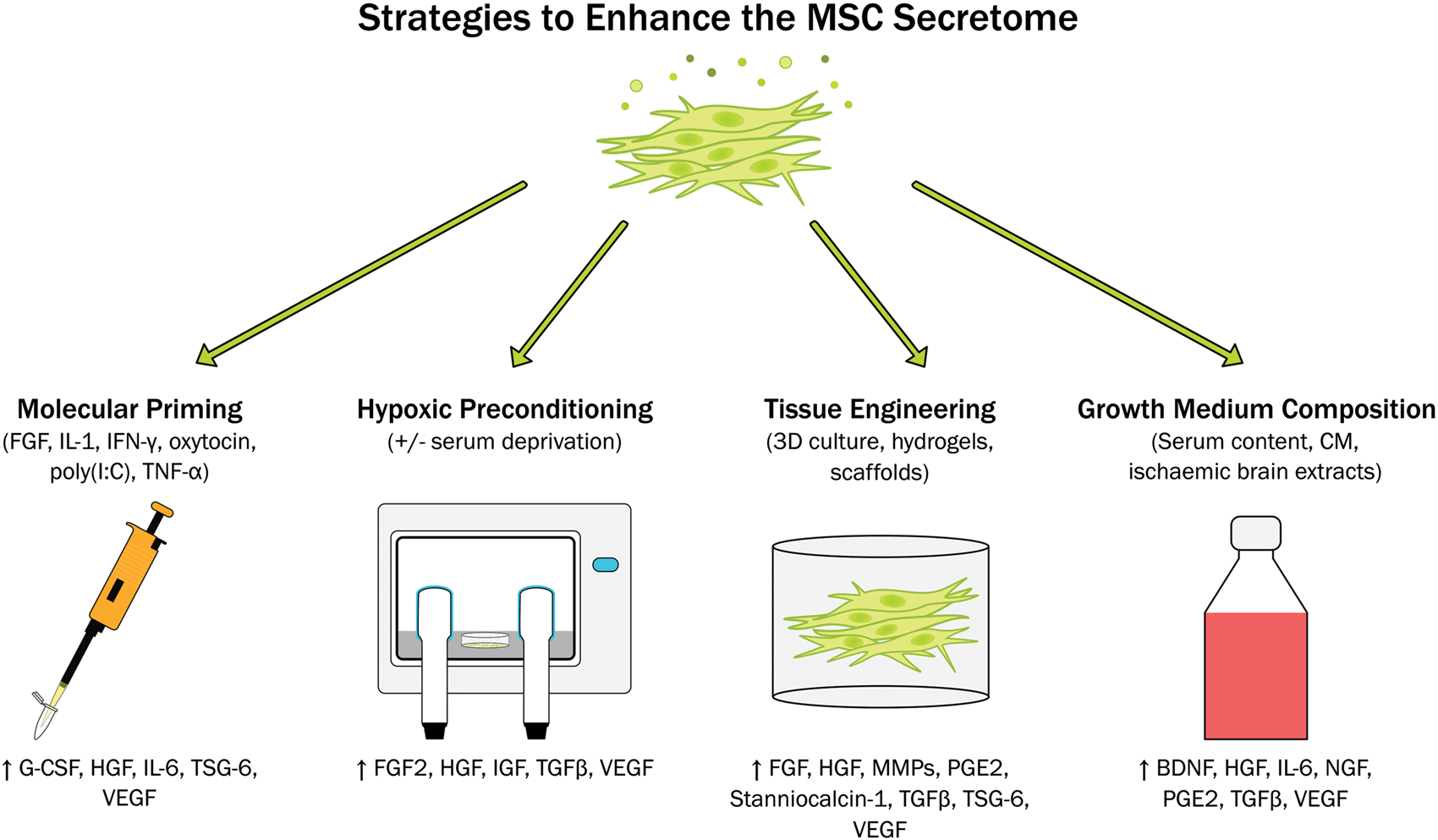
Molecular priming
Priming or preconditioning acts as a sub-lethal event that can trigger an adaptive response to a future injury or damage. Therefore, administration of “trained” cells better able to respond to the ischaemic and inflammatory environment post-stroke may further enhance the efficacy of MSC therapies. MSCs from different sources (mainly bone marrow, adipose, placenta and umbilical cord) and from different species (human, equine, murine) have been preconditioned or primed. Such in vitro preconditioning strategies can be selective and aimed at improving the secretion of certain factors such as anti-inflammatory TNF-α-stimulated gene 6 protein (TSG-6), 41 or to increase survival of MSC once transplanted. 42 Non-selective approaches aim to modulate the MSC secretome towards a more desirable phenotype by inducing the secretion of immunomodulatory, 43 anti-inflammatory 44 or pro-angiogenic molecules. 45
MSCs are known to be great immune modulators, so they are often used to decrease inflammatory responses. To enhance this characteristic, cells can be primed with inflammatory mediators such as IL-1, 43 TNF-α,41,46,47 IFN-γ48,49 or combinations of these. 50 In response to these priming stimuli, MSCs secrete higher concentrations of immunomodulatory mediators including prostaglandin E2 (PGE2), IL-6 and granulocyte-colony stimulating factor (G-CSF)43,51 and upregulate adhesion molecule expression.46,50 This leads to increased promotion of endogenous repair mechanisms including angiogenesis46,50 and osteogenesis 46 which has been shown to be beneficial in in vivo models of arthritis, 48 joint and cartilage injuries47,50 and bone regeneration. 46 It has been reported though, that inflammatory priming can lead to an increased immunogenicity. 50 As this can be detrimental in future cell therapies, short priming durations with low doses of pro-inflammatory mediators should be used to limit this undesirable effect. For example, our lab demonstrated 5 min of priming with IL-1α drove the MSC secretome towards a more anti-inflammatory phenotype which decreased secretion of TNF-α and IL-6 from inflamed mouse microglia. 43 A wide variety of molecules can be used to prime MSCs and modify their secretome. Indeed, the screening of libraries has already become a suitable strategy to detect active molecules. 52 As an example, polyinosinic and polycytidylic acid (poly(I:C)) can be used as a toll-like receptor 3 (TLR3) stimulus to induce an increased anti-inflammatory phenotype, 44 while oxytocin 53 or FGF245 have been used to increase the angiogenic potential of the MSC secretome.
Hypoxia
Another alternative to induce an improved response to ischemic environments is the use of hypoxic or ischaemic preconditioning. This has been shown to induce increased MSC proliferation and migration, 54 upregulation of glucose transporters and adhesion molecule expression, 55 and drive the secretome towards a pro-angiogenic phenotype. 56 More specifically, hypoxic preconditioning of bone marrow-derived MSCs induces increased secretion of FGF2, VEGF, HGF, TGF-β and IGF.57,58 This has also been reported in MSCs derived from other sources including placenta59,60 and adipose tissue. 61 Hypoxic preconditioning can enhance the therapeutic potential of MSCs in vivo preventing apoptosis of cardiomyocytes and promoting angiogenesis after myocardial infarction 62 as well increasing secretion of VEGF, HGF and FGF in a murine model of critical limb ischaemia. 54 Serum deprivation is often used in conjunction with hypoxic preconditioning as it improves the ability of MSCs to induce angiogenesis and endothelial proliferation.63,64
3D culture and biomaterials
The 3D culture of MSCs is another option to achieve a more effective therapy for ischaemic stroke.65–67 Culturing in 3D enhances the angiogenic potential of MSCs by increasing the secretion of molecules including VEGF, HGF and FGF2,65,68,69 and increases anti-inflammatory potential by secreting TSG-6, stanniocalcin-1, PGE2 or TGF-β amongst others.65,70,71 This 3D environment provides more physiological conditions, maintains stemness and increases cell survival and multipotency once transplanted.65,72 Additionally, this increases the ability of MSCs to activate endogenous mechanisms of tissue repair through increased secretion of factors such as matrix metalloproteinases (MMPs) and FGF2.67,73 MSC spheroids have already shown moderate success in promoting bone regeneration74,75 and in inflammatory models such as colitis. 76
In some studies, biomaterials including hydrogels, assembling peptides or scaffolds have been utilised to further enhance the anti-inflammatory and pro-trophic phenotype of the MSC secretome. Murphy et al. 77 showed that entrapping MSC spheroids in a fibril gel can increase secretion of VEGF and PGE2, increase endothelial cell proliferation and promote angiogenesis in a human 3D skin equivalent wound model. Similarly, conditioned medium (CM) derived from MSCs embedded in collagen and polyethylene glycol hydrogels induced stronger antioxidant and neuroprotective responses in SH-SY5Y cells. 78 MSCs cultured with self-assembly peptides induced in vitro outgrowth of axons and neurites from neurons following traumatic brain injury. 79 Combined administration of MSCs and biomaterials has been previously shown to promote repair in a number of disease models. For example, embedding MSCs in platelet lysate hydrogels increased engraftment as well as increasing the pro-angiogenic and neo-vascularisation activity of the transplanted cells in a murine model of critical limb ischaemia. 80
CM and serum preconditioning
When the molecule intended to trigger a particular effect is not known, or when a specific environment needs to be mimicked, CM or serum is another suitable option for modifying the MSC secretome. MSCs treated with endothelial growth medium show improved viability and endothelial-related functions, 81 while priming MSCs with serum from stroke animals increased proliferation and secretion of cytokines, thus improving their therapeutic potential. 82 Similarly, when cultured in rat ischaemic brain extracts, MSCs respond by increasing secretion of BDNF, VEGF, NGF and HGF. 83 The serum content of growth medium can have a profound effect on the MSC secretome. Zimmerman and McDevit 71 showed the secretion of immunomodulatory factors such as PGE2, IL-6 and TGF-β was far increased when MSC spheroids were cultured in growth media containing foetal bovine serum as compared with a specialised MSC serum-free medium.
Role of the MSC secretome in promoting repair in preclinical models of stroke
There is a substantial body of evidence demonstrating MSC transplantation promotes recovery in rodent models of stroke although the mechanisms of action have not been fully elucidated. A number of studies from the early 2000s began to hypothesise that the MSC secretome was involved. Zhao et al. 84 suggested that as intracranial (IC) administration of hMSCs one week after middle cerebral artery occlusion (MCAO) in spontaneously hypertensive (SHR) rats was associated with improvements in limb placement but differentiation was limited, recovery might be mediated through secretion of neurotrophic factors from the transplanted cells. Similarly, IV administration of MSCs also improved neurological deficits and the authors proposed neurotrophins from the MSCs decreased apoptosis and promoted endogenous neurogenesis. 28 Later work from the same lab also showed that MSC transplantation increased angiogenesis in the ischaemic boundary. 85 This was associated with increased endogenous VEGF and VEGF receptor 2 (VEGFR2) expression, which the authors hypothesised were upregulated by secretion of growth factors such as bFGF from the MSCs. In support of this, exogenous IGF-1 from transplanted cells has been detected in the core and ischaemic border zone three days post-MCAO, while expression of endogenous growth factors including VEGF, EGF and bFGF was increased in MSC-treated rats compared with controls. 86 Additionally, secretion of a number of other factors from MSCs engrafted in the ischaemic brain has been detected including BDNF, bFGF, CXCL12, platelet-derived growth factor-AA (PDGF-AA) and angiopoietin-2 (Ang-2).87,88
One neurotrophin of particular interest is BDNF which promotes neuronal survival and differentiation through interaction with tyrosine kinase receptors. 89 In preclinical models of stroke, IV BDNF administration reduced infarct volume, improved recovery and promoted neurogenesis.90,91 Furthermore, BDNF appears to be an important mediator in the MSC secretome preventing glutamate-induced neuronal death in vitro. 92 When transplanted into a stroke model, BDNF secretion from MSCs was associated with increased functional recovery, decreased lesion volume, decreased apoptosis and increased angiogenesis. 34 Several studies have shown that overexpression of BDNF in MSCs further enhanced repair and recovery.93–95 However, Koh et al. 96 demonstrated that neutralising BDNF did not completely ameliorate the observed improvements in neurological function following human umbilical cord-derived MSC transplantation, suggesting other mediators are important in promoting recovery after stroke.
Summary of studies investigating the efficacy of MSC therapies in preclinical models of cerebral ischaemia and the proposed involvement of secretome components.
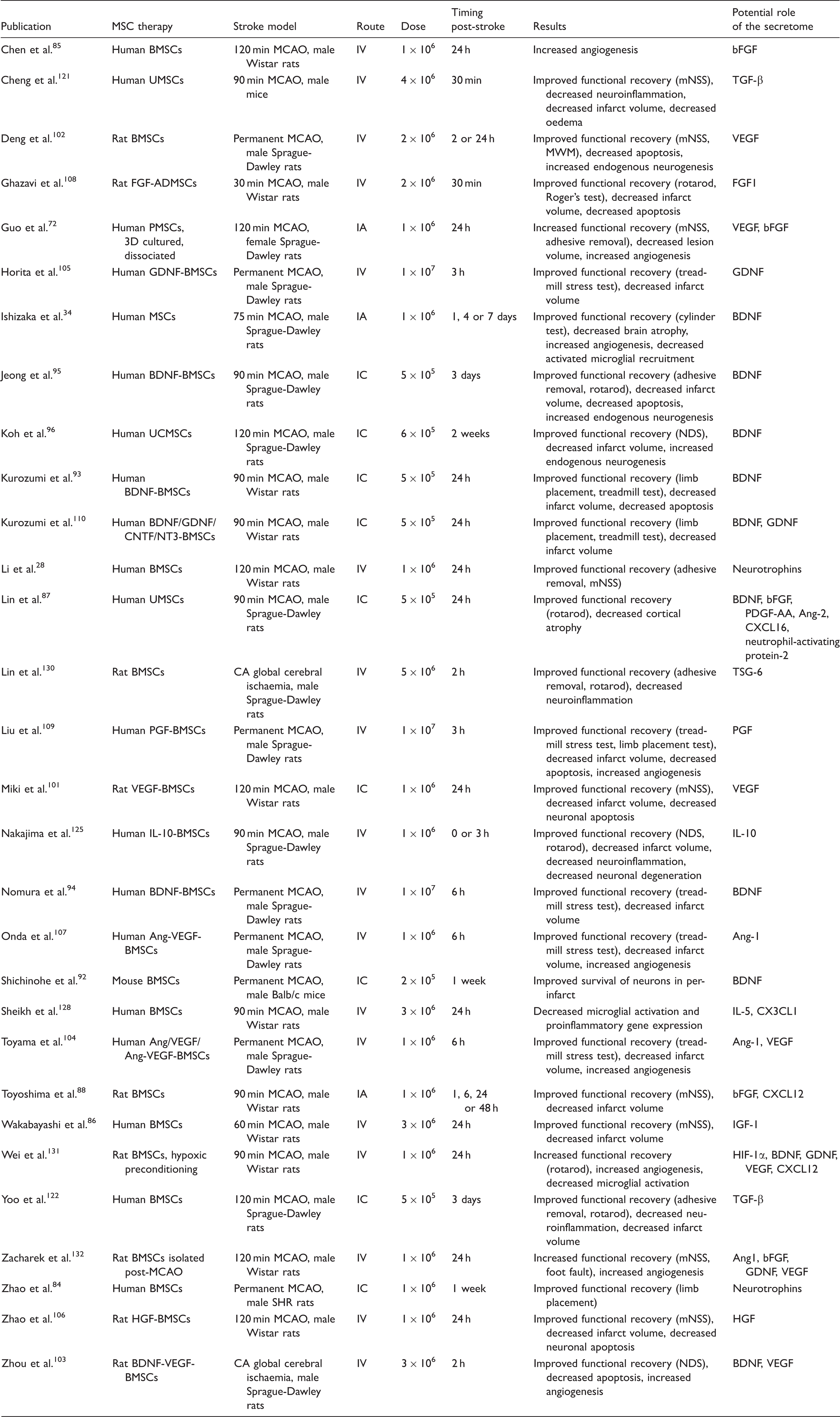
BMSCs: bone marrow-derived mesenchymal stem cells; CA: cardiac arrest; IN: intranasal: MCAO: middle cerebral artery occlusion; MWM: Morris water maze; mNSS: modified neurological severity score; NDS: neurological deficit score; PMSCs: placenta-derived mesenchymal stem cells; UMSCs: umbilical cord-derived mesenchymal stem cells.
Immunomodulation
While the consensus in the literature is that the MSC secretome promotes recovery after stroke through mechanisms including neuroprotection, neurogenesis and angiogenesis after stroke, its role in immunomodulation is not clear. MSCs exert numerous immunomodulatory effects on immune cell populations including inhibition of proliferation of natural killer (NK) cells, 111 inhibition of both B and T cell proliferation112–114 and suppression of dendritic cell (DC) differentiation and migration.115,116 Additionally, co-culture of MSCs drives the secretome of DCs, T cells, macrophages and NK cells towards anti-inflammatory phenotypes.117,118 A number of molecules secreted by MSCs including PGE2, TSG-6, TGF-β, HGF and IL-10 have been implicated in mediating these immunosuppressive effects.119,120 For example, Di Nicola et al. 113 showed TGF-β and HGF secretion was involved in MSC suppression of T-lymphocyte proliferation. 113 Following on from this, TGF-β secretion from transplanted MSCs improved the systemic inflammatory response after stroke decreasing Th17 cells and increasing regulatory T cells in the peripheral immune system. 121 This was associated with decreased infarct volume and improved functional recovery. Furthermore, transplantation of TGF-β silenced MSCs did not decrease CD68+ cell infiltration or prevent microglial cell death as demonstrated in non-modified cells. 122
IL-10, often referred to as an anti-inflammatory cytokine, is an inducer of immune tolerance and has previously been shown to have neuroprotective effects and decrease pro-inflammatory signalling in preclinical models of cerebral ischaemia.123,124 Transplantation of MSCs overexpressing IL-10 led to decreased microglial activation and pro-inflammatory cytokine (IL-6, TNF-α and IL-1β) concentrations in the brain after stroke compared with non-modified MSCs and vehicle. 125 Administration of IL-10-MSCs was also neuroprotective leading to decreased neuronal degeneration and improved functional recovery. CX3CL1 (fractalkine) may also have a role in immunomodulation after cerebral ischaemia. Its receptor CX3CR1 is expressed by microglia and CX3CL1-CX3CR1 signalling supresses neurotoxic microglia activity. 126 Secretion of CXC3CL1 from MSCs has previously been shown to shift microglia towards a neuroprotective phenotype. 127 Sheikh et al. 128 suggested CX3CL1 and IL-5 were involved in decreasing microglial activation and inhibiting expression of pro-inflammatory gene expression, namely COX-2 and iNOS, in the core and ischaemic border zone.
TSG-6 secretion from MSCs has previously been shown to decrease inflammation in peritonitis and corneal injury models.41,129 MSC administration in a cardiac arrest-induced global cerebral ischaemia rat model led to decreased serum pro-inflammatory cytokines and S100B concentrations and decreased expression of neutrophil elastase in the cerebral cortex. 130 While TSG-6 expression in the cerebral cortex was upregulated, it was not possible to determine whether this was due to secretion from the MSCs or endogenous cells.
Secretome modification
As discussed earlier, a number of in vitro strategies have been utilised to enhance the MSC secretome but few have investigated whether these lead to enhanced recovery of function in preclinical models of cerebral ischaemia. Transplantation of hypoxic preconditioned MSCs was superior to normoxic-treated cells leading to larger improvements in functional recovery, increased angiogenesis and decreased microglial activation. 131 The authors proposed this was mediated by enhanced secretion of trophic factors and reported upregulated expression of BDNF, VEGF, GDNF, and CXCL12 in hypoxic cells. Similarly, Zacharek et al. 132 demonstrated that MSCs isolated from rats after MCAO provided a better allogeneic stroke therapy compared with cells from naïve animals and was associated with increased Ang1, bFGF, GDNF and VEGF expression. The 3D culture of MSCs has also been shown to enhance recovery. MSCs cultured as spheroids and then dissociated prior to IA administration led to improved functional outcomes, increased angiogenesis and decreased lesion volume. 72
MSC CM treatment
In further support of the important role of the MSC secretome, CM has also been shown to promote recovery in rodent models of cerebral ischaemia. Egashira et al. 133 reported that adipose-derived hMSC CM administered by intracerebroventricular (ICV) injection 1 h prior to MCAO in inbred DDY mice led to decreased lesion volume and neurological deficits at 24 h post-stroke. Additionally, delayed administration of CM from spheroid cultured cells beginning at day 8 post-stroke led to decreased microglial apoptosis, increased endothelial cell proliferation and improved rotarod performance at day 15. 134 IV 135 and intranasal 136 administration of CM has also been reported to improve recovery.
MSC-derived EVs
Preclinical studies on the effect of MSC-derived exosomes on repair and recovery after ischaemic stroke.
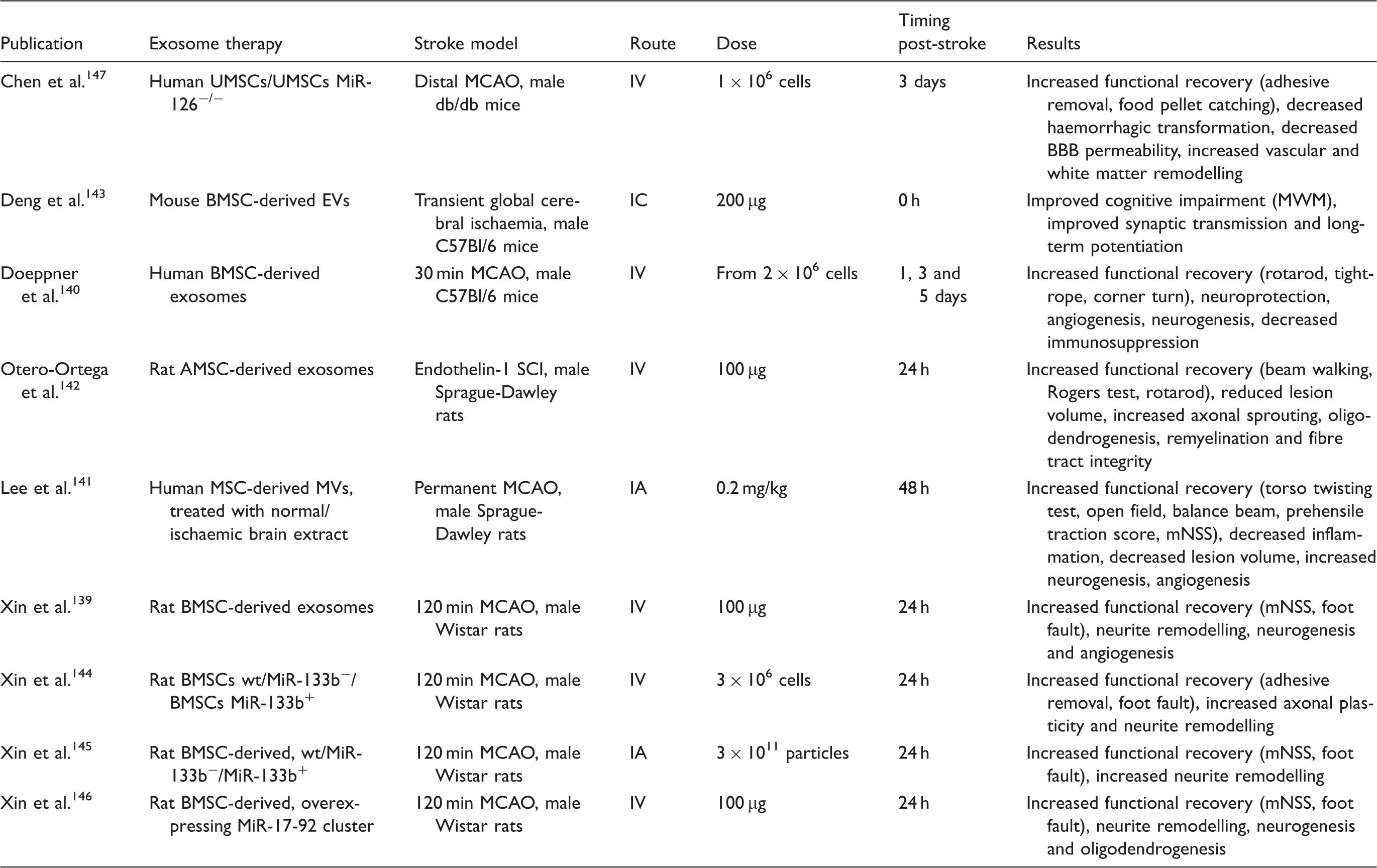
AMSCs: adipose-derived mesenchymal stem cells BMSCs: bone marrow-derived mesenchymal stem cells; EVs: extracellular vesicles; MCAO: middle cerebral artery occlusion; mNSS: modified neurological severity score; MWM: Morris water maze; MVs: microvesicles; SCI: subcortical infarct; UMSCs: umbilical cord-derived mesenchymal stem cells.
Conclusions and future directions
There is a growing body of evidence demonstrating the role of the MSC secretome in promoting recovery in rodent models of cerebral ischaemia. This has been proposed to occur through a number of mechanisms including decreased neuroinflammation, neuroprotection, increased angiogenesis and neurogenesis (Figure 2). However, there is currently no consensus in the literature on what mediators in the MSC secretome are important in promoting repair and functional recovery after stroke. While a strong case can be made for BDNF in particular with multiple citations supporting its role, neutralising BDNF did not completely abolish post-stroke recovery. It is therefore likely that a combination of mediators is important in promoting recovery and repair after stroke. In support of this, meta-analysis has demonstrated that G-CSF does not improve outcomes in stroke patients.
148
A number of in vitro strategies have been used to drive the secretome towards a more desirable anti-inflammatory and pro-trophic phenotype including priming with pro-inflammatory cytokines, hypoxic preconditioning, biomaterials and 3D culture (Figure 1). However, the efficacy of these approaches has not been extensively assessed in preclinical models.
The role of the mesenchymal stem cell secretome in promoting repair and recovery after ischaemic stroke. The main mechanisms of action are highlighted along with the proposed mediators. Ang: angiopoietin; BDNF: brain-derived neurotrophic factor; CXCL: chemokine C-X-C motif ligand; CX3CR1: CX3C chemokine receptor 1; bFGF: basic fibroblast growth factor; GDNF: glial cell line-derived neurotrophic factor; HGF: hepatocyte growth factor; HIF-1α: Hypoxia-inducible factor 1-alpha; IGF-1: insulin-like growth factor 1; IL: interleukin; PDGF-AA: platelet-derived growth factor AA; PGF: placental growth factor; TGF-β: transforming growth factor beta: VEGF: vascular endothelial growth factor.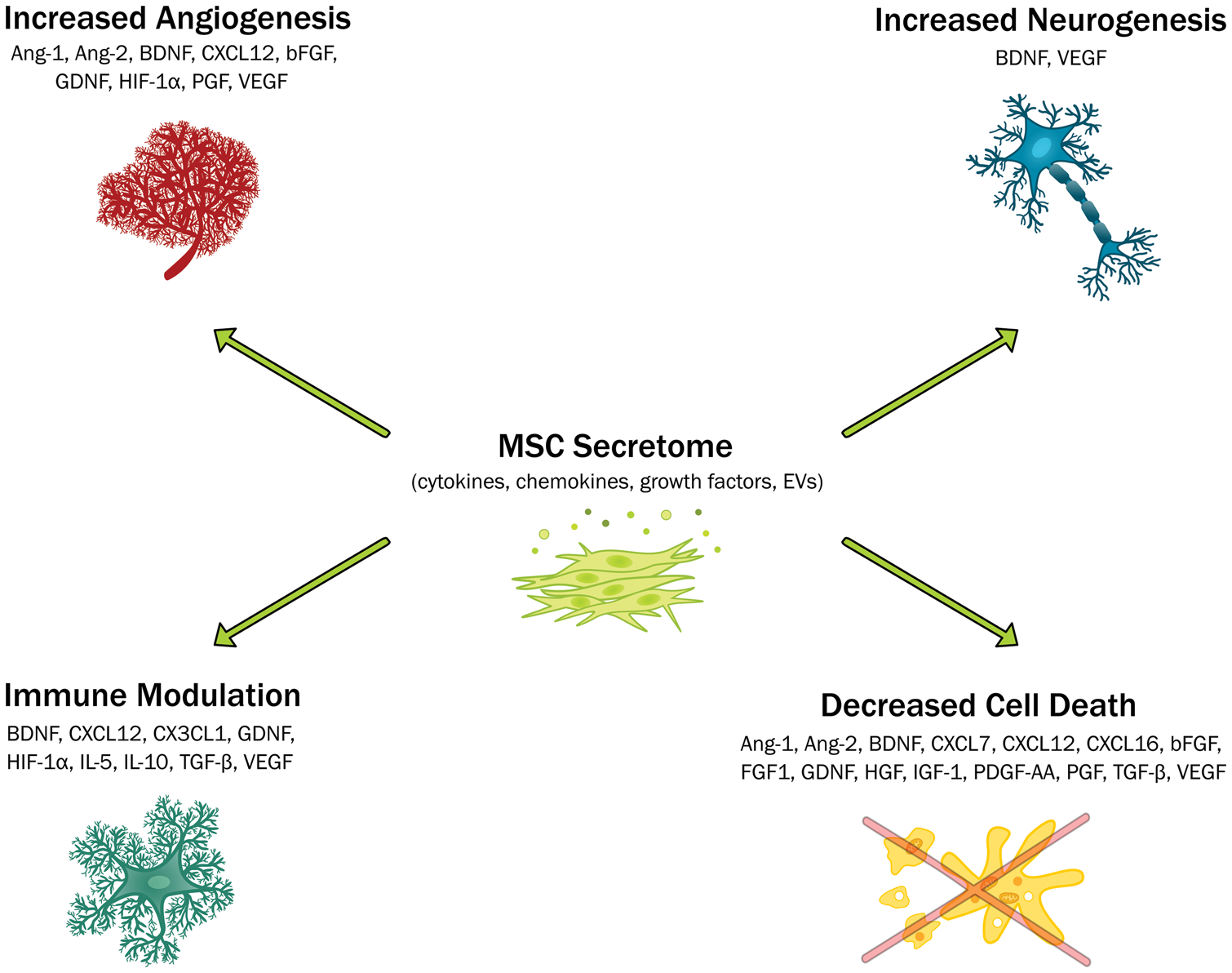
There are several challenges to be overcome in translating the MSC secretome into a safe and effective therapy for ischaemic stroke such as the optimal timing of administration. The majority of preclinical studies elected to administer MSCs, CM and exosomes during the acute phase of stroke (≤ 48 h) where secondary damage is mediated by reactive oxygen species, migration of immune cells to the ischaemic brain and production of pro-inflammatory cytokines such as IL-1. 149 As a number of studies have demonstrated immunomodulatory and neuroprotective effects of the MSC secretome, such a time point may hold therapeutic potential. In contrast, one study reported that administration of MSCs to rats at 1 month post-stroke also led to functional recovery. This was associated with decreased glial scarring and increased proliferating cells in the subventricular zone, suggesting MSC treatment may have promoted neurogenesis. 150 As MSCs secrete multiple growth factors which can activate endogenous repair mechanisms, administration at delayed time points should be investigated further. 151 Determining the optimal timing of administration may prove to be a difficult balancing act and repeated dosing should be considered. For example, VEGF induces vascular permeability so if administered at acute time points may increase BBB breakdown leading to increased cerebral oedema and exacerbate injury. Another challenge will be determining the best therapy. While MSCs are generally immune evasive and have been shown to be well tolerated in clinical trials in stroke, the increasing number of preclinical studies demonstrating the efficacy of MSC-derived CM and EVs could mitigate the need to administer cells. This may prove more translatable as these cell-free alternatives can be cryopreserved without any concerns over cell viability so could be stored for long periods of time and shipped worldwide. Another challenge will be determining the route of administration. Preclinical studies and clinical trials have employed both systemic routes such as IV and IV and direct routes such as IC. As improvements in recovery can occur independently of MSC engraftment or even migration to the ischaemic brain, perhaps systemic routes which are simpler, less invasive and less likely to cause adverse events should be adopted.
Looking forward, the biggest challenge to preclinical scientists is that there is currently no clear consensus on the optimum culture conditions and preconditioning strategy to maximise the regenerative potential of the MSC secretome. Future work should focus on assessing the efficacy of more approaches to modifying secretome in rodent models of cerebral ischaemia and increasing our understanding of the mediators involved in promoting repair. There is growing interest in cell-free approaches such as exosomes or CM and these should also be more fully investigated. In summary, while there are a number of hurdles to overcome on road to translation, the MSC secretome holds much potential as a regenerative therapy for ischaemic stroke.
Footnotes
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by an EPSRC and MRC Centre for Doctoral Training in Regenerative Medicine studentship grant EP/L014904/1.
Acknowledgements
The authors would like to thank Raymond Wong for his valuable feedback on the manuscript.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.