
Abstract
Embryonic mesenchymal stem cells (eMSCs) were first derived from human embryonic stem cells (hESCs) overexpressing green fluorescence protein (GFP). They expressed CD29, CD44, CD73, CD105, CD166 and nestin, but not CD34, CD45, CD106 SSEA-4 or Oct3/4. Twenty million eMSCs in 1mL of phosphate-buffered saline (PBS) were injected into the femoral veins of spontaneously hypertensive rats after transient middle cerebral artery occlusion. The migration and differentiation of the eMSCs in the ischemic brain were analyzed. The results revealed that eMSCs migrated to the infarction region and differentiated into neurons, which were positive for β-tubulin III, microtubule-associated protein 2 (MAP2), HuC, neurofilament and human nuclear antibody, and to vascular endothelial cells, which were positive for von Willebrand factor (vWF). The transplanted cells survived in the infarction region for at least 4 weeks. Adhesive removal function significantly improved in the first week after cell transplantation, and rotarod motor function significantly improved starting from the second week. The infarction volume in the eMSC group was significantly smaller than that in the PBS control group at 4 weeks after infusion. The results of this study show that when administered intravenously, eMSCs differentiated into neuronal and endothelial cells, reduced the infarction volume, and improved behavioral functional outcome significantly in transient focal cerebral ischemia.
Keywords
Introduction
Stroke is the leading cause of disability and the third cause of death in adults worldwide (van Gijn and Dennis, 1998). For a long time, researchers have attempted to replace the damaged neurons in the brain after stroke, but have had only limited clinical success. Recombinant tissue plasminogen activator (rtPA), approved by the FDA in the treatment of ischemic stroke, is beneficial for the recovery of blood flow with no effect on the damaged neuronal tissue or vascular endothelium. Recent studies on stem cell therapy in animal models of stroke have revealed promising results (Bliss et al, 2007; Chen et al, 2001a, b ; Shen et al, 2007b). Most of these studies used mesenchymal stem cells (MSCs) derived from bone marrow or umbilical cord blood.
Mesenchymal stem cells either from bone marrow or from umbilical cord blood have the potential to differentiate into many types of cells, including endothelial cells (Du and Taylor, 2007) and neuron-like cells (Kan et al, 2007; Kim et al, 2006; Tropel et al, 2006; Yang et al, 2006).
Despite the benefits of MSC therapy in animal models of cerebral ischemia, their sources are limited and extraction requires invasive procedures. Although the
Human embryonic stem cells (hESCs) have the potential to differentiate into any kind of cell in the body, and, theoretically, possess limitless expansion and pluripotent capacity. The derivation and long-term maintenance of hESCs
Materials and methods
Human Embryonic Stem cell Culture and Mesenchymal Stem Cell Differentiation
The H1 ESCs with stable overexpression of GFP were established previously and named H1EGFP (Liu et al, 2004). The cells were grown as described previously (Liu et al, 2004). Mesenchymal stem cell differentiation was performed using the methods described by Lian et al (2007) with some modifications. A confluent six-well plate of H1EGFP was trypsinized for 3mins at 37°C, neutralized, centrifuged, and resuspended in knockout Dulbecco's modified Eagle's medium (DMEM) (Gibco, Grand Island, NY, USA), supplemented with 10% knockout serum replacement medium (Gibco), and 5ng/mL basic fibroblast growth factor (bFGF) (Gibco) on a gelatinized 10-cm plate. The cells were trypsinized when they were confluent and sorted for CD73 (#550257; BD PharMingen, San Diego, CA, USA) positivity 1 week after the initial passage. Positive cells were cultured in MSC media (DMEM supplemented with 50 µg/mL gentamycin, 0.5mmol/L glutamine, 1% nonessential amino acids, 5ng/mL bFGF, and 10% fetal bovine serum (Invitrogen-Gibco, Carlsbad, CA, USA)) for a maximum of 10 passages.
Flow Cytometry Analysis of the Mesenchymal Stem Cells
Cells were dissociated with trypsin (0.25%) plus EDTA (1 mmol/L; Gibco), washed twice in PBS containing 0.1% bovine serum albumin, and suspended in a concentration of 6 × 104 cells in 0.1mL PBS. Five microliters of phycoerythrin (PE)-conjugated antibody was added to the cells, incubated for 30 mins at room temperature in the dark, washed twice in 0.1% bovine serum albumin/PBS, and immediately analyzed on a FACSCalibur flow cytometer (BD Biosciences, San Jose, CA, USA) with FlowJo acquisition and analysis software (Tree Star Inc., San Carlos, CA, USA). The antibodies used for detection of cell surface markers by FACS included CD34-PE (#550619), CD73-PE (#550257), CD106-PE (#555647), CD166-PE (#559263), and SSEA-4 (#560128) from BD Biosciences; and CD29-PE (#10-0297), CD44-PE (#16-0441), CD45-PE (#16-0459), CD54-PE (#16-0549), and CD105-PE (#12-1057) from eBiosciences (San Diego, CA, USA).
Transient Focal Cerebral Ischemia Model
Spontaneously hypertensive rats ((290 to 310 g, male, Charles River, Wilmington, MA, USA) were housed and cared for in accordance with the
Stem Cell Transplantation
Twenty-four hours after reperfusion, animals were anesthetized with 3.5% isoflurane and were maintained with 1.0% to 2.0% isoflurane in 70% N2O and 30% O2 with the use of a face mask. Approximately 20 × 106 eMSCs in 1.0mL of PBS were injected into the femoral vein. Control rats were injected with 1.0mL of PBS. Both groups of animals were intraperitoneally injected with cyclosporine A at a dose of 10 mg/kg body weight daily, beginning 1 day before cell or PBS injections until the animals were killed.
A total of 24 spontaneously hypertensive rats were used in this study. Twenty-one rats received MCAO and three received sham surgery. After 24 h of MCAO, 15 ischemic rats and all three sham-operated rats were injected with 20 × 106 eMSCs in 1.0mL of PBS through the femoral vein. The remaining six rats with MCAO, which received 1.0mL of PBS, formed the control group. The rats in the MCAO group that received eMSC injection were killed at 48 h (
Immunohistochemistry
Animals were fixed by intracardiac perfusion with 4% paraformaldehyde. Brains were postfixed in paraformaldehyde overnight at 4°C, cryoprotected with 30% sucrose/PBS, and were cut in 30-µm-thick coronal sections on a vibratome. Sections were washed with Tris A buffer (0.1 mol/L pH 7.6 Tris, 0.1% Triton X-100) and Tris B buffer (0.1 mol/L pH 7.6 Tris, 0.1% Triton X-100, 0.05% bovine serum albumin) three times for 5 mins each, and incubated with 10% goat serum in Tris B buffer for 1 h. Then mouse or chicken anti-GFP monoclonal antibody in Tris B was added, and sections were incubated at room temperature for 1 h. After washing the sections with Tris A and Tris B, they were incubated in second primary antibodies in 3% goat serum/Tris B overnight at 4°C. After washing with Tris A and Tris B, secondary fluorescence-conjugated antibodies were added and incubated for 1 h at room temperature, washed in PBS and coverslipped. The following primary antibodies were used: mouse anti-GFP (#A-11120, 2 µg/mL, Invitrogen), chicken anti-GFP (#A-ab13970, 1:800, Abcam), mouse anti-Oct4 (C10: sc 5279, 2 µg/mL, Santa Cruz Biotechnology, Santa Cruz, CA, USA), mouse anti-human nuclear (#Mab 1281, 1:80, Chemicon, Billerica, MA, USA), rabbit anti-glial fibrillary acidic protein (GFAP, 1:2000, Dako, Carpinteria, CA, USA), rabbit anti-microtubule-associated protein 2 (MAP2, #AB5622, 1:300, Chemicon), rabbit anti-human Nestin (#AB5922, 1:200, Chemicon), rabbit anti-β-tubulin III (#T2200, 350 ng/mL, Sigma, St Louis, MO, USA), rabbit anti-neurofilament (#AB1981, 1:200, Chemicon), rabbit anti-HuC (#AB5829, 1:200, Chemicon), or rabbit anti-von Willebrand factor (vWF #F3520, 1:500, Sigma). The secondary antibodies used were goat rabbit-IgG-Alexa Fluor 594 (#A11037, 1:400, Invitrogen) anti-mouse IgG-Alexa Fluor 488 (#A11029 1:400, Invitrogen), goat anti-mouse IgG-Alexa Fluor-635 (#A21052, 1:400, Invitrogen), and goat anti-chicken IgY-FITC (#ab7114, 1:500, Abcam). Negative controls were generated by adding the same concentration of primary mouse IgG (#I5381, Sigma), rabbit IgG (#I5006, Sigma), or chicken IgY polyclonal isotype control (#ab54498, Abcam). Images were then acquired on a Bio-Rad MRC 1024 inverted confocal microscope (Bio-Rad, Hercules, CA, USA) using sequential acquisition to eliminate ‘bleed-through’ fluorescence.
Behavioral Tests
In all animals, behavioral tests were performed on 3 consecutive days before MCAO and at 2, 4, 6, 8, 10, 12, 14, 16, 21, and 28 days after MCAO by an investigator who was masked to the experimental groups. The battery of tests consisted of the rotarod test to assess motor function and adhesive removal test to assess somatosensory function.
In the rotarod motor test (Hamm et al, 1994), the rats were first habituated to a stationary rod. After habituation, they were exposed to a rotating rod. Rotation of the rod was increased from 4 to 16 r.p.m., with the animals remaining in the cylinder for at least 300 secs. The animals were trained for 3 days before MCAO. The mean duration (in seconds) on the device was recorded for three rotarod test measurements 1 day before surgery. Motor test data are presented as percentage of mean duration (three trials) on the rotarod compared with the internal baseline control (before surgery).
In adhesive removal test (Schallert et al, 1997), two small pieces of adhesive-backed paper dots (1.9cm in diameter) were used as bilateral tactile stimuli occupying the distal–radial region on the wrist of each forelimb. The rat was then returned to its home cage. The time taken to remove each stimulus was recorded during three trials per day. Individual trials were separated by at least 5 mins. Before surgery, the animals were trained for 3 days. Once the rats were able to remove the dots within 12 secs, they were subjected to MCAO.
Measurement of Infarct Volume
The lesion size was estimated as a percentage of the whole brain according to an earlier report (Kim et al, 2007) using the following formula: [(area of contralateral hemisphere)– (area of remaining ipsilateral hemisphere)] × (100/2)/(area of contralateral hemisphere). The areas of both hemispheres were measured on eight serial coronal sections per brain (2mm apart) and stained with Cresyl Violet, and the area of the infarction was averaged over eight levels.
Quantification of Transplanted Cells
Cell quantification was performed according to the method described earlier with some modifications (Yang et al, 2008). Quantification of transplanted cells was performed in areas covering the transplant, which were captured under a × 20 objective through a fluorescent scope. Cell counts of GFP-positive cells in the transplant were performed on every 10th section using the MetaMorph software (Universal Imaging, Downingtown, PA, USA). The transplant borders were outlined based on the GFP staining. The total number of transplanted cells was estimated by taking into account the transplant area, the thickness of the brain section (30µm), the frequency of sections selected for analysis, and the number of sections obtained from the brain. Total cell number is approximately the sum of the cell counts from serial sections × the thickness of the section (30 µm) × 10. A rat brain usually produces 300 sections in our experiments. For comparison of cell survival at the different time points, the brain sections at the same position were counted, and the same formula was used to sum the number of cells at all time points. For each time point, three brain sections were counted.
Statistical Analysis
Data are presented as mean±s.d. Differences among groups were assessed by analysis of variance with Scheffe's
Results
Characterization of the Mesenchymal Stem Cells Derived from H1EGFP
H1EGFP cells were differentiated into MSC phenotypelike cells using the previously described method (Lian et al, 2007). We used anti-CD73 antibody to sort the CD73 + cells after H1EGFP trypsinization and culture in differentiation media for 1 week. CD73+ cells were expanded in the MSC media for 3 days. The cells were analyzed with a set of cell surface markers (Figure 1). The differentiated cells expressed a similar pattern of cell surface markers as BM-MSC, such as positivity for CD29, CD44, CD73, CD105, and CD166, and negativity for CD34, CD45, CD54, CD106, and SSEA-4. As the morphology and the cell surface marker expression were the same as those reported earlier (Lian et al, 2007), lineage-specific, such as adipogenic, chondrogenic, and osteogenic, differentiation experiments were not repeated. The differentiated cells were defined as eMSCs. The eMSCs were fibroblast-like in morphology and expressed GFP. Although they did not express Oct 3/4, they expressed nestin and β-tubulin III as shown in Figure 2. Embryonic mesenchymal stem cells did not express MAP2 or neurofilament in the culture (data not shown).
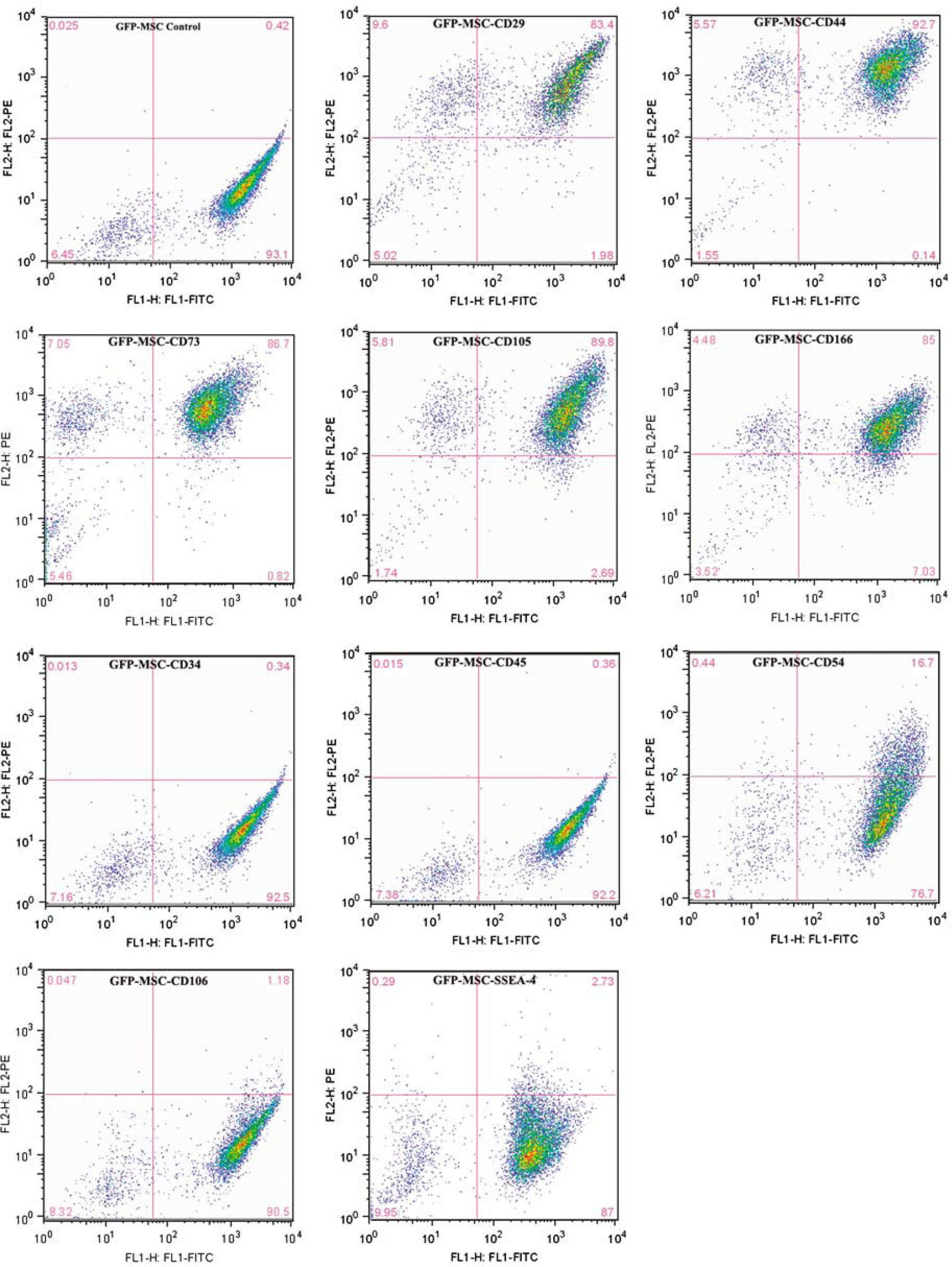
Flow cytometry analysis of cell surface markers in embryonic mesenchymal stem cells (eMSCs). Embryonic mesenchymal stem cells expressed CD29, CD44, CD73, CD105 and CD166, but did not express CD34, CD45, CD106 or SSEA-4. Approximately 10% of the cells expressed CD54. The cell surface markers expression pattern corresponds with that of BM-MSC.
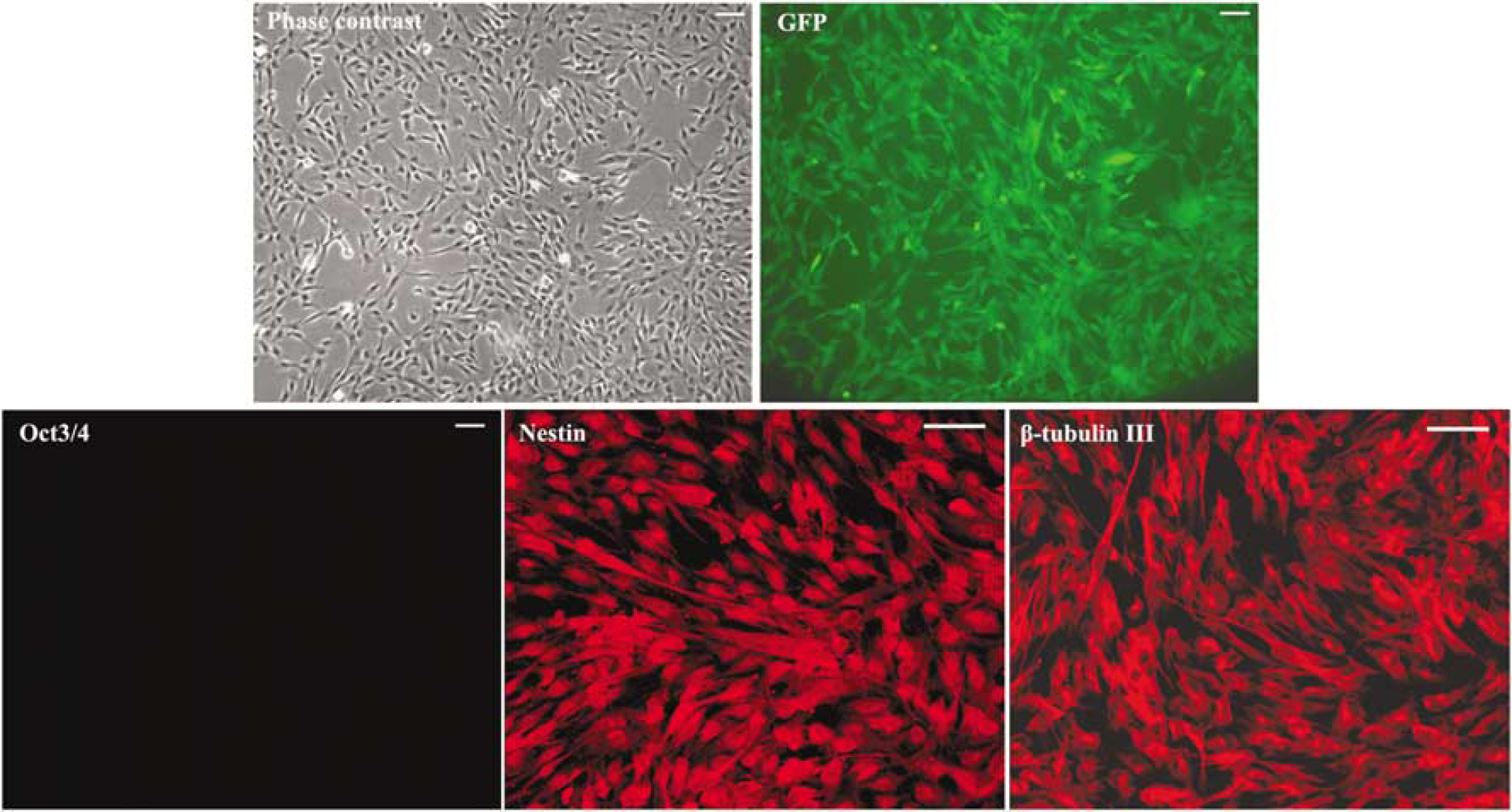
Morphology and immunocytochemistry of embryonic mesenchymal stem cells (eMSCs). Embryonic mesenchymal stem cells look like fibroblasts in morphology with stable GFP expression. They did not express Oct3/4, but expressed nestin and β-tubulin III, and a few cells look like neurons in morphology (bar=100 µm).
Migration of Transplanted Embryonic Mesenchymal Stem Cells into the Infarcted Hemisphere
To investigate the migration of the transplanted cells into the infarction area in the ischemic brain, three rats with MCAO that received eMSC were killed at 2 days after cell injection. The brain sections were stained with GFP antibody. The results showed that there were many GFP-positive cells located in the infarcted hemisphere, not only in the infarction region but also in the penumbra and striatum, whereas no GFP-positive cells were found in the contralateral hemisphere (Figure 3). There was no GFP positivity in the infarction or penumbra regions of PBS-treated rats (Figure 3). Moreover, there were no GFP-positive cells observed in the hemispheres of the sham surgery rats or PBS control rats (data not shown). As we focused on the neuroprotective effects of eMSC transplantation, distribution of the transplanted cells in other organs was not screened.
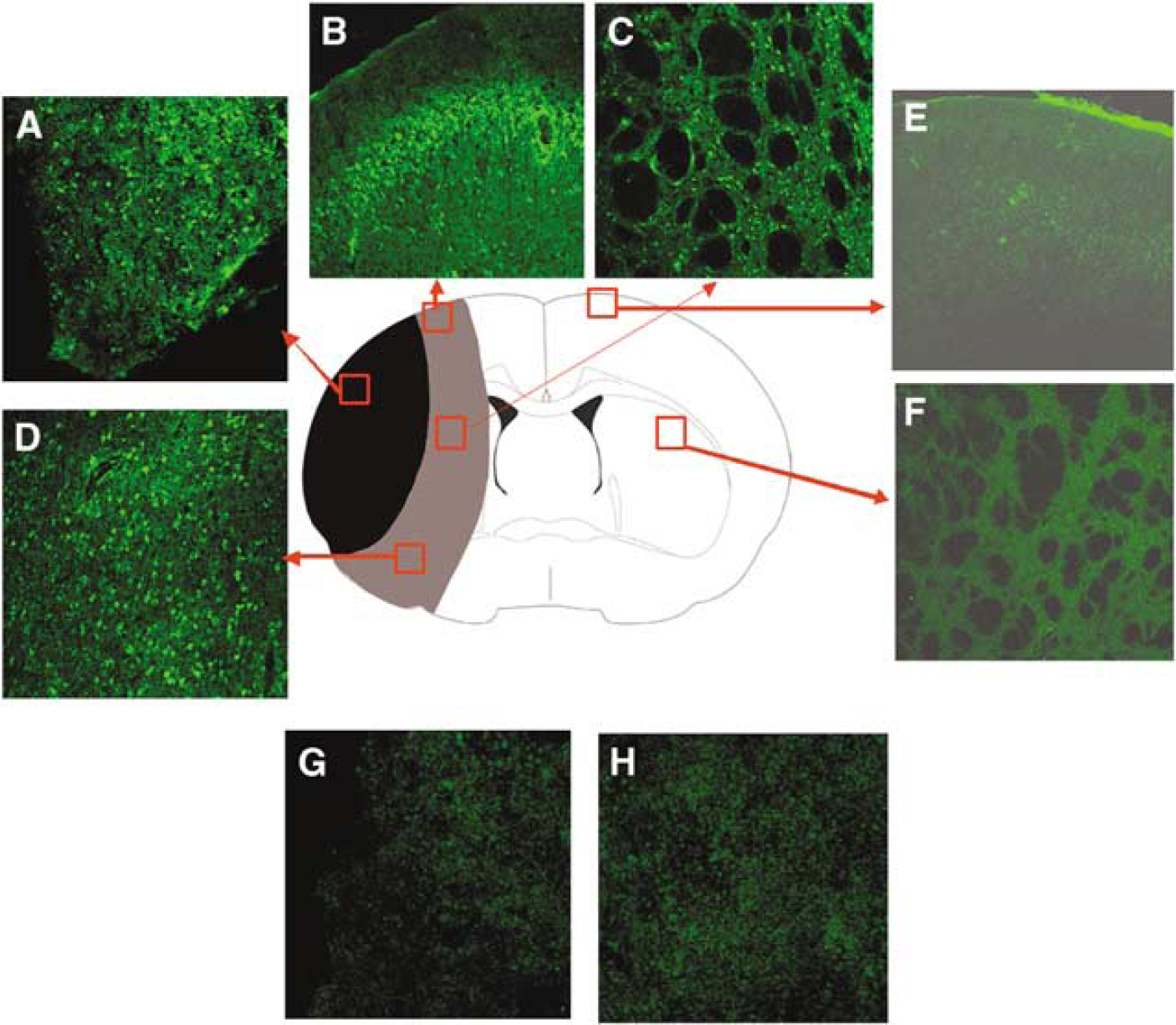
Migration of embryonic mesenchymal stem cells (eMSCs). At day 2 after eMSCs were injected to the MCAO rats through the femoral vein, eMSCs migrated to the infarcted hemisphere, distributed to a large area in the hemisphere. They were found not only in the infarction core (
Differentiation of Embryonic Mesenchymal Stem Cells in the Ischemic Brain
To examine the phenotypic differentiation of transplanted eMSCs in the ischemic brain, brain sections were stained with anti-GFP antibody and antibodies against different cell markers including neuronal, astroglial and endothelial markers. At 2 days after injection, some of the transplanted cells expressed several neuronal markers, such as β-tubulin III, MAP2, and neurofilament, in the ischemic brain (Supplementary Figure 1), indicating the neuronal differentiation of transplanted eMSCs. Z-stack confirmation was also performed to confirm colocalization (Supplementary Figure 1). Because there were many GFP-positive cells that did not express neurofilament, cross-reaction confirmation was not performed in neurofilament staining. There was no cross-reaction between vWF antibody and GFP or GFP antibody and vWF protein (Supplementary Figure 2). There was no cross-reaction between HuC antibody and GFP protein or GFP antibody and HuC protein (Supplementary Figure 3).
Further confirmation was done with triple staining. Brain sections were stained with mouse anti-human nuclear antibody, chicken anti-GFP antibody, and rabbit anti-neural marker. The secondary antibodies used were goat anti-mouse IgG-Alexa Fluor-635, goat anti-chicken IgY-FITC, and goat anti-rabbit-IgG-Alexa Fluor 594. The results showed that all GFP-positive cells were also human nuclear positive. Although some human nuclear-positive cells showed positivity for GFP and neural markers, such as β-tubulin III, MAP2, and neurofilament (Figure 4), a few human nuclear-positive cells were GFP negative. This might be caused by loss of GFP, because even before the injection, a few MCSs were not GFP positive (Figure 4). Comparison of sections stained at 2 days and 1 week after transplantation with those stained later showed the number of GFP-positive cells in the ischemic brain to decrease significantly at 4 weeks after transplantation, suggesting the loss of transplanted cells. The number of cells significantly decreased at 4 weeks compared with those at 2 days and 1 week after transplantation (**
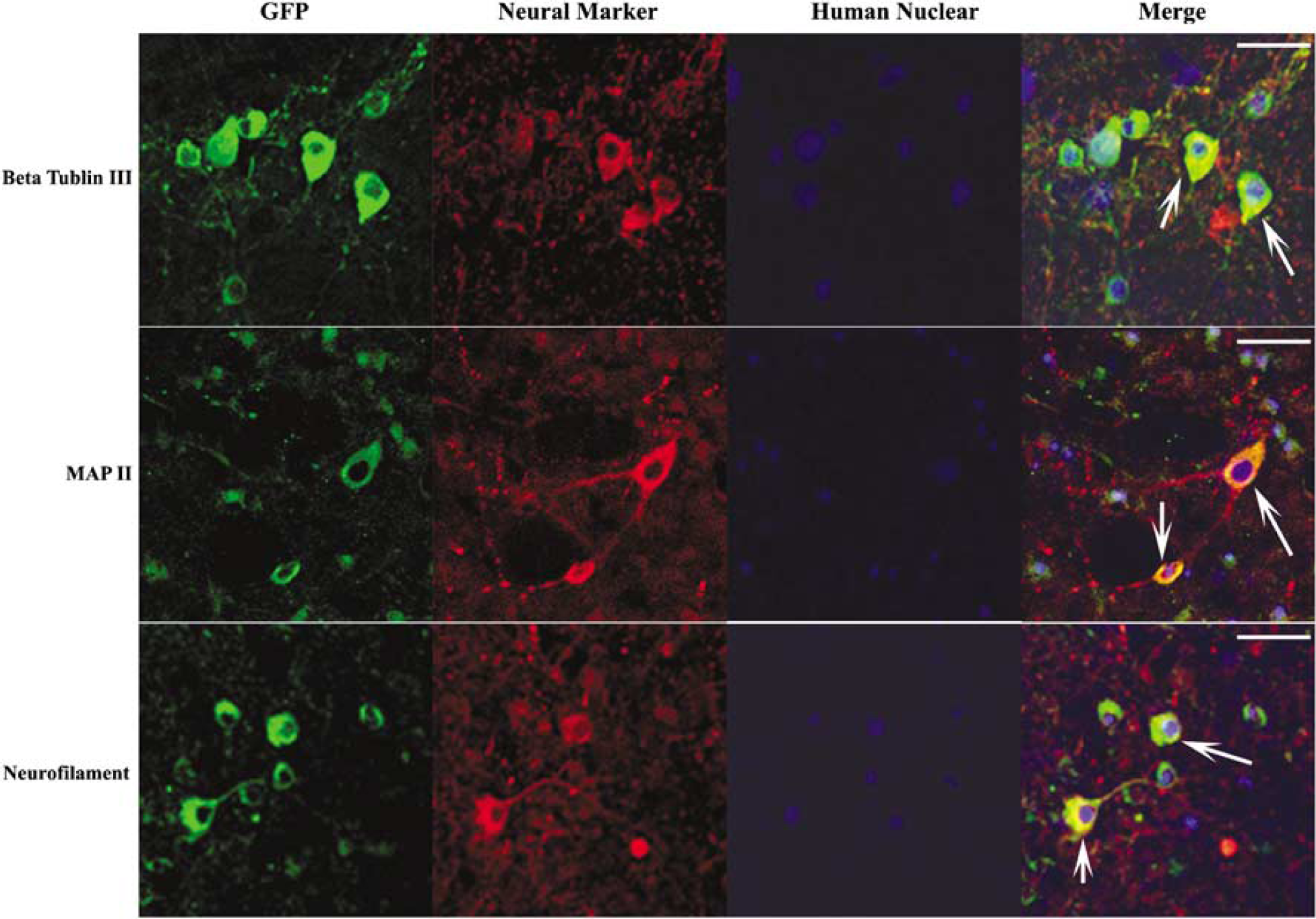
Triple staining with anti-human nuclear antibody. Brian sections were stained with mouse anti-human nuclear antibody, chicken anti-GFP antibody, rabbit anti-neural marker, β-tubulin III, MAP2, and neurofilament. Most human nuclear positive cells were also GFP positive. Some of these cells expressed β-tubulin III, MAP2, and neurofilament (bar=100 µm).
The surviving GFP-positive cells in the ischemic brain still expressed neuronal markers, including β-tubulin III, MAP2, neurofilament, and HuC (Figure 5). The majority of nestin-positive cells were located in the infarction core, whereas the majority of β-tubulin III-positive cells were located in the border of the infarct core and penumbra. The MAP2, neurofilament, or HuC-positive cells were located in the penumbra or healthy area.
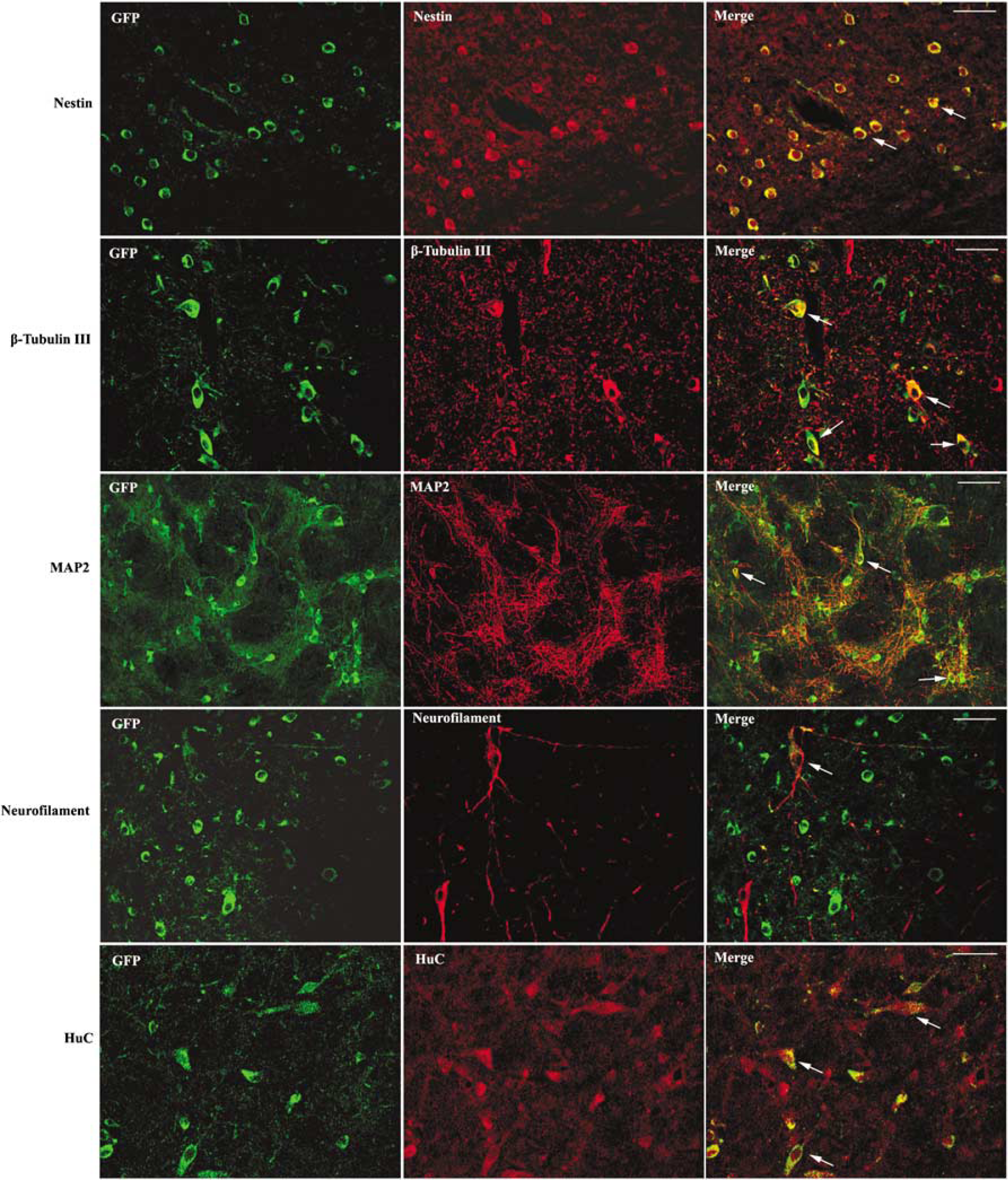
Differentiation of embryonic mesenchymal stem cells (eMSCs) in the infarction region. Embryonic mesenchymal stem cells express nestin, β-tubulin III, MAP2, neurofilament, and HuC in the infarction region. The majority of nestin-positive cells are located in the infarction core. The majority of β-tubulin III is located in the border of infarct core and the penumbra, whereas the MAP2-, neurofilament-, or HuC-positive cells are located in the penumbra or healthy area. All white arrows indicate the colocalization of two antibodies. They indicate the colocalization of GFP and nestin in the top panel, GFP and β-tubulin III in the second panel, GFP and MAP2 in the third, GFP and neurofilament in the fourth, and GFP and HuC in the bottom panel (bar=100 µm).
The transplanted cells did not differentiate into vWF-positive cells in the ischemic brain until 1 week after transplantation (Figure 6). These endothelial cells formed vascular structures within 2 weeks after transplantation (Figure 6). Astroglial differentiation of transplanted cells was not observed at 2 days (data not shown), 2 weeks (data not shown), or 4 weeks (Supplementary Figure 4) after transplantation.
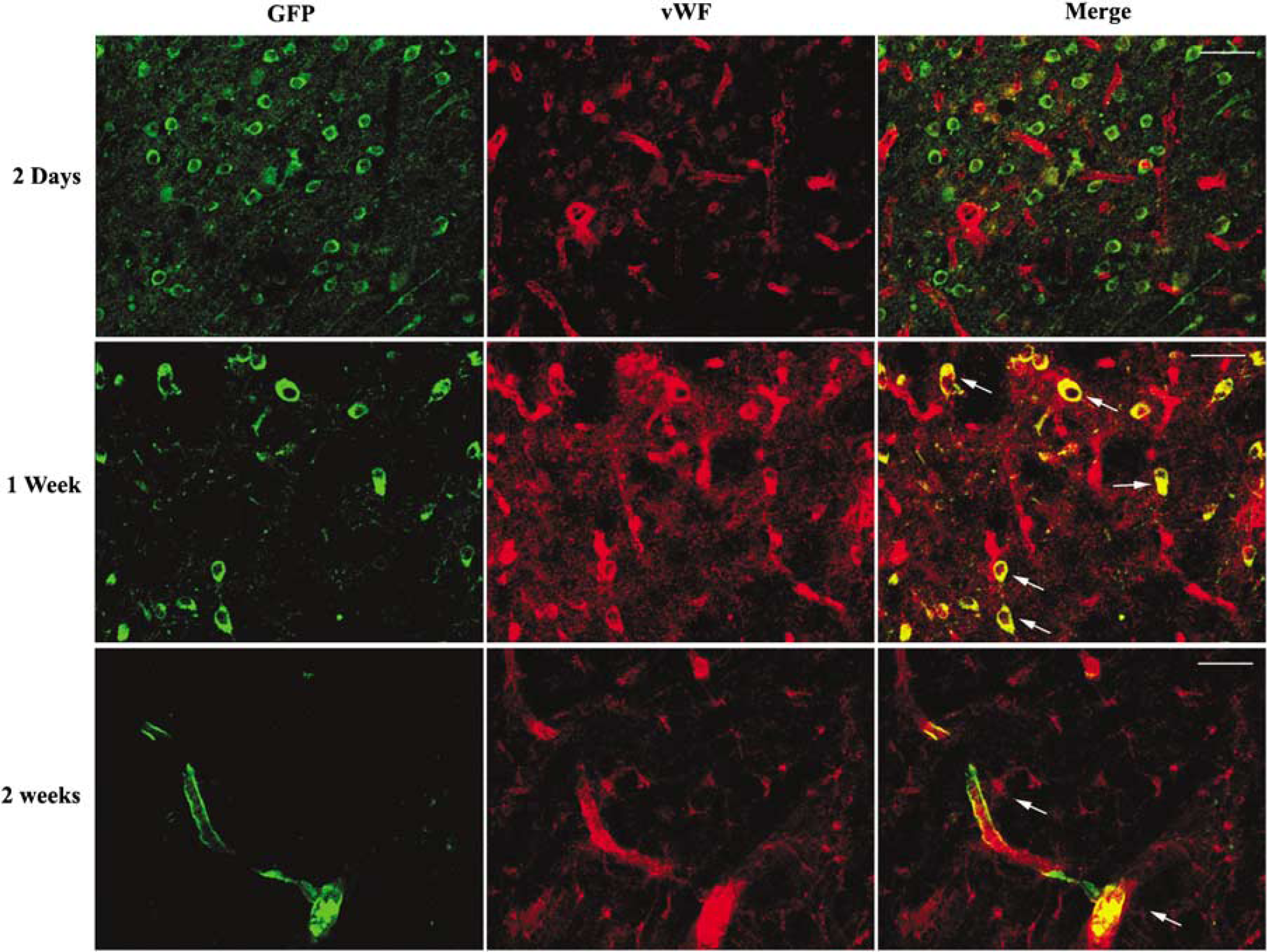
von Willebrand factor (vWF)-positive cells differentiation from eMSC. vWF positivity was not detected at day 2 after cell transplantation. At 1 week, there were many vWF-positive cells. At 2 weeks, vWF- and GFP-positive cells were observed to form vascular channels. White arrows indicate the colocalization of GFP and vWF at 1 week and 2 weeks after transplantation (bar=100 µm).
Effects of Embryonic Mesenchymal Stem Cell Transplantation on Functional Recovery
To evaluate the effect of eMSC transplantation on the recovery of functional deficits in rats subjected to MCAO, behavioral tests were performed. In the adhesive removal test, the mean time taken to remove adhesive from the right forepaw contralateral to the ischemic brain increased at days 2, 4, and 6 after transient MCAO compared with the baseline time in groups that received PBS injection and eMSC injection. Then, the mean time decreased at day 8 after ischemia, but was still higher than the baseline. The mean removal times at days 2, 4, and 6 in the eMSC injection group were significantly less than in the PBS control group (
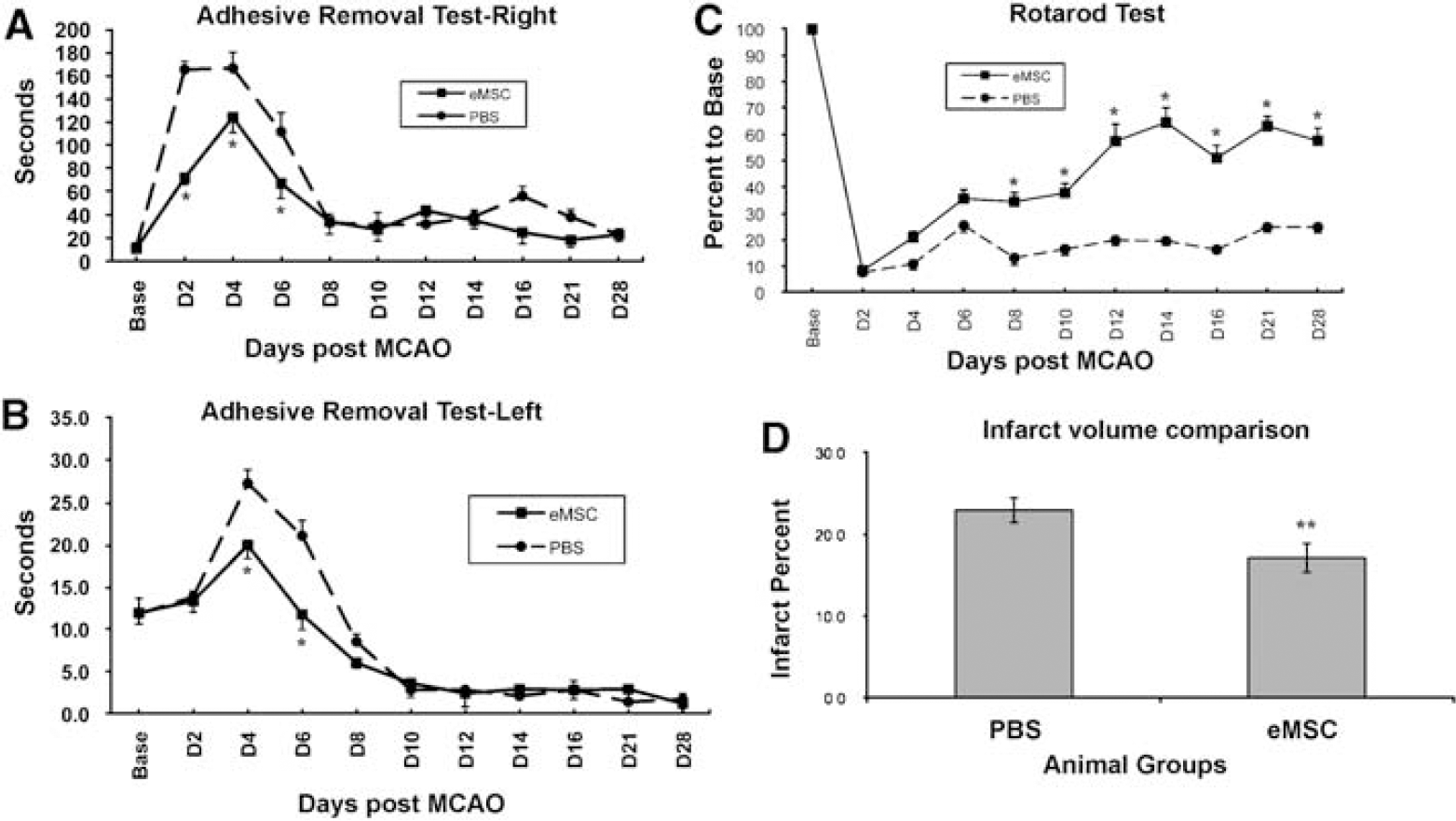
Behavioral test and infarction volume analysis. (
In the rotarod test, the time taken to stay on the rotating rod decreased at day 2 after MCAO, compared with the baseline time before ischemia in the rats that received either PBS or eMSC injection (Figure 7C). There was no significant recovery of functional deficit in the rotarod test from day 6 to day 28 after ischemia in the control group (Figure 7C). In the ischemic rats that received eMSC injection, gradual recovery of functional deficit in the rotarod test was observed starting from day 4 until day 28 after ischemia. The remaining time of the eMSC injection group was significantly longer than that of the PBS control group from day 8 through day 28 (
Infarction Volume
To evaluate the effect of eMSC transplantation on ischemic brain injury, infarction volumes were calculated in the rats 4 weeks after PBS or eMSC injection. The infarction volumes of the rats injected with eMSCs were significantly smaller than those of the rats injected with PBS (
Discussion
Earlier reports have shown successful transplantation of various types of stem cells, such as BM-MSCs (Chen et al, 2001a, b , 2003) and human umbilical cord blood stem cells (Chen et al, 2001c; Chien et al, 2006), to ischemic areas of the animal brain. Despite these promising results, there are some disadvantages of using BM-MSCs. First, the sources of adult MSCs are limited, depending mainly on volunteers. The age, sex, and drug usage of cell donors greatly influence the quality of MSCs. Adult MSCs are highly heterogeneous, not only in their immunogenicity but also in their differentiation and proliferation potential. Moreover, it is necessary to use invasive methods to harvest adult MSCs. In this study, MSC-like phenotype cells derived from hESCs were investigated in a rat model of focal cerebral ischemia. Mesenchymal stem cells derived from hESCs do not have the limitations of adult MSCs. Moreover, because of the success in reprogramming differentiated human somatic cells into pluripotent stem cells (iPS; Lowry et al, 2008; Nakagawa et al, 2008; Park et al, 2008; Takahashi et al, 2007; Yu et al, 2007), this strategy would allow the creation of patient-specific stem cells. It could be greatly beneficial in avoiding immune rejection against the engrafted MSCs in the host if iPS cell-derived MSCs were shown to be useful in stroke.
Various methods have been described for stem cell transplantation (Bliss et al, 2007). In this study, intravenous injection was chosen because an intracerebral injection, into the cortex, striatum or lateral ventricle, is invasive, traverses the normal brain parenchyma in a blind manner and can be potentially complicated by intracerebral hemorrhage or damage. Thus, intracerebral injection may not represent a practical and safe way of delivering stem cells in the clinical setting. Using the intravenous route, the transplanted eMSCs reached the infarction area at 2 days after injection. This result revealed that eMSCs have the same migration potential as BM-MSCs (Caplan and Dennis, 2006; Chen et al, 2003).
In addition to the route of transplantation, the quantity of transplanted cells injected may influence the outcome of ischemic cells. Theoretically, the more cells located in the damaged region, the greater was the potential for replacement or repair of the damaged cells. In this study, 20 × 106 cells were chosen for each animal. In a pilot study before this experiment, various quantities of cells ranging from 1 to 60 × 106 were injected. We observed that the animals were apt to die when they received greater than 30 × 106 cells. With cell counts less than 3 × 106, transplanted cells were hardly observed 2 weeks after the injection. We found the optimal range of the number of cells for transplantation to be 10 to 20 × 106.
Two days after the injection, there were numerous eMSCs located in the infarction area. Earlier reports revealed that BM-MSCs did not differentiate into neuron-like cells until the 10th day in
Behavioral functions in ischemic rats were significantly improved by transplantation of eMSCs. In the adhesive test, although there were no transplanted cells located in the right hemisphere, the adhesive removal function of the left forelimb significantly improved by the first week compared with the control group. This might be because of supportive factors secreted by the transplanted cells. The sensorimotor function was even better than that shown before surgery. It is possible that the animals may have a complementary regulatory mechanism in the right hemisphere when the left hemisphere is injured. The rotarod function was significantly improved beginning from the first week through the fourth week. Although this might be due to the survival and differentiation of the engrafted cells, other mechanisms such as release of growth factors cannot be ruled out.
Four weeks after injection, the number of GFP-positive cells decreased dramatically. A recent report by Yang et al (2008) indicated that transplanted hESC-derived neurons survived for at least 5 months in a rat model of Parkinson's disease. These investigators used cyclosporine A during the experiment. We also used cyclosporine A to suppress the rat immunological reaction to the transplanted cells; however, transplanted cells did not survive that long perhaps due to inflammatory reaction, or severe condition of the infarcted parenchyma. Further investigation is necessary to determine methods for improving the survival rate of transplanted cells.
Conclusion
This is the first study utilizing eMSCs derived from hESCs in ischemic stroke. Transplantation of eMSCs derived from hESCs through femoral vein resulted in migration and differentiation of stem cells into cells that stained positive for neuronal and endothelial cell markers in the ipsilateral hemispheres of rats subjected to transient focal cerebral ischemia. It also reduced the infarction volume and improved the behavioral functional outcome in ischemic rats. These results suggest the potential clinical applications of cell regenerative treatments in ischemic stroke.
Footnotes
Acknowledgements
We thank Dr Thaddeus G Golos of the Primate Center, University of Wisconsin, and Drs Dan Dan Sun and Raghu Vemuganti of the Department of Neurological Surgery, University of Wisconsin, for reviewing the paper.
The authors have no duality of interest to declare.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.