
Abstract
Ischemia can cause rapid neuronal damage. Previous studies have suggested that synaptic structures and cortical functions can be rescued if therapeutic interventions are applied in time, but the structural basis for this resilience remains incompletely understood. Here, we investigated the restoration of synaptic structures and postischemic plasticity of dendritic spines in the somatosensory cortices of mice by taking advantage of a reversible global cerebral ischemia model. Intravital two-photon imaging revealed that although dendritic structures were rapidly distorted after global ischemia, only a small percentage of spines were actually lost after transient ischemia. Electron microscopy indicated that most presynaptic electron-dense structures were still apposed to postsynaptic densities, and that the majority of disrupted synaptic structures were rapidly reinstated following reperfusion after transient ischemia. Repeated imaging suggested that restored dendrites survived the initial ischemia–reperfusion challenge. Importantly, spines on the restored dendrites underwent a rapid and sustained structural reorganization following transient ischemia. These findings suggested that disrupted synapses during transient ischemia could be rapidly restored after ischemia/reperfusion, and that restored dendritic structures remained plastic to rebuild the cortical network.
Introduction
The brain represents only ∼2% of the body weight but consumes ∼20% of total body's oxygen and glucose for energy supply. 1 Under normal physiological conditions, cortical microcirculation is more than sufficient to support the integrity and activity of the brain network.2–5 However, once the normal blood flow is interrupted by pathological insults such as ischemic stroke, brain tissue will suffer immediate damage.6–8
As one of the most energy-consuming structures of neurons, 9 dendrites and their spines represent the most vulnerable structures after loss of blood supply. Dendrites become beaded and spines are distorted within minutes after energy failure.2,10,11 As the major postsynaptic targets of excitatory synapses, alterations in spine morphology can rapidly alter synaptic transmission and circuit function.12–15 We and others have previously showed that synaptic structures can be rapidly damaged following ischemia,10,11 chilling, 16 oxygen-glucose deprivation, or other pathological insults.17,18 However, the damaged synaptic structures could be restored if these pathological insults were relieved in a timely manner. These findings are particularly important from a therapeutic perspective because cortical neurons are post-mitotic cells and generally do not regenerate. The restoration of synaptic structures offers the hope that synaptic transmission and cortical functions may recover if therapeutic interventions are applied in time. However, the mechanism for reversible restoration of synaptic structures after acute stroke remains incompletely understood. In addition, previous studies mostly focused on the acute effects of ischemia/reperfusion, whether the restored synaptic structures can survive the postischemic reperfusion damage and how the disrupted synaptic structures are reorganized to rebuild the damaged synaptic network remain largely unclear.
In the present study, reversible global cerebral ischemia model combined with electron microscopy (EM) and long-term transcranial two-photon microscopy were used to examine the recovery, survival, and plasticity of dendritic spines of layer 5 pyramidal neurons in the somatosensory cortices of transgenic mice expressing yellow fluorescent protein (YFP). Our results indicated that the disrupted synaptic structures after transient ischemia could be rapidly restored following reperfusion, and importantly, transient global ischemia induced rapid and sustained structural reorganization of dendritic spines.
Materials and methods
Animals
Transgenic mice expressing YFP in layer 5 pyramidal neurons (Thy1-YFP line H, JAX #003782) were purchased from the Jackson Laboratory and bred in animal facilities at Lanzhou University, School of Basic Medical Sciences. All the mice were given free access to water and standard chow diet and kept on a 12-h light/12-h dark cycle at room temperature (22 ±2℃). Mice aged one month of both sexes were used in the experiments. All experiments were performed in accordance with the regulations of Lanzhou University and the ARRIVE guidelines, and all experimental procedures and protocols in the study were approved by the Ethics Committee of Lanzhou University.
Global cerebral ischemia model
Reversible global cerebral ischemia model was used in all experiments. Cerebral ischemia was induced using a modified bilateral common carotid artery ligation (BCAL) model. 19 Mice were deeply anesthetized by intraperitoneal injection of a mixture of ketamine hydrochloride and xylazine (150 mg/kg ketamine and 25 mg/kg xylazine body weight). Rectal temperature was maintained at 37 ± 0.5℃ using a feedback-regulated heating pad during surgery. The bilateral common carotid arteries were carefully exposed and separated from carotid sheath using thin-tipped tweezers to avoid damaging the vagal nerve and then loosely encircled with surgical sutures. To induce ischemia, surgical sutures around the bilateral common carotid arteries were tightened. The animals were subjected to 20 min of transient ischemia, and then the surgical sutures were untied to reperfuse the blood vessels. The reduction of blood flow velocity (three arterioles were measured in each mouse) and damages of dendritic structures (dendritic beadings) during ischemia were evaluated by in vivo two-photon microscopy. BCAL model was considered successful when >90% reduction in blood flow and dendritic beadings were observed within 10 min after BCAL, and the reperfusion of blood flow was also confirmed in all mice subjected to ischemia. Sham-control mice received skull thinning and the same surgical exposure of bilateral common carotid arteries except for the ligation step.
Long-term in vivo transcranial imaging
The surgical procedure for imaging was performed as described previously.11,20 Briefly, the mouse was deeply anesthetized with a mixture of ketamine hydrochloride and xylazine, and body temperature was maintained at 37 ± 0.5℃ using a heating pad. The skull of the mouse was exposed and tightly glued to a metal frame made by sticking two double-edged razor blades face to face together, and then fixed to a custom-made steel plate. A 2 × 2 mm2 area of the skull was thinned using a high-speed drill over the right somatosensory cortex centered on the coordinates of −1.5 mm from bregma and 2.0 mm from midline. For optimal image quality, skull thickness was thinned to ∼25 µm. The animal was then fitted into an Olympus FV1000 two-photon microscope (Olympus, Japan) and apical dendritic tufts of layer 5 neurons were transcranially imaged using a water-immersion objective (25×, 1.05 numerical aperture, Olympus, Japan). For time-lapse imaging of dendritic and spine structures, Ti:sapphire laser was tuned to 920 nm to excite YFP, and repeated Z-series of 40 to 60 optical sections (1024 × 1024 pixel arrays) were collected with a step size of 0.75 µm. The image data were collected at fixed time points after the start of ischemia or reperfusion.
To measure blood flow velocity, the mouse was intravenously injected with 20 µl Texas Red-dextran (10 mg/ml, Invitrogen, USA). A line along the center axis of an arteriole (10–15 µm in diameter) was drawn and the velocity of red blood cells (RBCs) was measured by repeated line scanning.11,21 After imaging, the mouse was put into a separate cage and the animal body temperature was maintained at 37 ± 0.5℃ using a heating pad until the mouse was fully mobile.
Transmission electron microscopy (TEM)
A total of 13 mice divided into three groups (sham group n = 4, 20 min of ischemia group n = 5, and 20 min of ischemia followed by 3 h of reperfusion group n = 4) were used for TEM. Mice were transcardially perfused with mixed fixative containing 2.5% glutaraldehyde and 2% paraformaldehyde in PBS (pH 7.4). 22 The brain of the mouse was removed immediately, and the cerebral cortex from the similar cortical region selected for two-photon imaging was collected, cut into pieces, and then put into the same mixed fixative. After 1 h of fixation, the cortex pieces were cut into smaller pieces and continued to be fixed overnight in fresh fixative at 4℃. The cortex pieces were then washed in PBS, post-fixed for 1 h in 1% osmium tetroxide, dehydrated by ethanol, and embedded with Epon 812. 50 nm ultrathin sections from the upper layers of cortex were made and stained with uranyl acetate and lead citrate, and examined under TEM (JEM-1230, Japan).
Data statistics and analysis
Two-photon image analysis was performed using ImageJ software. Morphological classification of dendritic spines has been described in detail previously.23–25 Here, we divided spines into three main categories: thin spines, mushroom spines, and stubby spines. To quantitate changes in dendritic spines, the same dendritic segments collected at different time points were carefully identified from the 3D image stacks. 26 In all, 100–200 spines were counted in each mouse from three images of dendrites. To describe the pathological changes of spines, we used “lost” when a pre-existing spine was not visible during ischemia, and “restored” when a pre-existing spine disappeared during ischemia and then re-appeared at the same site after reperfusion. To identify the distorted spines during ischemia from 3D image stacks, the image contrast was increased. Percentage of spine elimination was calculated as the number of lost spines/pre-existing number of spines. Percentage of spine formation was calculated as the number of new spines/pre-existing number of spines. About 20–30 images in each mouse were taken randomly for EM analyses. All synapses were counted for analysis of the density of synapses in single photo (measured area of 7.65 µm2). Five to ten synaptic boutons were randomly selected in each mouse for counting the total number of synaptic vesicles per synaptic bouton.
All data analyses were performed using SPSS statistics 20.0. EM results were compared using one-way analysis of variance (ANOVA) followed by post hoc test. Statistical significance in spine formation and elimination rates was determined using independent-samples t-test. Data were presented as mean ± SEM; p < 0.05 was taken as the level of significance.
Results
Disruption and in situ restoration of synaptic structure after transient ischemia/reperfusion
Cortical microcirculation is tightly coupled to neuronal activity and the integrity of neural network is highly dependent on local blood flow.27,28 We first used two-photon in vivo imaging technique to examine the effects of transient global ischemia on synaptic structures by ligating the bilateral common carotid arteries (Figure 1(a) and (b)). In somatosensory cortex where ∼98% reduction in blood flow was induced (Figure 1(d), BCAL 3 min; n = 5 mice, three arterioles in each mouse), we observed that dendrites became beaded and spines were distorted within 10–20 min after ligation (Figure 1(c) and (e), BCAL 20 min). Although dendrites were markedly distorted, two-photon in vivo imaging revealed that most spines were still present on blebbed dendrites, and only 13.2 ± 0.9% of spines disappeared following 20 min of BCAL (Figure 1(e) and (f), BCAL 20 min; n = 5 mice). Among these lost spines, 95.6 ± 3.9% of them were found located at sites where dendritic beadings were formed. We did not observe any newly formed spines during ischemia. After ischemia, the surgical sutures were untied to reperfuse the blood vessels. The blood flow was restored to pre-ischemia level within 3 min after reperfusion (Figure 1(d), reperfusion 3 min). We found most of the beaded dendrites restored within 3 h after reperfusion following 20 min of BCAL, and majority of the lost spines were also restored at this time point (Figure 1(e) and (f), reperfusion 3 h). The restoration of the lost spines was traced following reperfusion, and repeated imaging indicated that 78.8% and 96.9% of them re-appeared at 1 and 3 h after reperfusion, respectively (Figure 1(f), reperfusion 1 and 3 h; n = 5 mice). Remarkably, the majority of restored spines retained their positions and characteristic morphologies after recovery from ischemia/reperfusion (among these identified restored spines, 90.1% mushroom spines, 97.5% stubby spines, and 66.7% thin spines were of the same type as before ischemia at 3 h after reperfusion, n = 5 mice).
Disruption and restoration of dendritic structures following transient global cerebral ischemia/reperfusion. (a) Schematic diagram showing experimental methods for inducing ischemia and for in vivo imaging. (b) Timeline for in vivo imaging. Filled arrowheads indicate the imaging time points being discussed. (c) Two-photon image showing Texas Red-dextran labeled blood vessels (red) and dendritic structures (green) before ischemia, 20 min after ischemia, and 3 h after reperfusion. Dendrites became beaded after transient ischemia and restored following reperfusion. Note the extravasation (leakage of blood plasma, arrowhead) surrounding the blood vessel at 20 min after ischemia. Reduced extravasation around the vessels at 3 h after reperfusion indicated a recovery of BBB. (d) Two-photon line scanning images showing changes in blood flow velocity and flux in an arteriole labeled in (c) (white arrow) before ischemia, 3 min after ischemia, and 3 min after reperfusion. (e) Magnified view of the yellow-boxed region in (c) showing structural changes in dendritic spines during ischemia/reperfusion. Note although dendrites were severely distorted, most spines were still present on blebbed dendrites. Filled arrowhead indicates a stable spine on a narrow filament between dendritic beadings, and chevron indicates a stable spine on a dendritic beading during ischemia, and asterisk indicates a spine partially collapsed down into the dendritic shaft during ischemia. A small percentage of spines transiently disappeared during ischemia, but restored after reperfusion (open arrowhead indicates a spine completely collapsed down into the dendritic shaft during ischemia, and restored after reperfusion). The majority of restored spines retained their positions and characteristic morphologies after ischemia/reperfusion. Arrow indicates a new spine formed during reperfusion. (f) Quantification of lost spines during BCAL/reperfusion. The image contrast was increased to identify the distorted spines during ischemia from 3D image stacks. Only 13.2 ± 0.9% of spines disappeared during 20 min of ischemia (n = 5 mice). Please note that most of the spines only temporarily disappeared during ischemia, and 78.8 ± 4.3% and 96.9 ± 1.4% of these lost spines re-appeared at 1 and 3 h after reperfusion, respectively. Data are presented as mean ± SEM.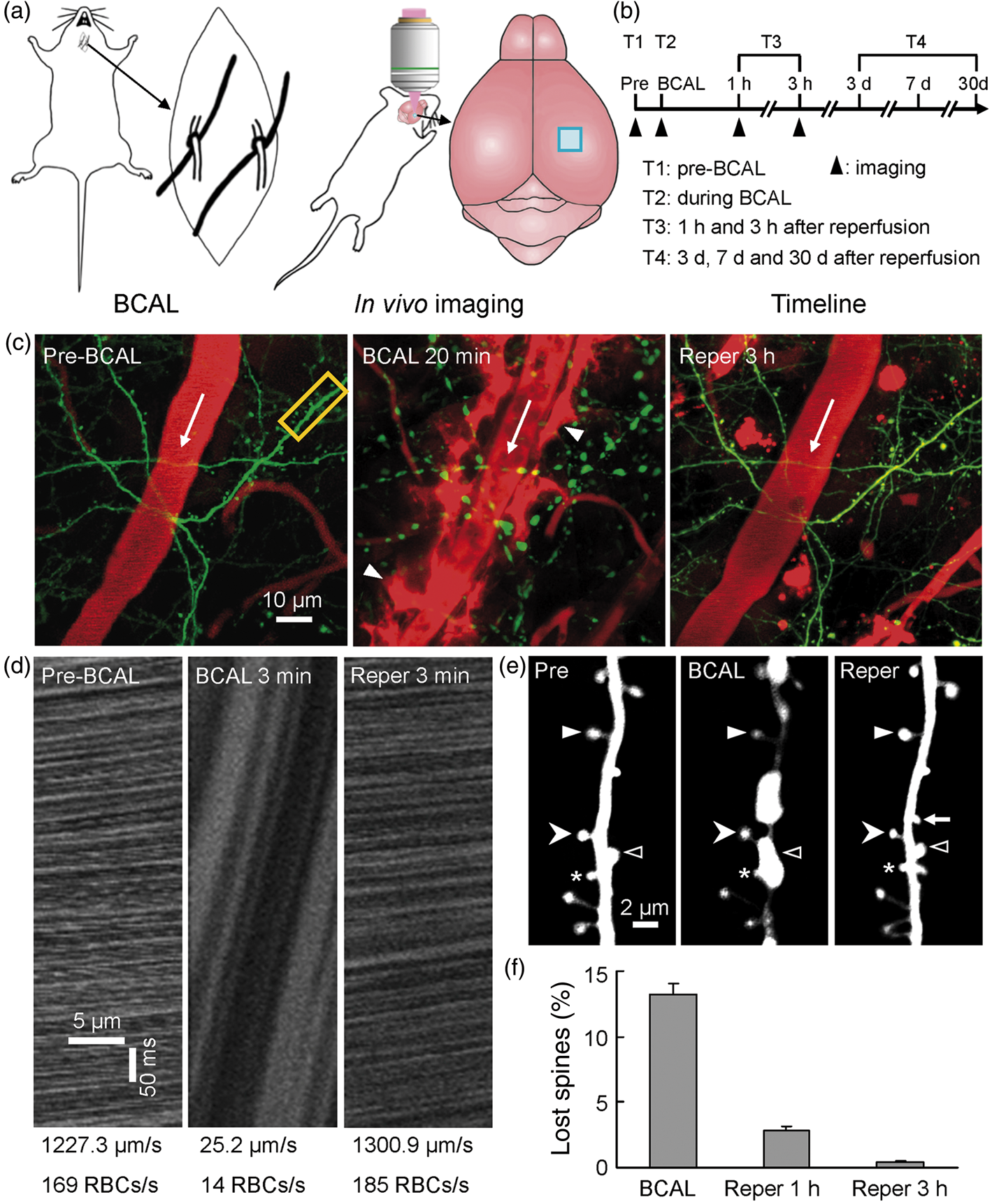
In addition, we observed a leakage of Texas Red-dextran or Evans blue labeled blood plasma within 20 min after BCAL (Figure 1(c) and Supplementary Figure 1, BCAL 20 min), indicating a rapid disruption of blood–brain barrier (BBB) integrity following ischemia. Most of the extravasation was cleared at 3 h after reperfusion, suggesting a restoration of BBB function. We analyzed the change of YFP signal in dendrites during ischemia and after reperfusion and found that YFP signal was greatly reduced during ischemia (Supplementary Figure 2(a) and (c)), but the fluorescence recovered to pre-stroke level at 1 h after transient ischemia. A previous study has suggested a decrease in pH following ischemia. 29 To evaluate whether pH is associated with the change in YFP signal, we performed an experiment to analyze the effect of pH on YFP fluorescence using fixed slices, and found that YFP signal was greatly reduced in acidic solution but can recover when pH was restored (Supplementary Figure 2(b) and (d)). We also assessed the impact of transient ischemia on axons and observed that axons became beaded during 20 min of BCAL, but these damaged axons were restored following 1 h of reperfusion (Supplementary Figure 3). These results indicated that transient ischemia can cause substantial distortion or damage to dendrites and their spines, axons, and BBB, but these changes in structures can be largely restored following reperfusion.
To further understand the structural basis for the restoration of synaptic structures after transient ischemia, we examined changes in organelles, synaptic number and pre- and postsynaptic structures during transient ischemia and after reperfusion with EM (Figure 2). TEM data indicated that microtubules were disrupted in beaded dendrites during ischemia (Figure 2(a), BCAL 20 min, asterisk). 20 min of BCAL also caused drastic changes in synaptic structures (Figure 2(b), BCAL 20 min). The presynaptic and postsynaptic structures such as postsynaptic density were obscured (Figure 2(b), BCAL 20 min, chevron). In addition, the number of synaptic vesicles was significantly reduced during ischemia (Figure 2(c); n = 5 mice for ischemia, n = 4 mice for sham, p = 0.0002). Although synaptic ultrastructures were altered, we found most presynaptic electron-dense structures were still apposed to postsynaptic densities and there was no significant decrease in synapse density during ischemia (Figure 2(d); 1.70 ± 0.06 (/µm2) for sham group versus 1.61 ± 0.07 (/µm2) for ischemia group, p = 0.16), indicating most damaged spines may still have remaining synaptic contacts. Thus, we believed that most spines and synapses were still there although their structures were altered. We further assessed the recovery of organelles and synaptic structures after reperfusion. The structures of microtubules largely re-assembled at 3 h after reperfusion (Figure 2(a), reperfusion 3 h, arrow). But the number of synaptic vesicles was still significantly lower than sham-treated group at 3 h after reperfusion (Figure 2(c); p = 0.008). Noteworthy, the density of synapses at 3 h after reperfusion was significantly higher than those in sham-treated mice (Figure 2(d); 2.00 ± 0.12 (/µm2) for reperfusion group versus 1.70 ±0.06 (/µm2) for sham group from 10 to 20 TEM images per mouse, n = 4 mice for each group, p = 0.016), indicating a net increase on the order of 18% in synapse number after reperfusion.
Changes in dendritic ultrastructure during transient global cerebral ischemia/reperfusion. (a) Electron micrograph showing characteristic dendritic morphology in animals subjected to sham treatment, 20 min of transient ischemia, and 20 min of ischemia followed by 3 h of reperfusion. Healthy dendrites had uniformly distributed microtubules. Microtubules were disrupted and holes in cytoplasm appeared after 20 min of ischemia. Microtubules recovered after reperfusion. Arrows show microtubules and asterisk indicates a dendrite with disrupted cytoplasmic components. (b) Alterations in synaptic ultrastructures during transient ischemia/reperfusion. The number of synaptic vesicles (arrowheads) was reduced after ischemia. Presynaptic active zone and postsynaptic density (chevron) were blurred after ischemia but recovered following reperfusion. (c) Quantification of synaptic vesicles after transient ischemia/reperfusion. (d) Quantification of synapse density during ischemia/reperfusion. There was no significant change in the density of synapses after ischemia (ischemia group: 1.61 ± 0.07 (/µm2), n = 5 mice; sham group: 1.70 ± 0.06 (/µm2), n = 4 mice; p = 0.16), but the density of synapses was significantly increased at 3 h after reperfusion (ischemia/reperfusion group: 2.00 ± 0.12 (/µm2), n = 4 mice; sham group: 1.70 ± 0.06 (/µm2), n = 4 mice; p = 0.016). Data are presented as mean ± SEM, *p < 0.05, **p < 0.01, and ***p < 0.001.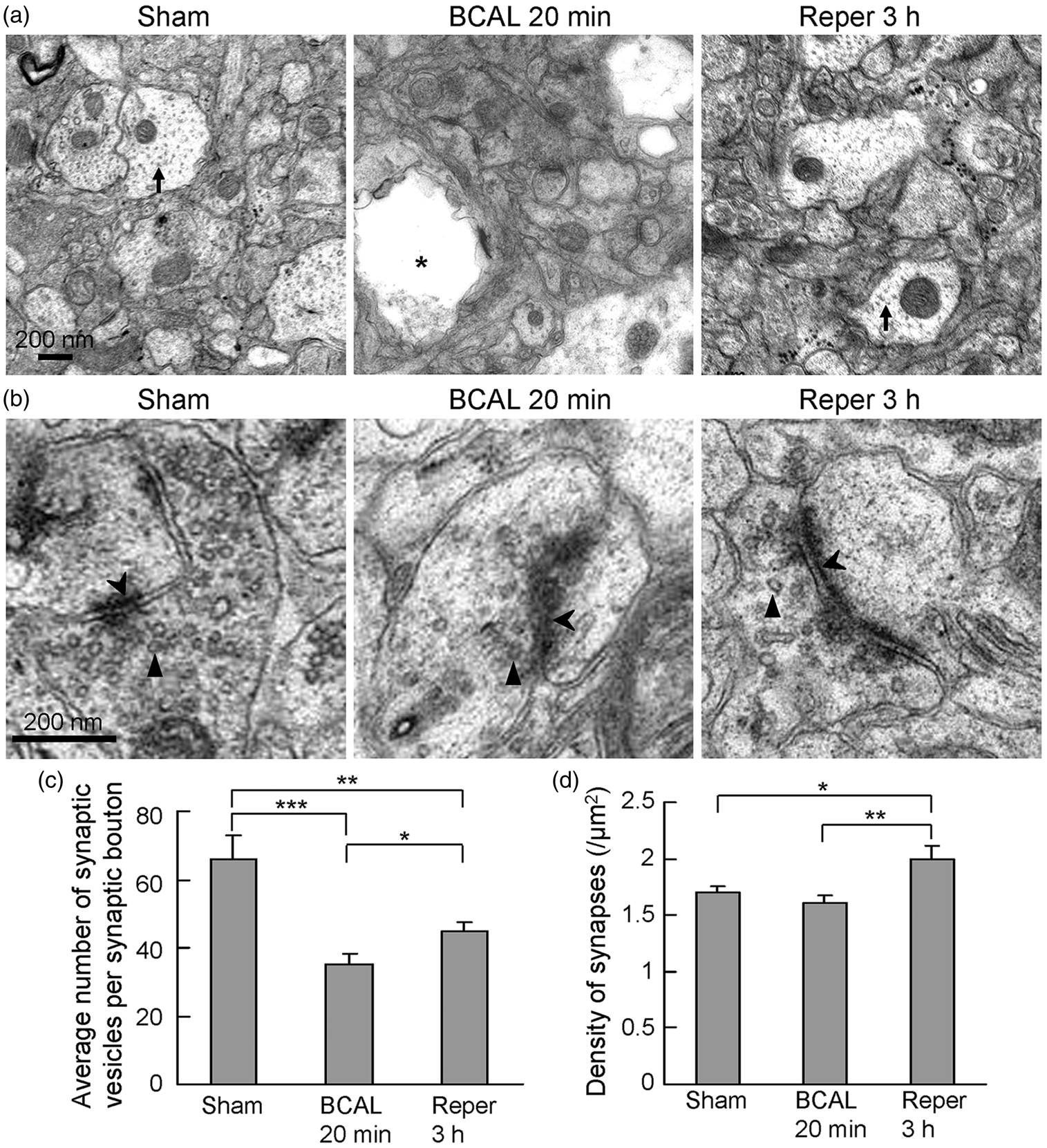
Rapid and sustained reorganization of dendritic spines after the initial ischemia–reperfusion challenge
Our above data suggested that blebbed dendrites rapidly recovered and majority of spines were restored within hours when blood vessels were reperfused after 20 min of ischemia. We further determined the impact of transient ischemia on the stability of synaptic structures (Figure 3(a) to (c)). Remarkably, we observed that dendritic spines on the recovered dendrites experienced a significant reorganization as assessed by the formation and elimination of spines (Figure 3(d) and (e)). In healthy young animals, the rate of spine elimination is normally higher than the rate of spine formation.26,30 Transient ischemia significantly increased the rate of spine formation at 1 and 3 h after reperfusion compared with sham-treated group (Figure 3(d)). The elimination rate of spines between pre-ischemia imaging and 1 h after reperfusion was also significantly increased (Figure 3(e); Pre-1 h, ischemia 5.3 ± 0.8% versus sham 2.1 ± 0.5%, p = 0.011, n = 6 mice for each group). However, the lost spines between pre-ischemia imaging and 1 h after reperfusion included some spines that were not restored from ischemia. When the temporarily lost spines were ruled out, the elimination rate for ischemic group between pre-ischemia imaging and 1 h after reperfusion was not significantly different from that in sham-treated group, indicating transient ischemia/reperfusion preferentially affected spine formation but not elimination. Consistent to this result, there was no significant difference in elimination rate between the ischemia group and sham-treated group when dendrites were largely restored at 3 h after reperfusion (Figure 3(e), Pre-3 h, ischemia 4.7 ± 0.4% versus sham 4.1 ± 0.5%, p = 0.21, n = 5 mice for each group). In addition, the rate of spine elimination in postischemic mice over a 2-h period (Figure 3(e), 1–3 h) during reperfusion was also not changed compared with sham-treated animals. Importantly, the rate of spine formation was higher than the rate of spine elimination in postischemic mice over 2 h during the reperfusion period (1–3 h) (Figure 3(d) and (e), 1–3 h; formation rate: ischemia 5.8 ± 0.1% versus sham 1.4 ±0.4%, p < 0.001, n = 6 mice for each group; elimination rate: ischemia 2.2 ± 0.4% versus sham 2.6 ± 0.5%, p = 0.3, n = 6 mice for each group). When the temporarily lost spines were ruled out, the formation rate was still significantly higher than the elimination rate (formation rate: 3.4% and elimination rate: 2.2%, p < 0.05).
Rapid increase in spine turnover after transient ischemia/reperfusion. (a) Timeline for short-term in vivo imaging. Filled arrowheads indicate the imaging time points being discussed. (b,c) Time-lapse imaging of dendritic structures showing spine turnover in a sham-treated mouse (b) and mouse subjected to 20 min of ischemia/reperfusion (c). Arrows, open arrowheads, and filled arrowheads indicate newly formed spines, eliminated spines, and stable spines, respectively. (d) The percentage of spine formation was significantly higher in postischemic mice compared with sham-treated animals (n = 6 mice for each group, p < 0.05, and p < 0.001). (e) Group data showed that spine elimination rate was significantly increased at 1 h after reperfusion (ischemia/reperfusion group: 5.3 ± 0.8%, n = 6 mice; sham group: 2.1 ± 0.5%, n = 6 mice; p = 0.011), but the rate was not significantly changed when most of spines were restored at 3 h after reperfusion (ischemia/reperfusion group: 4.7 ± 0.4%, n = 5 mice; sham group: 4.1 ± 0.5%, n = 5 mice; p > 0.05), and there was no significant change in the rate of spine elimination over a 2-h period (1–3 h following reperfusion) (ischemia/reperfusion group: 2.2 ± 0.4%, n = 6 mice; sham group: 2.6 ± 0.5%, n = 6 mice; p > 0.05). Data are presented as mean ± SEM, *p < 0.05, and ***p < 0.001.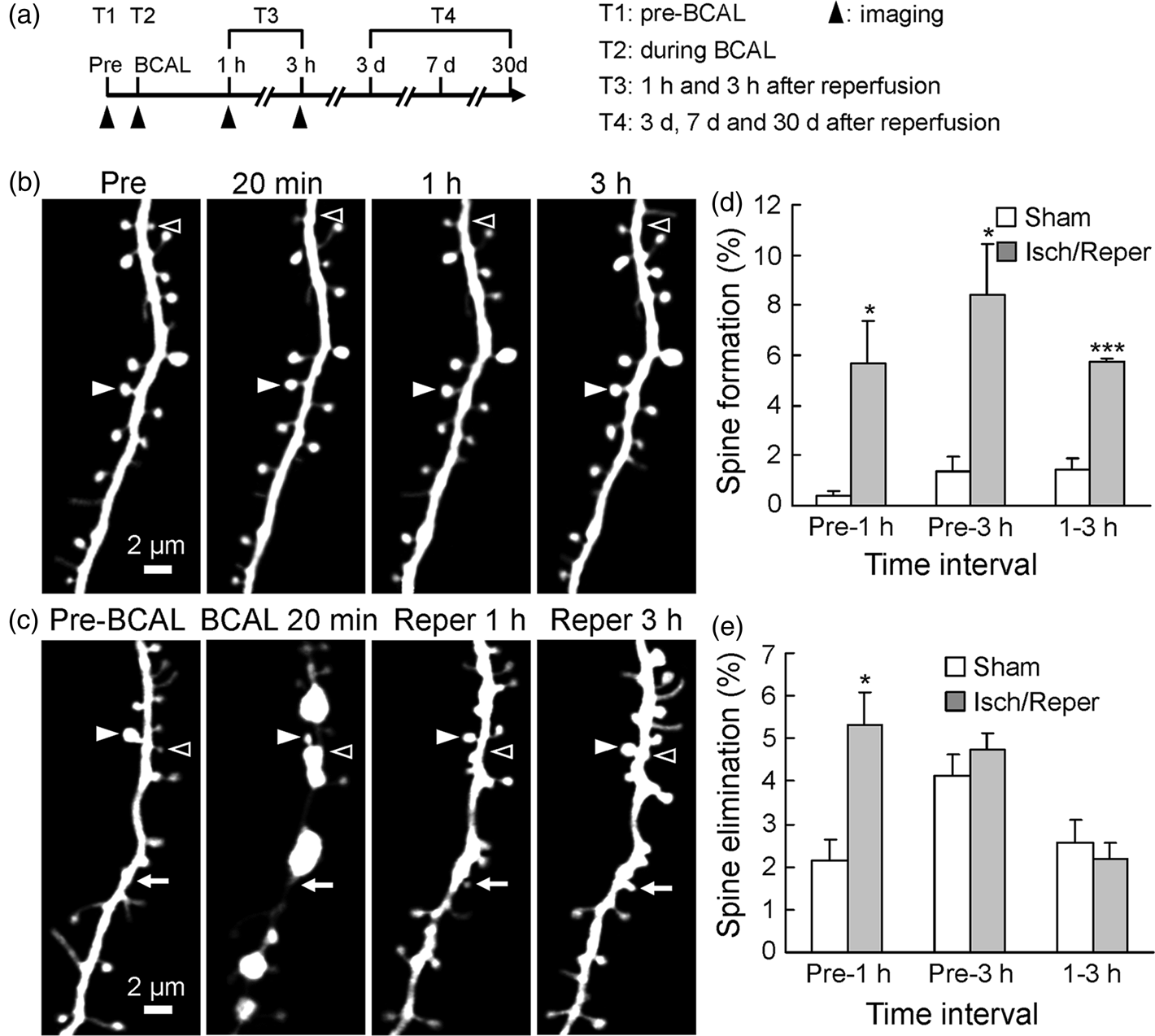
To further evaluate long-term effects of transient ischemia on structural stability of the brain, we traced the restored dendrites for up to 1 month using transcranial two-photon imaging technique (Figure 4(a) to (d)). Repeated imaging suggested that these restored dendrites survived (Figure 4(b)), and spines on these dendrites underwent a sustained reorganization over the next 30 days (Figure 4(d)). Both spine formation and elimination rates were significantly increased in ischemic group compared with age-matched sham control group at 3 days following ischemia/reperfusion (Figure 4(e) and (f); p < 0.05, n = 5 mice). Spine formation rate was still higher over 3–7 days and 7–30 days in ischemic group (Figure 4(e)). However, there was no significant difference in the rate of spine elimination over 3–7 days and 7–30 days following transient ischemia (Figure 4(f)).
Long lasting effect of transient ischemia on the turnover of dendritic spines. (a) Timeline for long-term in vivo imaging. (b) Repeated imaging shows the restored dendrites survived the initial ischemia–reperfusion challenge and were relatively stable over 1 month. (c,d) High-resolution images of dendritic branches show spine formation and elimination in sham control (c) and ischemic (d) mice. The dendritic segment in (d) is from the boxed region in (b). Arrows, open arrowheads, and filled arrowheads indicate newly formed spines, eliminated spines, and stable spines, respectively. (e) The percentage of spines formed is higher over different periods of time in mice subjected to ischemia/reperfusion compared with sham-treated animals (n = 6 mice for each group, p < 0.05). (f) Spine elimination rate increased over the first 3 days after reperfusion but showed no significant changes over 3–7 days and 7–30 days for sham control and ischemic animals. Please note that spines elimination here represents true spine loss and is different from the temporary spine disappearance during ischemia. Data are presented as mean ± SEM, *p < 0.05.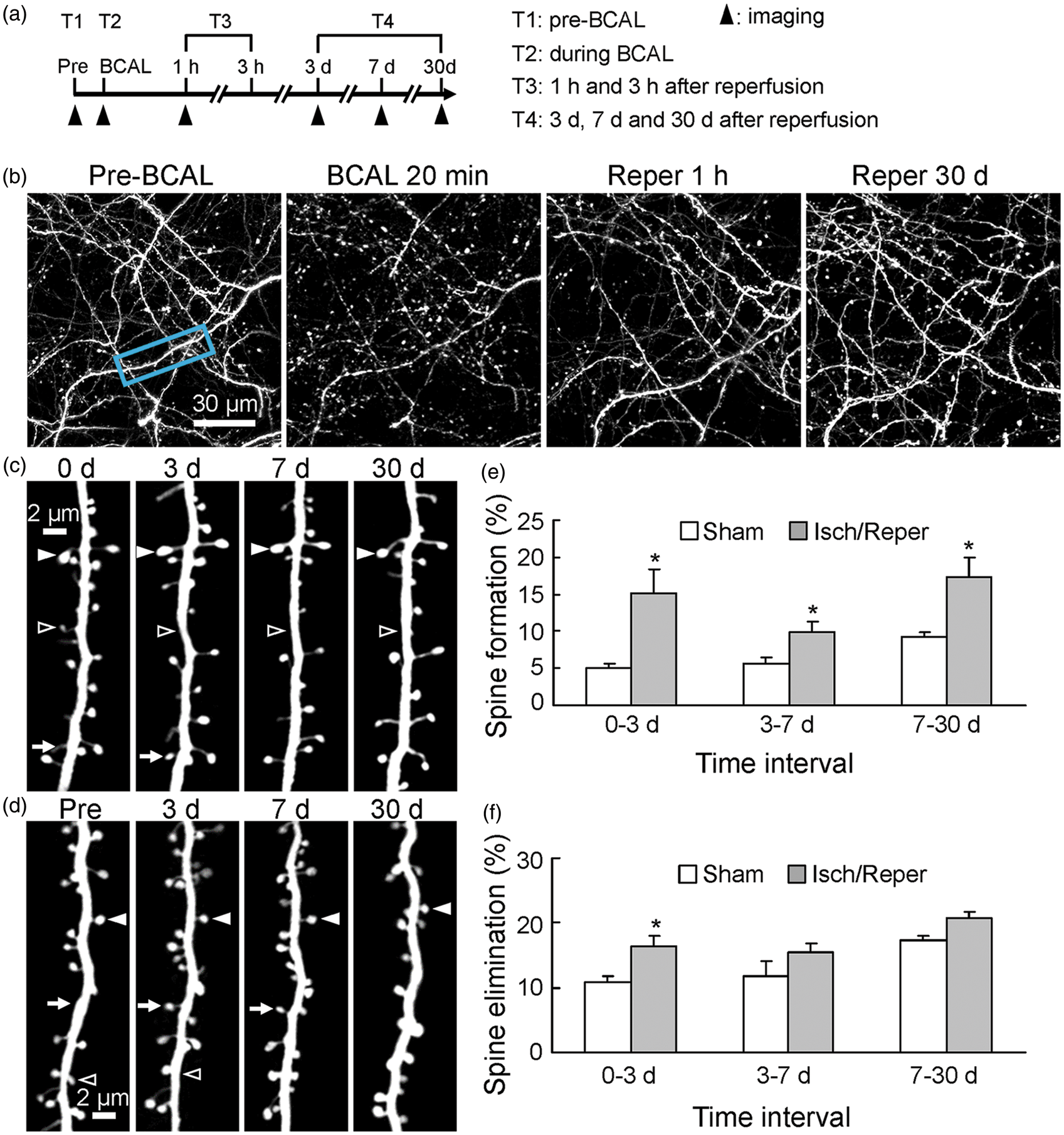
Stabilization of newly formed spines after transient ischemic stroke
Dendritic spines experienced a drastic reorganization and new spines were formed during reperfusion after transient ischemia (Figure 3(d)). What were the fates of these newly formed spines? We traced the turnover of these new spines formed within 1 h following reperfusion (Figure 5(a) to (c)). Repeated imaging indicated that 8.3% of spines generated within 1 h after reperfusion were still present at 30 days after reperfusion (Figure 5(d), ischemia/reperfusion, 1 h to 30 days). We observed that small dendritic protrusions transformed into mushroom-shaped spines (Figure 5(e), arrow), suggesting some newly formed spines were integrated into the cortical circuits. Interestingly, new spines formed within 1 h following reperfusion were more stable than that in sham treated animals over a 2-h period (between 1 and 3 h after start of reperfusion) (Figure 5(b) to (d); 88.9 ± 3.9% for ischemia/reperfusion group versus 33.3 ± 9.6% for sham group, p = 0.0059). However, the stability of new spines that were generated within 1 h after reperfusion over a 3 - or 30-day interval was not significantly different between sham and ischemia/reperfusion group (Figure 5(d); p > 0.05).
Stability of newly formed spines. (a) Timeline for in vivo imaging. Filled arrowheads indicate the imaging time points being discussed. (b,c) Time-lapse imaging of dendritic segments shows spine formation over 30 days in sham control (b) and ischemic (c) mice. Arrows indicate newly formed spines. (d) Stability of new spines formed within 1 h after reperfusion over different periods of time. New spines formed within 1 h following reperfusion were more stable than in sham treated animals over a 2-h period (1–3 h after reperfusion, n = 5 mice for each group, p = 0.0059), but were not significantly different over a 3- or 30-day imaging interval (n = 5 mice for each group, p > 0.05). (e) Example of a spine that was newly formed after transient ischemia/reperfusion and was stable over 30 days (arrows). Data are presented as mean ± SEM, **p < 0.01.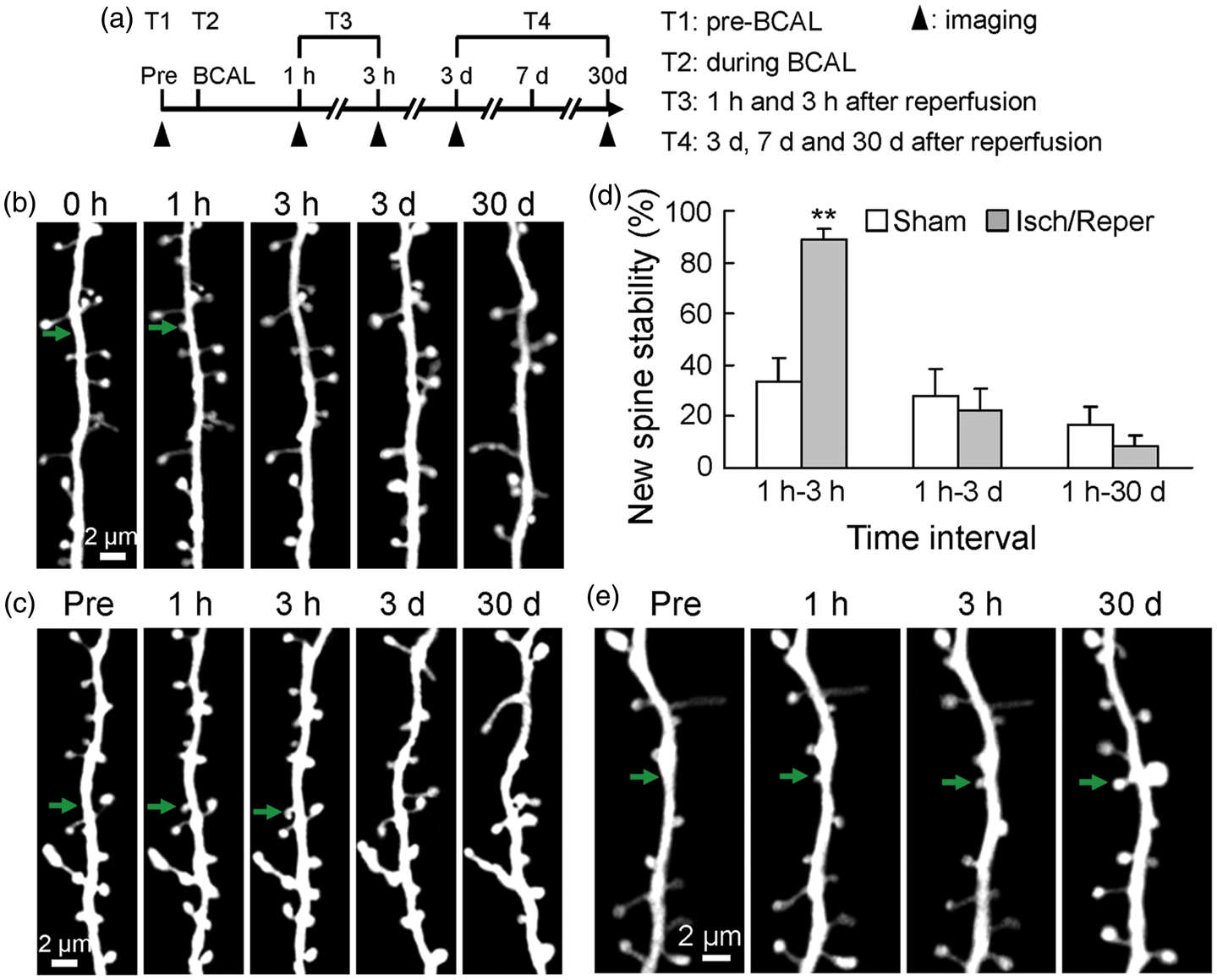
We next evaluated the effects of transient ischemic stroke on animal behaviors (Supplementary Figure 4). All mice which were subjected to transient ischemic stroke survived and there was no significant difference in body weight between the sham and ischemia groups (data not shown). The rotarod test showed that there was a significant difference in walking time on the rotarod between the sham and ischemia group at 12 h after reperfusion (Supplementary Figure 4(a); n = 6 mice for each group, p = 0.041), but there was no significant difference after this time point (p > 0.05), suggesting a rapid recovery in motor coordination ability after transient ischemia. Spontaneous locomotor activity recovered more slowly. We detected a significant difference between the sham and ischemia group in the total distance of spontaneous activity at 1 and 3 days after reperfusion (Supplementary Figure 4(b); n = 6 mice for each group, p < 0.05), but the difference was not significant after 7 days following reperfusion (p > 0.05).
Discussion
Massive disappearance of spines has been reported in ischemia, oxygen-glucose deprivation, NMDA or kainate exposure, chilling and other pathological conditions, but lost spines can be restored if these pathological insults were relieved soon enough.8,10,11,16–18,31 These studies provided vital information about the reversibility of dendritic spines in different experimental conditions, but the underlying structural basis remains unresolved. The present study extended these findings by showing that majority spines or synapses were not permanently lost during transient ischemia and the restoration of dendrites was associated with the reorganization of spines. Although dendrites were heavily distorted following ischemia, here we observed that majority of spines were still attached on beaded dendrites and only ∼13% of spines disappeared after 20 min of ischemia (Figure 1(e) and (f), BCAL 20 min). The difference in the degree of spine loss in different conditions may be related to the nature and severity of the insults. In addition, the signals of fluorescent proteins or dyes and therefore the imaging quality may also affect the quantification of spines. Ischemia caused a drastic fading of YFP signals (Supplementary Figure 2(a) and (c)); thus, the percentage of lost spines could be overestimated due to the quenching of fluorescent signals and severe distortion of dendritic structures. Furthermore, it should be noted that the transient disappearance of a spine during ischemia does not necessarily represent a loss of synapse. 17 EM data showed that there was no significant decrease in the density of synapses after transient ischemia (Figure 2(d), BCAL 20 min), suggesting most lost spines may still have remaining synaptic contacts. Indeed, we observed that most presynaptic structures were still apposed to postsynaptic structures during ischemia even cytoplasmic components were severely disrupted (Figure 2(a), BCAL 20 min). The survived connections may facilitate the in situ restoration of synaptic structures. Consistent with previous in vitro observations,16,17 our in vivo images indicated that most spines disappeared at sites where local dendritic beadings occurred (Figures 1(e) and 3(c), BCAL 20 min, open arrowheads). Therefore, these temporarily lost spines during ischemia were most likely masked by dendritic beadings or collapsed down into the dendritic shafts. 17
We observed a significant loss of synaptic vesicles and obscured pre- and postsynaptic structures after transient ischemia (Figure 2(b) and (c)). Despite the extensive alterations in synaptic structures, repeated live images indicated that the distorted synaptic structures could be rapidly restored in situ after transient ischemia/reperfusion (Figure 1(e) and (f), reperfusion 1 and 3 h). These data are in agreement with previous reports that spines are resilient structures and can recover from various pathological damages.10,11,16 Because spine morphology is highly correlated with its function, restoration of spine structures may contribute to the recovery of synaptic transmission.32,33 Remarkably, we found that the majority of restored spines retained their characteristic morphologies (Figure 1(e), reperfusion 3 h, open arrowheads), suggesting synaptic strength and circuit function can also be restored. Using intrinsic optical signal imaging, a previous study by Murphy et al. has shown that cortical function can be largely restored soon after structural recovery (within 0.75–2 h). 10 Overall, these data indicated that synapses were resistant to ischemic perturbation and could be rapidly restored after reperfusion.
We took advantage of the two-photon imaging technique to monitor the formation and elimination of dendritic spines following ischemia/reperfusion. Dendritic spines are relatively stable but can undergo constant modifications in healthy animals. The stability and plasticity of synaptic structures may provide a structural basis for learning and memory storage,26,34–37and can be altered under pathological conditions.35,38,39 In the present study, we found ∼13% of spines disappeared during transient ischemia (Figure 1(f)), but the majority (∼97%) of these lost spines re-appeared within 3 h after reperfusion. Thus, only a small percentage of spines (<1%) were actually lost after transient ischemia and most of the invisible spines were only temporarily masked or stretched by surrounding dendritic beadings due to energy failure. In addition, we didn’t observe significant formation of new spines during ischemia, suggesting that the physiological spine turnover was largely arrested during ischemia.
Although spine didn't turn over significantly during ischemia, we observed a drastic increase in spine turnover when the blood vessels were reperfused. During ischemia, most spines were heavily distorted and both pre- and postsynaptic structures were altered. The drastic alterations in dendritic structures are associated with a rapid loss of cortical functions. 10 Previous studies indicated that remodeling of neuronal structures and circuits can effectively contribute to restoration of functions after stroke, 40 and increased synaptic plasticity in peri-infarct cortex following focal ischemia was associated with functional recovery or compensations.41,42 Thus, the rapid increase in spine turnover following reperfusion may be associated with the restoration of circuit function. Our results indicated that transient ischemia preferentially affected spine formation but did not affect spine elimination acutely. An increase in the spine formation rate was observed following reperfusion (Figure 3(d)). Although the spine elimination rate at 1 h after reperfusion appeared to be increased (Figure 3(e)), the lost spines at this time point included some temporarily lost spines. Indeed, when dendrites were completely restored, the spine elimination rates between the ischemia group and sham-treated group over a 2-h period (1–3 h) during reperfusion were not significantly different.
Consistent with a net increase in spine formation, our EM data suggested that there was an increase in synapse density following reperfusion (Figure 2(d), reperfusion 3 h). The increase in spine/synapse number may represent an adaptive response of postischemic neurons to compromised cortical functions. Since functional recovery occurs later than structural recovery, 10 cortical functions are transiently compromised and new spine or synapse formation may compensate for the damaged synapse function during recovery. In addition, we found an increased percentage of preexisting spines were lost over 3 days following ischemia/reperfusion, suggesting transient ischemia may cause some permanent changes to cortical networks. The increase in spine turnover may reflect a rebuilding of damaged synaptic circuitry. We found that newly formed spines during acute ischemia/reperfusion could be stably maintained and transformed into mushroom-shaped spines that are generally considered to be synaptic spines (Figure 5(e), arrows), 43 suggesting acutely formed spines were involved in circuit reconstruction. Interestingly, we observed that the new spines generated earlier during reperfusion were more stable, indicating that new spines generated earlier during reperfusion were more likely to form synaptic connections to compensate for the damaged cortical function. Consistent with the observation that transient ischemia preferentially affected spine formation but did not affect elimination within 1–3 h following reperfusion, we found spine formation rates were still higher over 3–7 days and 7–30 days after ischemia (Figure 4(e)), 44 indicating a sustained reorganization of synaptic structures following ischemia. However, we observed a transient increase in spine elimination over 0–3 days (Figure 4(f)). This transient increase in spine elimination may be related to increased plasticity of synaptic structures following ischemia/reperfusion damage.45,46 Although we only monitored dendritic changes in somatosensory cortex, we expect a similar change in other cortical regions considering the global ischemia model we used. Clinical and animal studies suggest that ischemia can cause long-term structural plasticity of cortical neurons.47–49 Our long-term repeated imaging indicated that the rate of spine formation continued to increase over 1 month (Figure 4(e)), indicating transient ischemia had a prolonged effect on circuit stability. The sustained reorganization of synaptic structures may contribute to the rewiring of neuronal circuits and the recovery of cortical functions.
In conclusion, our data suggested that disrupted synaptic structures could be restored after ischemia/reperfusion, and the restored synaptic structures could survive transient ischemia and remain plastic to refine the cortical circuit. However, these conclusions are largely based on the responses of layer 5 cortical neurons and whether synapses of other layers of neurons have a similar response remains to be determined. Further studies to reveal the mechanisms of synaptic reorganization and circuit remodeling following ischemia may help to establish new strategies for clinical treatment of ischemic stroke.
Footnotes
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: National Natural Science Foundation of China (Grant Nos. 81171174 and 31471045) and State Key Laboratory of Neuroscience (SKLN-2015B06).
Acknowledgements
We thank Shaobo Sun, Yana Wang, Minglang Zhou, Chenchen Pan, and Yixin Yan for assistance with preliminary experiments, and Liping Guan and Qingxiang Gao at the Core Facility of School of Life Sciences, Lanzhou University for technical support.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Authors' contributions
L.Z. and S.Z. designed the research plan; L.Z., L.W., and C.W. performed the experiments; L.Z., L.W., F.J., and Y.R. analyzed the data; L.Z. and S.Z. wrote the paper.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.