
Abstract
The obese rodent serves as an indispensable tool for proof-of-concept efficacy and mode-of-action pharmacology studies. Yet the utility of this disease model as an adjunct to the conventional healthy animal in the nonclinical safety evaluation of anti-obesity pharmacotherapies has not been elucidated. Regulatory authorities have recommended employing disease models in toxicology studies when necessary. Our study investigated standard and exploratory toxicology parameters in the high-fat diet (HFD)-induced obese, polygenic Sprague-Dawley rat model in comparison to chow diet (CD)-fed controls. We sought to establish feasibility of the model for safety testing and relevance to human obesity pathophysiology. We report that both sexes fed a 45% kcal HFD for 29 weeks developed obesity and metabolic derangements that mimics to a certain extent, common human obesity. Minor clinical pathologies were observed in both sexes and considered related to CD versus HFD differences. Histopathologically, both sexes exhibited mild obesity-associated findings in brown and subcutaneous white fat, bone, kidneys, liver, lung, pancreas, salivary parotid glands, and skeletal muscle. We conclude that chronic HFD feeding in both sexes led to the development of an obese but otherwise healthy rat. Therefore, the diet-induced obese Sprague-Dawley rat may serve as a suitable model for evaluating toxicity findings encountered with anti-obesity compounds.
Keywords
The growing rise in the global obesity pandemic is an alarming cause for concern. The concomitant health and economic burden associated with this chronic disease are staggering and pose as a public health threat worldwide (Tremmel et al. 2017; Collaborators et al. 2017). Obesity is a disorder of the energy homeostasis system. It is characterized by an imbalance of the biological process involved in precisely matching food intake to that of energy expenditure, in order to achieve long-term body weight stability (Schwartz et al. 2017; Morton, Meek, and Schwartz 2014). Obesity is a complex multifactorial disease (Schwartz et al. 2017), afflicting in addition to overweight individuals, over half of the world’s population today (Organization for Economic Cooperation and Development 2017).
Anti-obesity pharmacotherapies are recommended as an adjuvant treatment for patients with a body mass index (BMI) ≥27 kg/m2 with at least one weight-associated comorbidity (e.g., type 2 diabetes, hypertension, hyperlipidemia) and for those with a BMI ≥30 kg/m2 who have failed lifestyle therapies (Srivastava and Apovian 2018). Anti-obesity compounds are generally designed to induce body weight loss by attenuating food intake, stimulating energy expenditure, or a combination thereof (Srivastava and Apovian 2018; Kim et al. 2014). Emerging anti-obesity pharmacotherapies in development target key neurocircuits involved in the homeostatic regulation of feeding and/or energy expenditure to potentially achieve clinically efficacious weight loss (Srivastava and Apovian 2018; Kim et al. 2014).
Genetic and dietary-induced obese rodent models are a commonly utilized tool to interrogate the complex pathways and genetic underpinnings of energy homeostasis regulation and human obesity pathogenesis (Lutz and Woods 2012; Vickers, Jackson, and Cheetham 2011; Barrett, Mercer, and Morgan 2016). Rodents and humans share similarities in the neuroanatomy and neural circuits involved in the control of energy homeostasis (Barrett, Mercer, and Morgan 2016). It is not surprising that rodents, in general, respond similarly as humans to the food intake and body weight lowering effects induced by clinically effective anti-obesity compounds (Vickers, Jackson, and Cheetham 2011). Obese rodents serve as a commonly utilized animal model of human disease for proof-of-concept efficacy and mode-of-action pharmacology studies, which are critical components of the preclinical development of anti-obesity drug candidates (Morgan et al. 2013).
Despite the conclusion that the obese rodent is an indispensable tool for the aforementioned reasons, this model is rarely used in nonclinical safety testing of novel pharmaceuticals (Morgan et al. 2013). Nonclinical toxicity evaluation and safety assessment are normally conducted in healthy rodents (e.g., Sprague-Dawley [SD] rats and CD-1 mice), based on their stringently controlled health status and well-characterized historical background findings (Morgan et al. 2013; Dixit and Boelsterli 2007). The use of an animal model of disease in nonclinical safety testing poses as a considerable challenge from both a toxicology and a pathology perspective due to the following limitations: (1) poorly characterized species or strain, (2) lack of historical data on spontaneous background findings, (3) limited life span, and (4) considerable heterogeneity in primary or secondary disease-associated pathology (Morgan et al. 2013). These factors may confound the ability to discern whether pathological findings identified in toxicity studies are attributed to (1) incidental age-related or background findings, (2) known primary or secondary manifestations of the disease, (3) drug-related adverse or nonadverse effects, or (4) secondary effects related to exaggerated pharmacology (Morgan et al. 2013).
Nonclinical safety assessment studies in healthy rodent models are estimated to predict approximately 40% of all human adverse events (Dixit and Boelsterli 2007). This suggests that this model may have limited value and may serve as a poor surrogate for a treatment-related adverse event in a human with disease (Dixit and Boelsterli 2007). Safety remains a significant barrier to anti-obesity drug development. Several previously licensed compounds have been withdrawn from the market attributed to safety concerns related to adverse effects (Agahi and Murphy 2014).
The worldwide obesity pandemic has made the development of clinically safe and efficacious anti-obesity pharmacotherapies a long sought goal for patients and clinicians alike (Srivastava and Apovian 2018; Kim et al. 2014). There is clearly an unmet need to enhance predictability of toxicities that may be encountered in the human target population (Morgan et al. 2013). An obese rodent model that closely mimics human obesity may result in improved predictability and understanding of toxicities encountered with anti-obesity compounds (Morgan et al. 2013). Regulatory authorities have recommended employing disease models as an adjunct to the conventional healthy rodent in the safety evaluation of novel pharmaceuticals when relevant (International Conference on Harmonisation of Technical Requirements for Registration of Pharmaceuticals for Human Use and Guidelines 1997).
Our study addressed the question of whether a diet-induced obese (DIO) SD rat model can improve the evaluation of toxicity findings encountered with anti-obesity compounds. We selected the outbred SD rat strain for our studies for three reasons: (1) the healthy, normal weight SD rat is a commonly utilized strain in toxicology studies for pharmaceutical drug development; (2) following chronic exposure to a highly palatable high-fat diet (HFD), the DIO SD rat develops an obese state that shares many characteristics in common with human obesity. These include a polygenic inheritance (Levin et al. 1997; Mercer and Archer 2005) and development of key features of the metabolic syndrome (Madsen et al. 2010); and (3) anti-obesity compounds (e.g., sibutramine and liraglutide) that have a marked effect on body weight in this DIO rat model have a similar effect in humans (Madsen et al. 2010).
The objective of our study was to establish the DIO SD rat for safety testing and ensure that this animal disease model has human relevance. Our specific aims were to address the following criteria as stipulated by the regulatory authorities (Morgan et al. 2013): (1) characterize spontaneous background changes and unintended manifestations of the obese rat model in comparison to healthy, normal weight control rats and (2) determine the level of homogeneity of this model with respect to human obesity pathophysiology. We conducted a longitudinal study, which characterizes the standard toxicology parameters (i.e., clinical pathology and histopathology) in SD rats fed either a 45% kcal HFD (akin to the obesogenic Western diet [Cordain et al. 2005]) or a chow diet (CD) for up to 33 weeks of age. We employed an in-depth exploratory analysis of the endocrine, metabolic, and bone and reproductive consequences of chronic HFD feeding in SD rats beyond the standard approach of a traditional toxicology study. We report that 29 weeks of HFD feeding led to the development of an obese but otherwise healthy SD rat model. Histopathologically, both sexes exhibited only mild obesity-associated disease manifestations and spontaneous background findings comparable to that of normal control animals. We conclude that the DIO SD rat model may be ideally suited for nonclinical safety assessment of anti-obesity drug candidates.
Material and Method
Animals and Dietary Treatment
Cohort one consisting of 32 male and 32 female SD rats (NTac: SD) and, subsequently (2 weeks later), cohort two consisting of 16 male and 16 female SD rats were obtained from Taconic Biosciences (Ejby, Denmark). On arrival, all animals were 4 weeks of age, pair-housed under specific pathogen-free conditions in a temperature-controlled room (20–22°C, 45–65% relative humidity) with a 12-hr:12-hr light: dark cycle. Four-week-old rats were provided with ad libitum (ad-lib) access to water and either standard CD, n = 24 per gender, comprising 3.19 kcal/g, 12% fat, 64% carbohydrate, and 24% protein (Altromin diet 1320; Altromin Spezialfutter GmbH & Co. KG, Lage, Germany), or a 45% kcal HFD, n = 24 per gender, comprising 4.73 kcal/g, 45% fat, 35% carbohydrate, and 20% protein (D12451, Research Diets, New Brunswick, NJ) for a total of 29 weeks. All animals were provided with environmental enrichment consisting of plastic rat shelves, wood gnawing blocks, Enviro-Dri nesting material (Brogaarden, Gentofte, Denmark), and supplemented twice weekly with Trio Munch grains (containing an equivalent energy level as the Altromin diet). Animals were identified individually with an implantable subcutaneous microchip identification device (UNO BV, The Netherlands). All experimental protocols were approved by the Danish Animal Experiments Inspectorate under the Danish Ministry of Environment and Food of Denmark and the Novo Nordisk Ethical Review Council.
Experimental Design, Clinical Observations, and Exclusion Criteria
Clinical observations were performed twice weekly, and both food intake and body weight were measured twice a week. Whole-body composition (total fat and lean mass) was measured in ad-lib-fed rats at 13, 20, and 32 weeks of age (between 12:00 p.m. and 2:00 p.m.) using an EchoMRI™ scanner system (EchoMRI LLC, Houston, TX). Any animals incidentally single-housed or exhibiting adverse clinical signs (e.g., hunched posture, body weight loss >20%, inactivity, decreased food consumption) and associated abnormal changes in clinical pathology were subject to early termination and excluded from the study, which was the case for two HFD-fed females.
Estrous Cycle Measurements
Estrous cyclicity in female rats was assessed at 17 and 30 weeks of age. Vaginal smears were harvested using a sterile cotton tip moistened with 0.9% saline for 10 consecutive days (between 9:00 a.m. and 11:00 a.m.). Smears were transferred to glass slides, fixed with methanol, air-dried, and stained with hematoxylin and eosin (H&E) before microscopic examination (10×). Staging into estrous, metestrous, diestrous, and proestrous were assessed independently by two researchers, and the animal group identities were masked to the observers. The estrous stages were the four different cell types as previously described (Cora, Kooistra, and Travlos 2015). One complete estrous cycle was defined as proestrous–estrous–metestrous–diestrous of 4 to 5 day duration. Animals exhibiting a cycle duration of 4 to 5 days were classified as regular, whereas rats exhibiting shorter or longer cycle length and/or extended estrous (>48 hr) were classified as irregular.
Clinical Pathology Analysis
For coagulation parameters, blood was collected from conscious ad-lib-fed rats via sublingual vein puncture at 12 and 19 weeks of age. Additionally, the same blood sampling method was used for hematology and clinical chemistry parameters, collected at 13 and 20 weeks of age (between 9:00 a.m. and 11:00 a.m.). Blood for coagulation was placed into sodium citrate–containing Eppendorf tubes, subsequently centrifuged, and plasma was separated, aliquoted, and stored at −20°C for subsequent assay analysis. Blood for hematology was collected into ethylenediaminetetraacetic acid (EDTA) syringes, requiring no centrifugation and analyzed the same day at room temperature. Blood for clinical chemistry was collected into Eppendorf tubes and allowed to clot for 1 hr at room temperature, subsequently spun, followed by serum separation and storage in plain Cobas vials at −20°C for subsequent assay analysis.
Blood was also collected at necropsy (33 weeks of age) via the abdominal aorta puncture under inhalational isoflurane anesthesia in ad-lib-fed rats. These samples were processed for coagulation, hematology, and clinical chemistry parameters per the same aforementioned protocol. All clinical pathology measurements were performed by CiToxLAB Scantox A/S, (Lille Skensved, Denmark). Coagulation measurements were determined by IL Test™/ACL™ (Instrumentation Laboratories, Automated Coagulation Laboratory, Milano, Italy). Hematology measurements were directly determined by ABX Pentra DX120SPS (Montpellier, France). The mean cell volume, mean cell hemoglobin, and mean cell hemoglobin concentration were calculated by the ABX Pentra DX120SP. Clinical chemistry measurements were determined by Cobas 6000 with standard Roche kits (Roche Diagnostics, Mannheim, Germany). The ion selective electrode using Cobas 6000 determined sodium, potassium, and chloride values. Finally, sorbitol dehydrogenase (SDH) and β-hydroxybutyrate (β-HB) values were measured by Cobas 6000 using kits from Instruchemie and Randox, respectively.
Plasma Hormone Analysis
Sublingual blood samples were collected at 13 and 20 weeks of age and by abdominal aorta puncture at necropsy (33 weeks of age) in ad-lib-fed rats (at the same time as the sample collection for clinical pathology). Blood was placed into appropriate EDTA-treated tubes, centrifuged, plasma separated, aliquoted, and stored at −80°C for subsequent plasma hormone assay analysis. Insulin and insulin-like growth factor 1 (IGF-1) levels were analyzed using in-house-generated luminescent oxygen channeling immunoassay as previously described (Tsonkova et al. 2018). Briefly, for quantification of insulin levels, the assay used anti-rat insulin HUI018 mAb conjugated acceptor beads, biotinylated polyclonal guinea pig antibody pAb 4077 (raised against rat insulin), and generic streptavidin-coated donor beads. For quantification of IGF-1 levels, the assay used anti-rat IGF-1 mAb (clone 4F38) conjugated acceptor beads, biotinylated mAb clone 1F6 (also raised against rat IGF-1), and generic streptavidin-coated donor beads. During the assay, a bead–analyte–immune complex was created, resulting in light output which was measured on a Perkin Elmer Envision reader. The lower limit of quantification (LLOQ) of the assay for insulin and IGF-1 was 0.12 and 20 ng/ml, respectively. Leptin levels were measured using the AlphaLisa assay kit (Cat no. AL521C; PerkinElmer, Waltham, MA) per manufacturer instructions with the following modification: A calibrator row made from WHO International Leptin Standard (NIBSC code 97/626) was used as a substitute for the kit calibrator. The LLOQ for leptin assay is 1.2 ng/ml. Insulin, IGF-1, and leptin assays were performed by Research Bioanalysis (Novo Nordisk A/S, Måløv, Denmark). Adiponectin, osteocalcin, and c-terminal telopeptides of type I collagen [CTX-1] levels were measured, respectively, by rat ELISA adiponectin kit (Cat no. EZRADP-62 K; Merck Life Science A/S, Hellerup, Denmark), Rat-MID™ osteocalcin EIA (Cat no. AC-12F1; IDS immunodiagnosticsystem, Frankfurt, Germany), and RatLaps™ EIA (Cat no. AC-06F1; IDS immunodiagnosticsystem), per manufacturer instructions. Pituitary hormones (follicle-stimulating hormone [FSH], luteinizing hormone [LH], prolactin, adrenocorticotropic hormone [ACTH], and thyroid-stimulating hormone[TSH]) and thyroid hormone (thyroxine [T4]) were measured by Envigo CRS Limited (Cambridgeshire, UK) using Milliplex MAP Magnetic Bead Panel kits (Rat Pituitary kit, Cat no. RPTMAG-86K; Rat Steriod Thyriod Hormone kit, Cat no. STTHMAG-21K; Rat Bone Panel 1 kit, Cat no. RBN1MAG-31K). These analytes were detected by Luminex MagPix 4.2 per manufacturer instructions (Merck Life Science A/S). Testosterone, progesterone, and estradiol levels were measured by Envigo CRS Limited using EIA kits (Cat. nos., DX-EIA-1559, DX-EIA-1561 and DX-EIA-5774; DRG International Inc., Springfield, NJ) per manufacturer instructions.
Homeostasis Model Assessment–Insulin Resistance (HOMA-IR) Index
The HOMA-IR index was used to quantify insulin resistance and pancreatic β-cell function (Cacho et al. 2008). The formula for this method is insulin (µU/ml) × glucose (mg/dl)/2,430. This simple, accurate index has been validated in both humans (Matthews et al. 1985) and rats (Cacho et al. 2008) against the hyperinsulinemic-euglycemic clamp-derived insulin sensitivity index (SI), the known gold standard approach for measuring in vivo whole-body insulin sensitivity.
Urinalysis
Thirty two-week-old rats were removed from their home cages in the morning (7:00 a.m.), placed individually in metabolic cages, and provided with ad-lib access to water. These animals were fasted during the 4-hr urine collection. Urine samples obtained were subsequently stored at 4°C and analyzed the same day by Siemens Multistix 10 SG/Clinitek Advantus for the following parameters: specific gravity, pH, protein, leucocytes, nitrites, blood, glucose, ketones, bilirubin, and urobilinogen. Urine colors were visually assessed by the following scores: light yellow, yellow, dark yellow, brown, dark brown, and reddish brown. The urine turbidities were also visually examined and assigned one of the following scores: clear, light turbid, turbid, and very turbid. Subsequently, the urine samples were spun for microscopic examination (40×) of the sediment and the following parameters were examined: erythrocytes, leucocytes, epithelial cells, crystals, urates, hyaline and granular casts, and bacteria. The incidence of these parameters in the spun sediment was assessed by the following scores: no trace (no trace in 2–3 visual fields), trace (a few in 2–3 visual fields), slight (a few in each visual field), moderate (several in each visual field), and marked (numerous in each visual field). Analysis of urinalysis parameters were performed by CiToxLAB Scantox A/S.
Terminal Investigations
The rats were terminated at 33 weeks of age and subjected to a complete necropsy. The terminal body weights were recorded, and blood was collected via abdominal aorta for clinical pathology and plasma hormonal analysis. The necropsies were performed at the same time (between 7:00 a.m. and 11:00 a.m. for females and between 10:00 a.m. and 3:00 p.m. for males) over several days, and rats were terminated in a sequence to allow satisfactory intergroup comparisons. Macroscopic findings were recorded. Selected organs were weighed (all paired organs were weighed together) according to Table 1. The following organs were sampled and fixed in phosphate-buffered neutral 4% formaldehyde for histological examination: adrenal glands, aorta, bone (femur/tibia with femur–tibial joint, bone marrow [sternum], and bone marrow smear [humerus]), brain, brown fat (interscapular), epididymides, esophagus, heart, intestine (small and large), kidneys, larynx, liver, lung (with bronchi), lymph nodes (mandibular and mesenteric), mammary glands (inguinal region), ovaries, pancreas, parathyroid glands, Peyer’s patch, pituitary gland, prostate gland, salivary glands (parotid, mandibular and sublingual), sciatic nerve, seminal vesicles, skeletal muscle, skin, spinal cord (cervical, thoracic and lumbar), spleen, stomach (glandular and nonglandular), thyroid glands, tongue, trachea, thymus, urinary bladder, uterus (with cervix), vagina, and white fat (gonadal, retroperitoneal, and inguinal depots). Furthermore, eyes, optic nerves, and harderian glands were preserved in Davidson’s fixative, and testes were fixed in modified Davidson’s fixative. White fat samples were collected from three different regions: the gonadal and retroperitoneal depots representing the intra-abdominal white fat and the inguinal depot representing the subcutaneous white fat (Berryman et al. 2011; Sackmann-Sala et al. 2012). The brain was trimmed into six levels as described by Bolon et al. (2013), excluding the olfactory bulb level. The sternum and left femur/tibia were decalcified in EDTA. The tissues were embedded in paraffin, sectioned at approximately 4 to 5 µm, and stained with H&E. Liver cryosections from selected HFD-fed animals were stained with Oil Red O to verify the presence of neutral lipid droplets. When appropriate, a five-grade severity system was used (minimal, mild, moderate, marked, and severe).
Effect of Diet-induced Obesity on Group Mean Absolute and Adjusted Organ Weight Values (Grams) in Rats at 33 Weeks of Age.
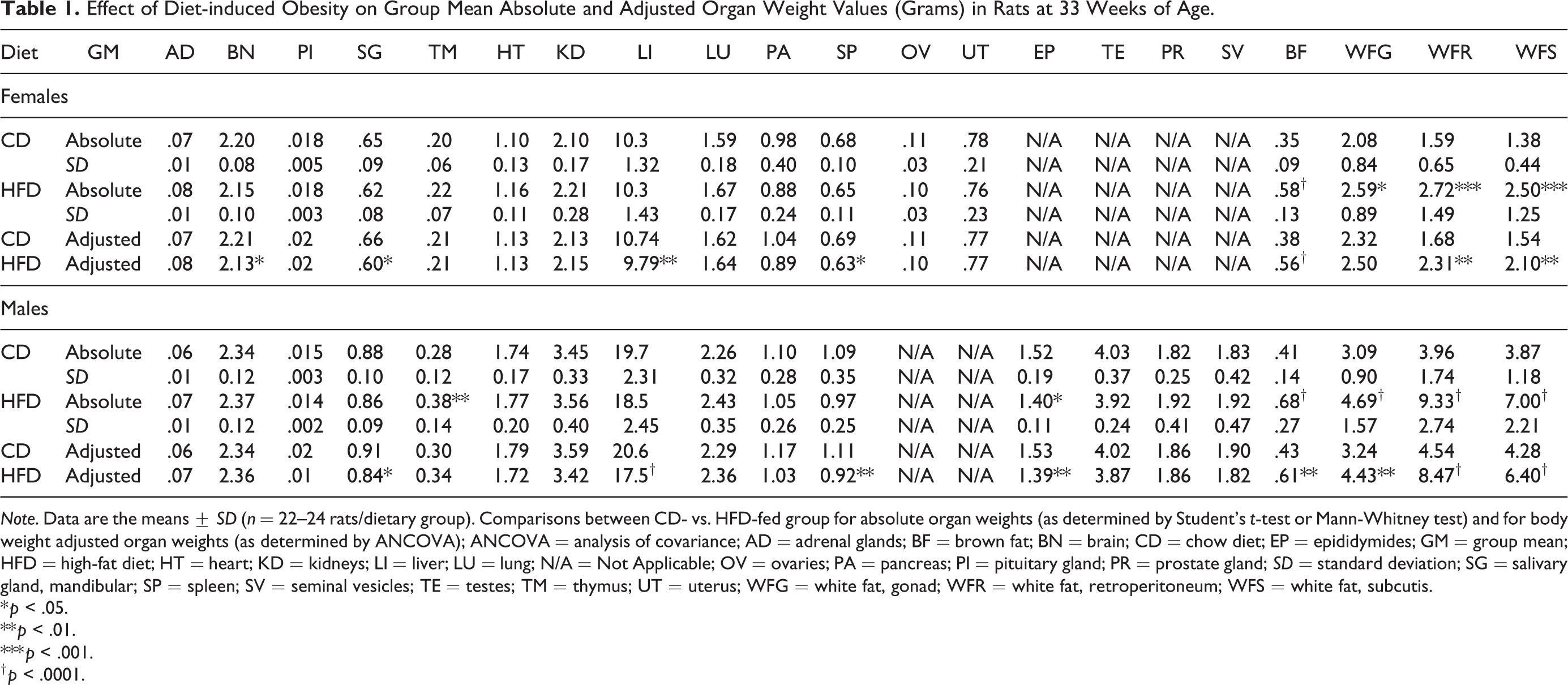
Note. Data are the means ± SD (n = 22–24 rats/dietary group). Comparisons between CD- vs. HFD-fed group for absolute organ weights (as determined by Student’s t-test or Mann-Whitney test) and for body weight adjusted organ weights (as determined by ANCOVA); ANCOVA = analysis of covariance; AD = adrenal glands; BF = brown fat; BN = brain; CD = chow diet; EP = epididymides; GM = group mean; HFD = high-fat diet; HT = heart; KD = kidneys; LI = liver; LU = lung; N/A = Not Applicable; OV = ovaries; PA = pancreas; PI = pituitary gland; PR = prostate gland; SD = standard deviation; SG = salivary gland, mandibular; SP = spleen; SV = seminal vesicles; TE = testes; TM = thymus; UT = uterus; WFG = white fat, gonad; WFR = white fat, retroperitoneum; WFS = white fat, subcutis.
* p < .05.
** p < .01.
*** p < .001.
† p < .0001.
Proximal Tibia Bone Growth Plate Analysis
The cessation of longitudinal bone growth for the proximal tibia epiphysis plate does not occur until after 10 months of age in rats (Smith, Varela, and Samadfam 2017). Hence, this allows for the examination of the effect of HFD feeding on the tibia growth plate. The section of the decalcified left tibia bone including the femur–tibial joint was cut in the sagittal center plane to ensure the optimal presence of the proximal tibial growth plate. Subsequently, the fixed paraffin-embedded bone tissues were cut at nominal thickness of 4 µm (RM2255 Microtome; Leica, Denmark), mounted on Superfrost microscope slides, and stained with H&E. The stained tibia sections were scanned in the Aperio AT2 slidescanner at 40× magnification. The height of the epiphyseal growth plate of the proximal tibia was determined in total by measuring the length of the proliferative and the hypertrophic zone as previously described (Eshet et al. 2004). The length of the hypertrophic zone was calculated using the total height minus the length of the proliferative zone. Fifteen random selected measurements perpendicular to the growth plate were made in all animals using the ruler utility in the Leica’s Aperio ImageScope program, and group identities were masked to the observer.
Histomorphometry Analysis of Osteoclasts in the Proximal Tibia Bone
Paraffin-embedded tibia bone sections were cut at nominal thickness of 4 µm followed by immunohistochemistry staining with the osteoclast marker, CD-68. The sections were deparaffinized in xylene and rehydrated in decreasing concentrations of ethanol. Antigen retrieval was performed over night at 60°C in Tris-EGTA (TEG) buffer at pH 9.0. Endogen peroxidase and alkaline phosphatase activity were blocked by a 10-min incubation with dual block (Cat no. S2003, DAKO, Glostrup, Denmark) followed by a 30-min incubation with 3% Bovine Serum Albumin (BSA). The primary antibody CD-68 (Cat no. Ab125212, Abcam, Cambridge, UK) was diluted 1:750 in a Tris-Buffered Saline (TBS) buffer containing 3% BSA and incubated 30 min at room temperature. CD-68 immunoreactivity was visualized using the Brightvision Goat anti-Mouse-AP (Cat no. VWRKDPVM55AP, ImmunoLogic, Duiven, The Netherlands) added to 20% rat serum and developed with permanent Red (Cat no. K0640, DAKO, Glostrup, Denmark). The slides were counterstained with Mayer’s hematoxylin (Sigma-Aldrich, Søborg, Denmark) to detect cell nuclei and scanned by Leica’s Aperio AT2 slidescanner at 40× magnification. The osteoclast and the trabecular bone were quantified under masked animal group identity examination, using scanned bone section imported into the newCAST software (7.02.3195, Visiopharm, Hoersholm, Denmark). The sampled area of interest (AOI) was chosen to be just below the epiphyseal growth plate in the proximal tibia bone with a depth of 1,412 µm excluding the tibial tuberosity. The trabecular bone volume and the volume fraction of the osteoclasts were estimated by point counting stereological techniques and random sampling of view fields controlled by the newCAST software covering ∼25% of AOI as previously described (Recker 1983; Howard and Reed 2010). The trabecular bone volume fraction (TB VF, %) was calculated as the ratio of the trabecular bone to the total area (marrow and trabecular bone) in the AOI. The CD-68-positive osteoclasts were counted within the vicinity of the bony tissue and osteoclast volume fraction (osteoclast VF, %) quantified by the following equation (Howard and Reed 2010):
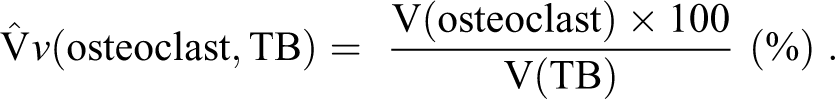
Statistical Analysis
All results are expressed as means ± standard deviation. There was no interaction identified for the effect of cohort on the phenotype (body weight, fat mass, and lean mass) at 13, 20, and 32 weeks of age as determined by a repeated measurement model. Therefore, all data for cohort one and two were combined for presentation and analyzed in a model with additive effects of cohort and treatment. Initially, a bimodality hypothesis was tested based on the observed variation in body weight gains for the animals in response to chronic HFD feeding (Online Supplemental Figure 1). Specifically, a normal (unimodal) and a mixture of two normal (bimodal) distribution models with a cohort effect were fitted to the data, separately for each sex and diet (i.e., CD vs. HFD for comparison). Model selection between a unimodal and a bimodal distribution was by the Bayesian information criterion. For both sexes in the CD group, body weight gains fit a unimodal model. For HFD-fed females, the two components collapsed, and it was only possible to fit a unimodal model. For HFD-fed males, the two component model was degenerate, with one of the components exhibiting almost zero variance, and therefore, the unimodal model was selected.
For single time point data, a two-sample unpaired Student’s t-test (normally distributed data) or a Mann-Whitney test (data not normally distributed) was used for two-group comparisons using GraphPad Prism version 7.04, 2017 (GraphPad Software, San Diego, CA). Multiple time point data were analyzed using a repeated measurement model with multiplicity adjusted post hoc test of treatment effect (SAS, Institute Inc., Cary, NC, USA). Organ weights were evaluated based on both absolute (Student’s t-test or Mann-Whitney test) and body weight–adjusted values by analysis of covariance (ANCOVA) using terminal body weights as a covariate (SAS), in accordance to the recommendation by Jarvis et al. (2011). The urinalysis parameters were analyzed by χ2 test for trend for group incidence and severity (GraphPad Prism). The ratio of regular to irregular estrous cycle duration between the dietary groups for the females was assessed by Fisher’s exact test (GraphPad Prism). In all instances, p value of <.05 was considered statistically significant.
Results
Mortality and Clinical Signs
No deaths and no abnormal clinical signs were observed in either sex in both the CD and HFD group. One HFD-fed female rat was terminated at 25 weeks of age due to the observation of body weight loss, hunched posture, and abnormalities in clinical pathology. Macroscopically, the animal showed enlarged kidneys and dilated ureters, and the histopathological examination revealed severely dilated pelvis of the kidneys. Since this finding was observed in only one animal, it may be a congenital lesion (Suttie, Leininger, and Bradley 2018) and considered unrelated to dietary treatment. Thus, all data for this rat were excluded from the study.
The HFD-fed DIO Phenotype
The HFD was initiated at 4 weeks of age and resulted in both sexes gaining greater body weight and whole-body fat mass than CD-fed controls by 13 weeks of age, and these parameters remained significantly higher at all ages (Table 2). Both sexes at 32 weeks of age achieved a modest increase in body weight (8% for females vs. 9% for males) and a 2-fold increase in fat mass (Table 2). Both sexes on either diet exhibited a wide range of body weight gains after 29 weeks of dietary feeding (Online Supplemental Figure 1A, B). Yet the body weight gains for both sexes were normally distributed (see Statistical Analysis subsection for further details), which agrees with previous studies (Archer et al. 2003; Mercer and Archer 2005). The obesity induced by feeding the HFD was attributed, in part, to the higher daily caloric intake (∼7.7%) observed throughout the study in both sexes (Online Supplemental Table 1). HFD feeding significantly attenuated lean mass in males at 20 and 32 weeks of age, which did not occur in the females (Table 2).
Effect of Chronic HFD Feeding on Body Weight and Body Composition in Rats at 13, 20, and 32 Weeks of Age.
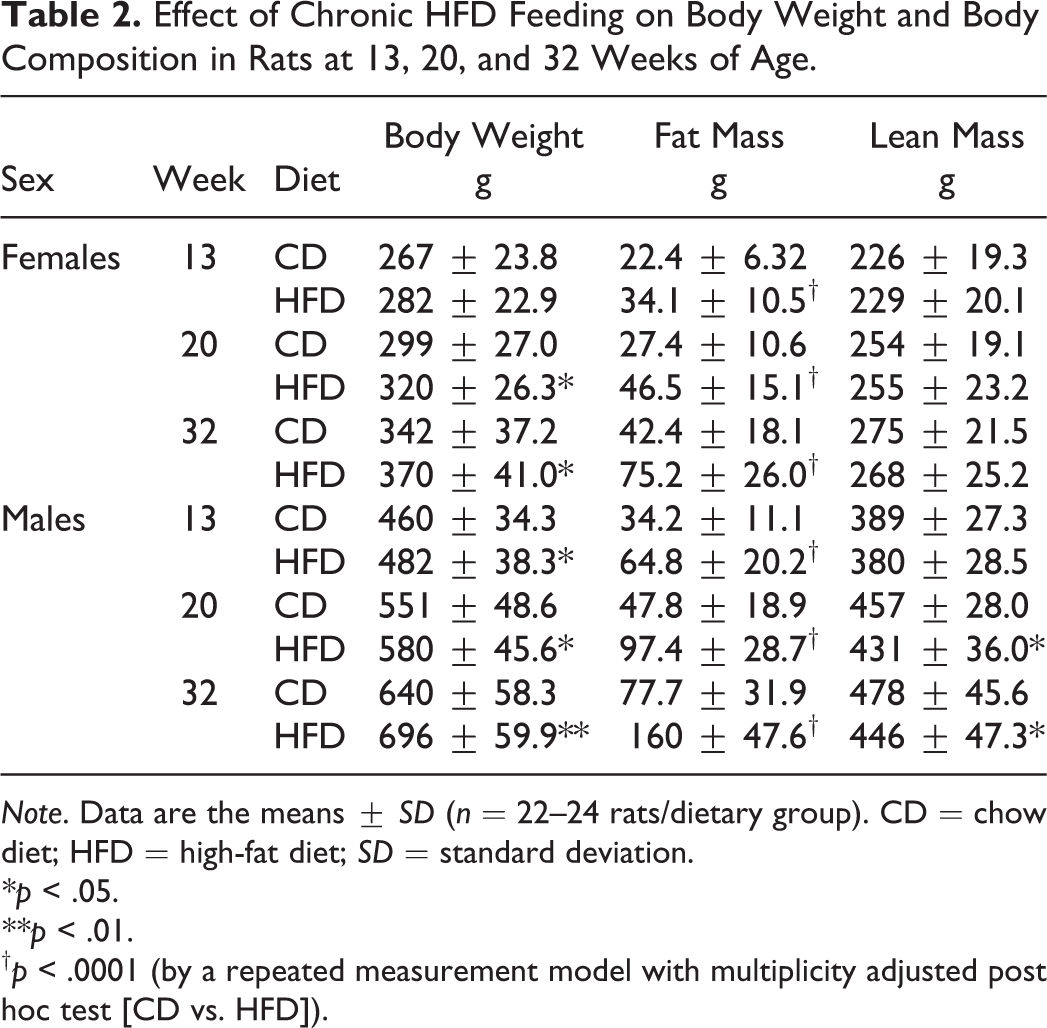
Note. Data are the means ± SD (n = 22–24 rats/dietary group). CD = chow diet; HFD = high-fat diet; SD = standard deviation.
* p < .05.
** p < .01.
† p < .0001 (by a repeated measurement model with multiplicity adjusted post hoc test [CD vs. HFD]).
We determined the impact of HFD feeding in SD rats on components of the metabolic milieu, by assessing several hormonal, metabolite, and plasma lipid biomarkers relative to CD-fed controls over the course of the study. Circulating levels of TSH, T4, and ACTH were attenuated in both sexes at 33 weeks of age (Online Supplemental Table 1). Leptin levels were significantly elevated by 2- to 3-fold at all ages in the HFD-fed females and males, respectively (Table 3). HFD-fed females exhibited an initial increase in basal plasma insulin levels by 66% (13 weeks of age) and had a subsequent tendency for higher insulin levels throughout the study (Table 3). HFD-fed males achieved an increase in plasma insulin levels that remained significantly elevated by 2-fold at 20 and 33 weeks of age (Table 3). Both levels of insulin and leptin were highly correlated with the degree of adiposity at study termination (Online Supplemental Table 2), which is a hallmark manifestation of diet-induced obesity.
Impact of Chronic HFD Feeding on Hormonal and Metabolic Milieu in Rats at 13, 20, and 33 Weeks of Age.
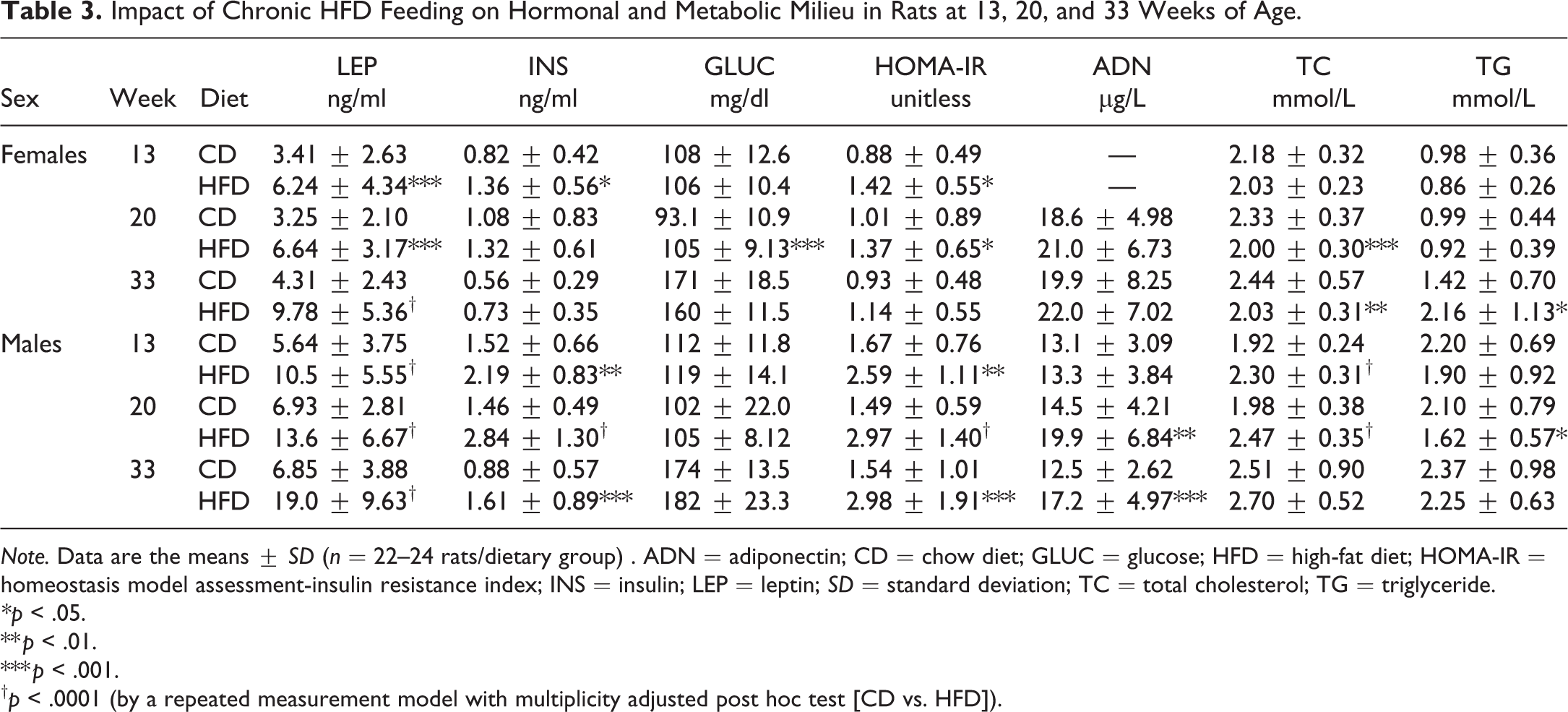
Note. Data are the means ± SD (n = 22–24 rats/dietary group) . ADN = adiponectin; CD = chow diet; GLUC = glucose; HFD = high-fat diet; HOMA-IR = homeostasis model assessment-insulin resistance index; INS = insulin; LEP = leptin; SD = standard deviation; TC = total cholesterol; TG = triglyceride.
* p < .05.
** p < .01.
*** p < .001.
† p < .0001 (by a repeated measurement model with multiplicity adjusted post hoc test [CD vs. HFD]).
There were only minimal alterations of basal plasma glucose levels throughout the study. However, the HOMA-IR index remained higher at all ages in these HFD-fed animals (Table 3), indicating increased insulin resistance. The degree of insulin resistance (HOMA-IR) and adiposity was highly correlated in all animals (males: r = .8, p < .0001; females: r = .5, p = .0003). Plasma levels of adiponectin were increased by ∼38% in only the HFD-fed males (Table 3). Minimal fluctuations in circulating total cholesterol (TC) were observed at all ages; HFD-fed females and males exhibited a minor decrease and increase, respectively, in these levels (Table 3). Unlike the males, HFD-fed females exhibited a mild increase in plasma triglycerides (TG) by 52% at 33 weeks of age (Table 3). TG levels were highly correlated to HOMA-IR (r = .44; p = .0023) in the females.
Hematology, Coagulation, and Clinical Chemistry Parameters
Assessment of hematology parameters, in comparison to CD-fed animals (Table 4 and Online Supplemental Table 3), revealed that HFD-fed males exhibited a minor decrease in both red blood cell count and hematocrit at all ages. These were accompanied by minor increases in mean cell hemoglobin and mean cell hemoglobin concentration at 20 and 33 weeks of age. At all ages, there was an observed trend for a minimal decrease in white blood cell counts for both sexes in the HFD group. For the HFD-fed females, this correlated with the observed decrease in total neutrophil and lymphocyte counts observed at 13 and 20 weeks of age. Platelets (PLT) were lower in both sexes in the HFD group at all ages. These were accompanied by a marginal shortening of activated partial thromboplastin time (APTT) and prothrombin time (PT) and a decrease in fibrinogen (FIB) levels (Table 5).
Effect of Diet-induced Obesity on Hematology Parameters in Rats at 13, 20, and 33 Weeks of Age.
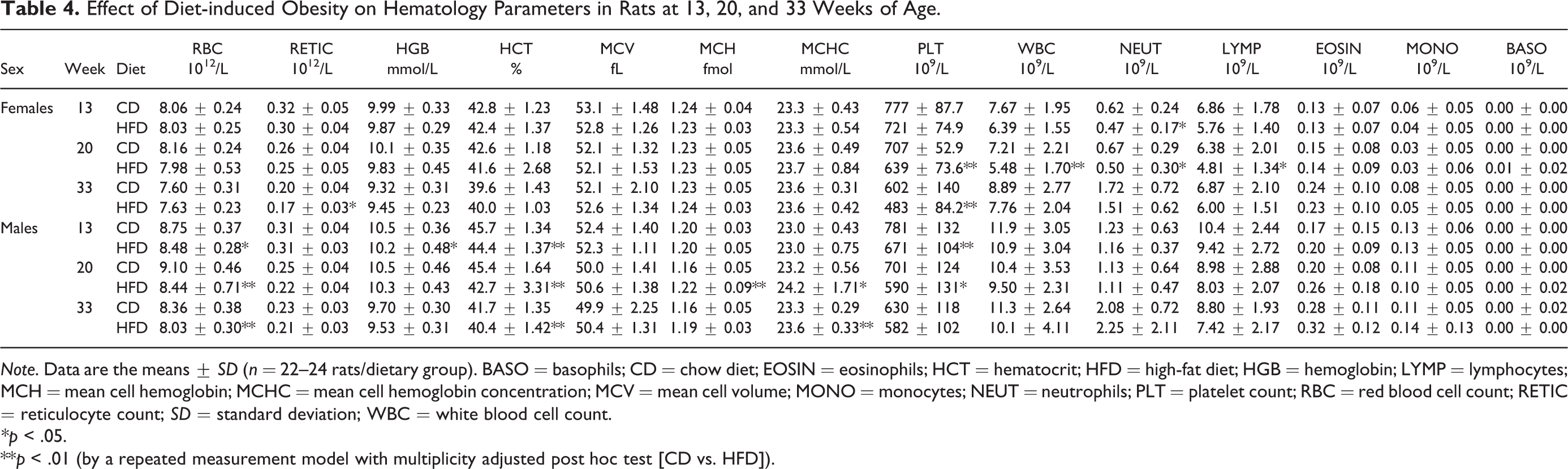
Note. Data are the means ± SD (n = 22–24 rats/dietary group). BASO = basophils; CD = chow diet; EOSIN = eosinophils; HCT = hematocrit; HFD = high-fat diet; HGB = hemoglobin; LYMP = lymphocytes; MCH = mean cell hemoglobin; MCHC = mean cell hemoglobin concentration; MCV = mean cell volume; MONO = monocytes; NEUT = neutrophils; PLT = platelet count; RBC = red blood cell count; RETIC = reticulocyte count; SD = standard deviation; WBC = white blood cell count.
* p < .05.
**p < .01 (by a repeated measurement model with multiplicity adjusted post hoc test [CD vs. HFD]).
Effect of Diet-induced Obesity on Coagulation Parameters in Rats at 12, 19, and 33 Weeks of Age.
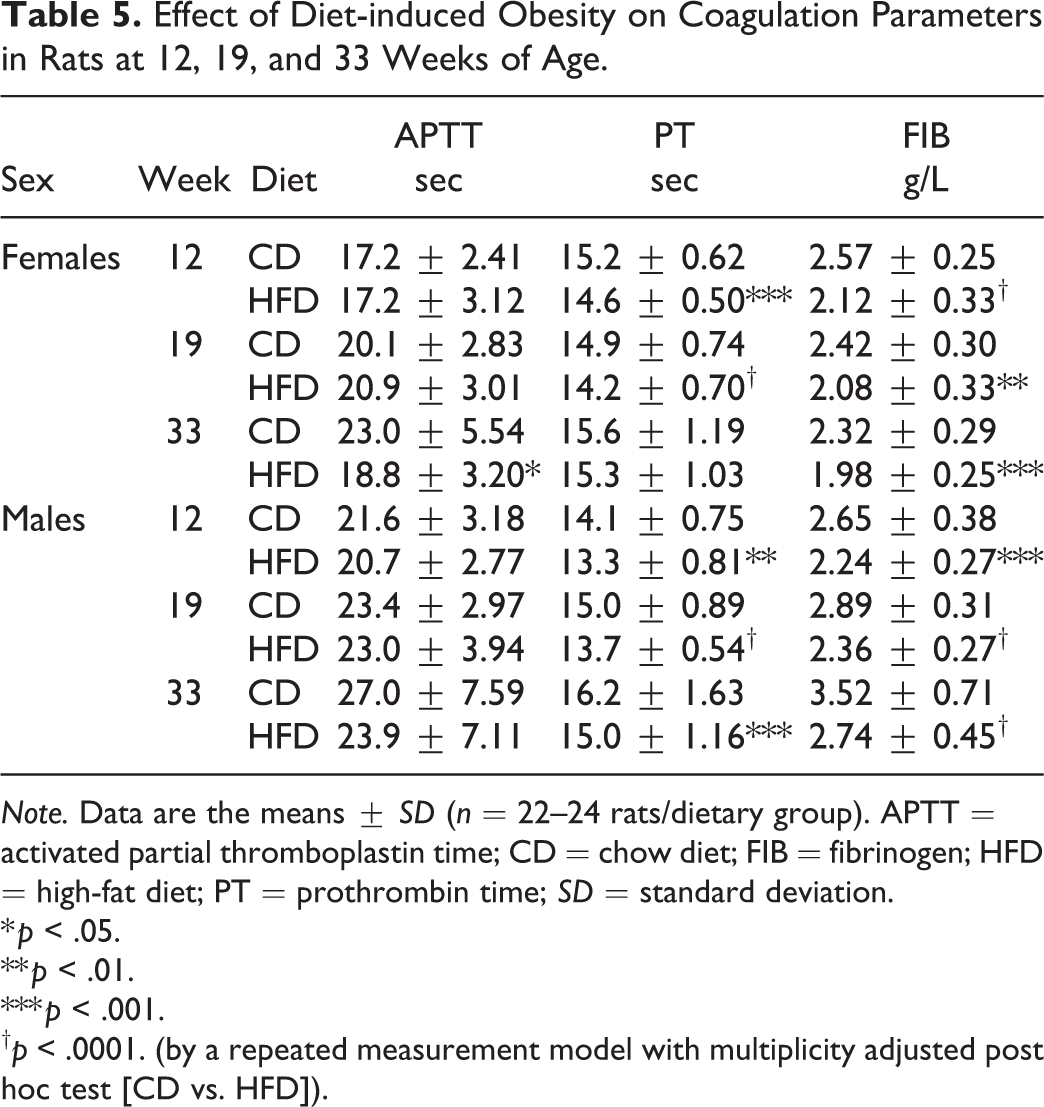
Note. Data are the means ± SD (n = 22–24 rats/dietary group). APTT = activated partial thromboplastin time; CD = chow diet; FIB = fibrinogen; HFD = high-fat diet; PT = prothrombin time; SD = standard deviation.
* p < .05.
** p < .01.
*** p < .001.
† p < .0001. (by a repeated measurement model with multiplicity adjusted post hoc test [CD vs. HFD]).
Assessment of clinical chemistry parameters (Table 6) revealed a minor increase in total bilirubin levels in both sexes in the HFD group by 20 and 33 weeks of age. Furthermore, a small but consistent decrease in alanine aminotransferase (ALAT) and aspartate aminotransferase (ASAT) was observed in the HFD-fed animals in both sexes at all ages. These changes were accompanied by a minor decrease in both urea (carbamide) and total protein levels in the HFD-fed males by 20 and 33 weeks of age. In contrast to HFD-fed males, total protein levels trended to increase in HFD-fed females at 20 and 33 weeks of age primarily due to an increase in albumin levels. SDH levels were increased by ∼2-fold and accompanied by a decrease in β-HB in both sexes in the HFD-fed group at 20 and 33 weeks of age. Assessment of plasma electrolytes (Table 6) revealed that magnesium (Mg) was consistently decreased in both sexes in the HFD-fed group at all ages. Apart from a minor trend for an increase in chloride levels, no other electrolyte changes were observed. Minimal fluctuations in creatinine were observed: increased in HFD-fed females (at all ages) and decreased in males (33 weeks of age; Table 6).
Effect of Diet-induced Obesity on Clinical Chemistry Parameters in Rats at 13, 20, and 33 Weeks of Age.
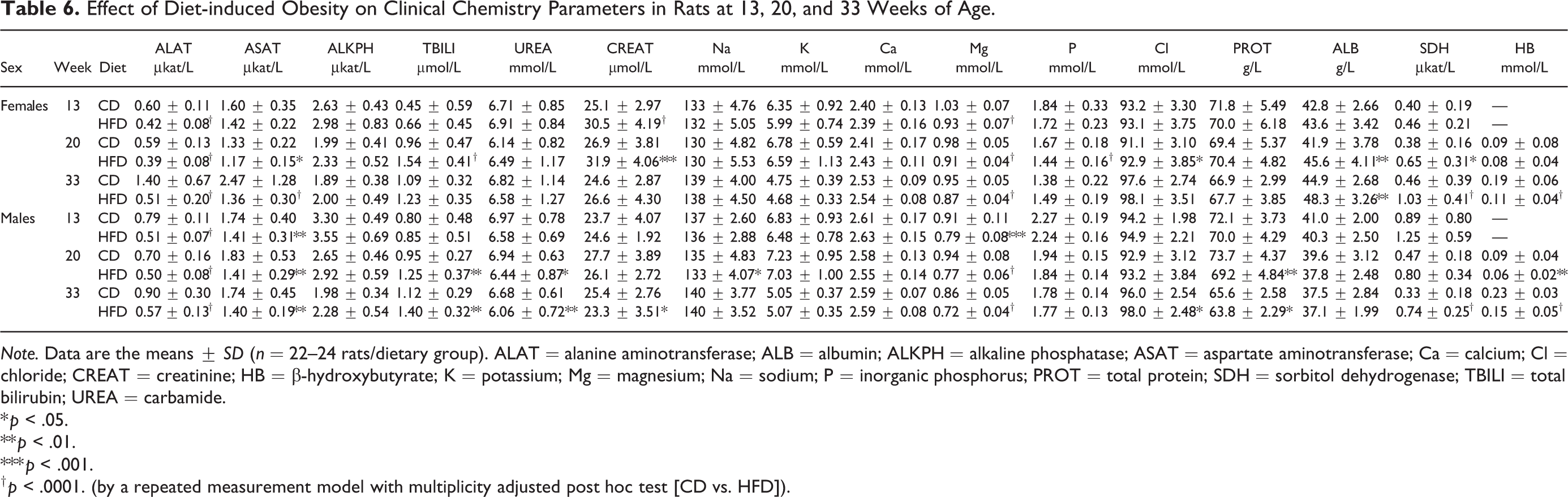
Note. Data are the means ± SD (n = 22–24 rats/dietary group). ALAT = alanine aminotransferase; ALB = albumin; ALKPH = alkaline phosphatase; ASAT = aspartate aminotransferase; Ca = calcium; Cl = chloride; CREAT = creatinine; HB = β-hydroxybutyrate; K = potassium; Mg = magnesium; Na = sodium; P = inorganic phosphorus; PROT = total protein; SDH = sorbitol dehydrogenase; TBILI = total bilirubin; UREA = carbamide.
* p < .05.
** p < .01.
*** p < .001.
† p < .0001. (by a repeated measurement model with multiplicity adjusted post hoc test [CD vs. HFD]).
Urinalysis
Assessment of urinalysis parameters revealed that a lower pH, ranging from 6.0 to 7.5, was observed at a higher incidence in both sexes in the HFD group, in comparison to the CD-fed rats (Online Supplemental Table 4). In the HFD group, a higher incidence of animals exhibited a lower urinary content of ketones and epithelial cells (males), crystals and urates (both sexes), and bacteria (females; Online Supplemental Tables 4 and 5). There were no differences between the dietary groups for both sexes in regard to urinary specific gravity, protein, leucocytes, and presence of blood (Online Supplemental Table 4). Nor were there any differences in nitrites, glucose, bilirubin, urobilinogen, hyaline, and granular casts (data not shown).
Organ Weights
Both sexes in the HFD group had higher body weights at termination, which can influence absolute organ weights. Hence, the data were evaluated based on both absolute and body weight–adjusted values by ANCOVA (Bailey, Zidell, and Perry 2004; Jarvis et al. 2011). An increase in absolute weights was observed for the brown adipose tissue (BAT) and the three white adipose tissue (WAT) depots in both sexes in the HFD group (Table 1). When adjusted for body weight, all weights except the female gonadal fat pad were increased. The degree of hyperinsulinemia and hyperleptinemia correlated positively with the weights of WAT depots in both sexes (Online Supplemental Table 2). Both the absolute and the adjusted epididymides weight were decreased in the HFD-fed males. Decreases in the adjusted weights of the spleen, liver, and salivary gland in both sexes and the brain in females were observed without any differences in the absolute weights. Increases in absolute thymus weight in the males were observed in the HFD group (Table 1).
Histological Findings
HFD-feeding-related histopathological findings in the animals were observed in BAT, subcutaneous WAT, femur, kidneys, liver, lung, pancreas, parotid salivary glands, and skeletal muscle (Tables 7 and 8). There were no histological differences observed between the dietary groups in the remaining organs (Online Supplemental Table 6A, B). Organs without histological findings were not included in the Online Supplemental Tables.
Histological Findings in Female Rats at 33 Weeks of Age—Group Incidence and Severity.
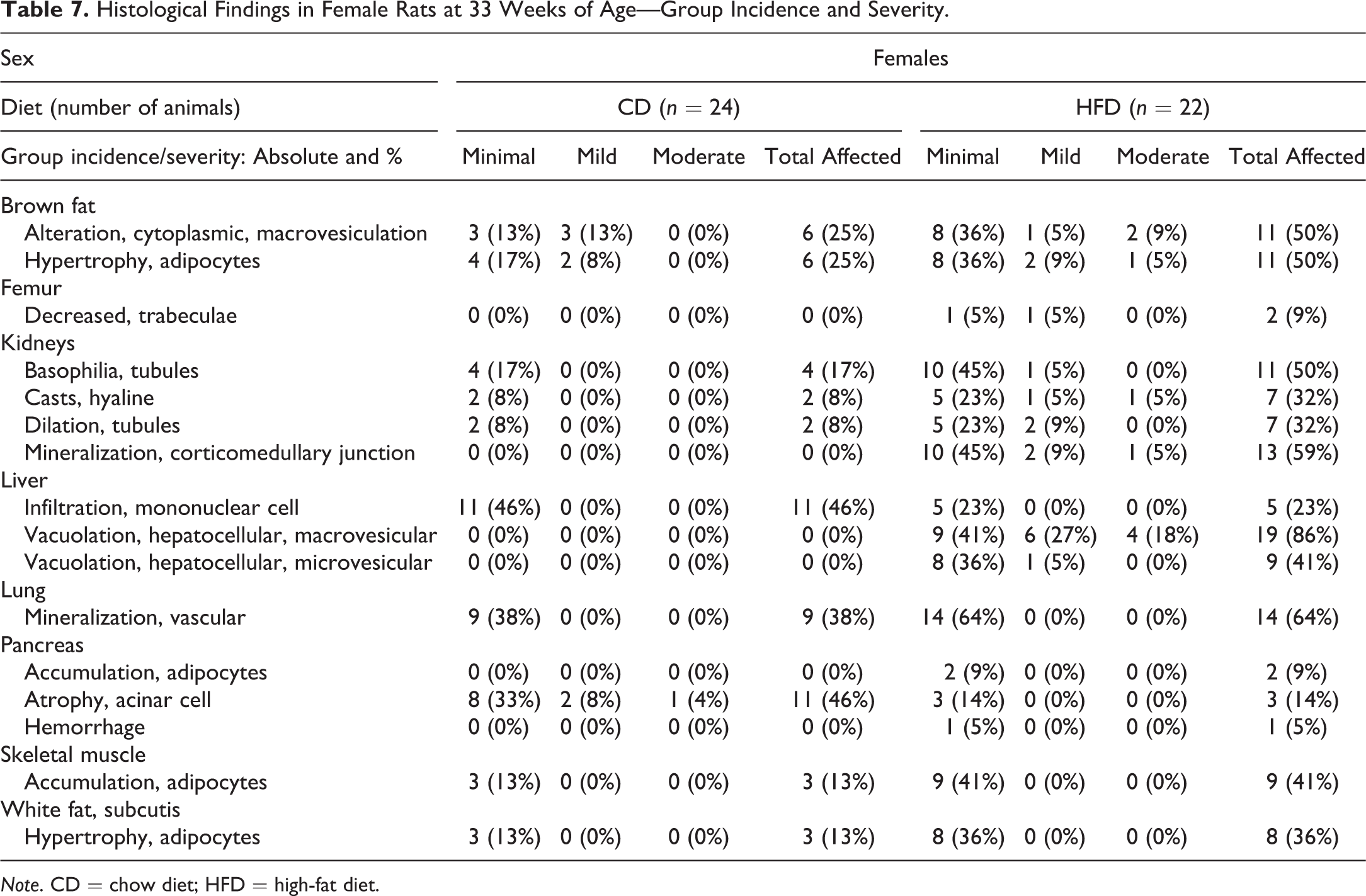
Note. CD = chow diet; HFD = high-fat diet.
Histological Findings in Male Rats at 33 Weeks of Age—Group Incidence and Severity.
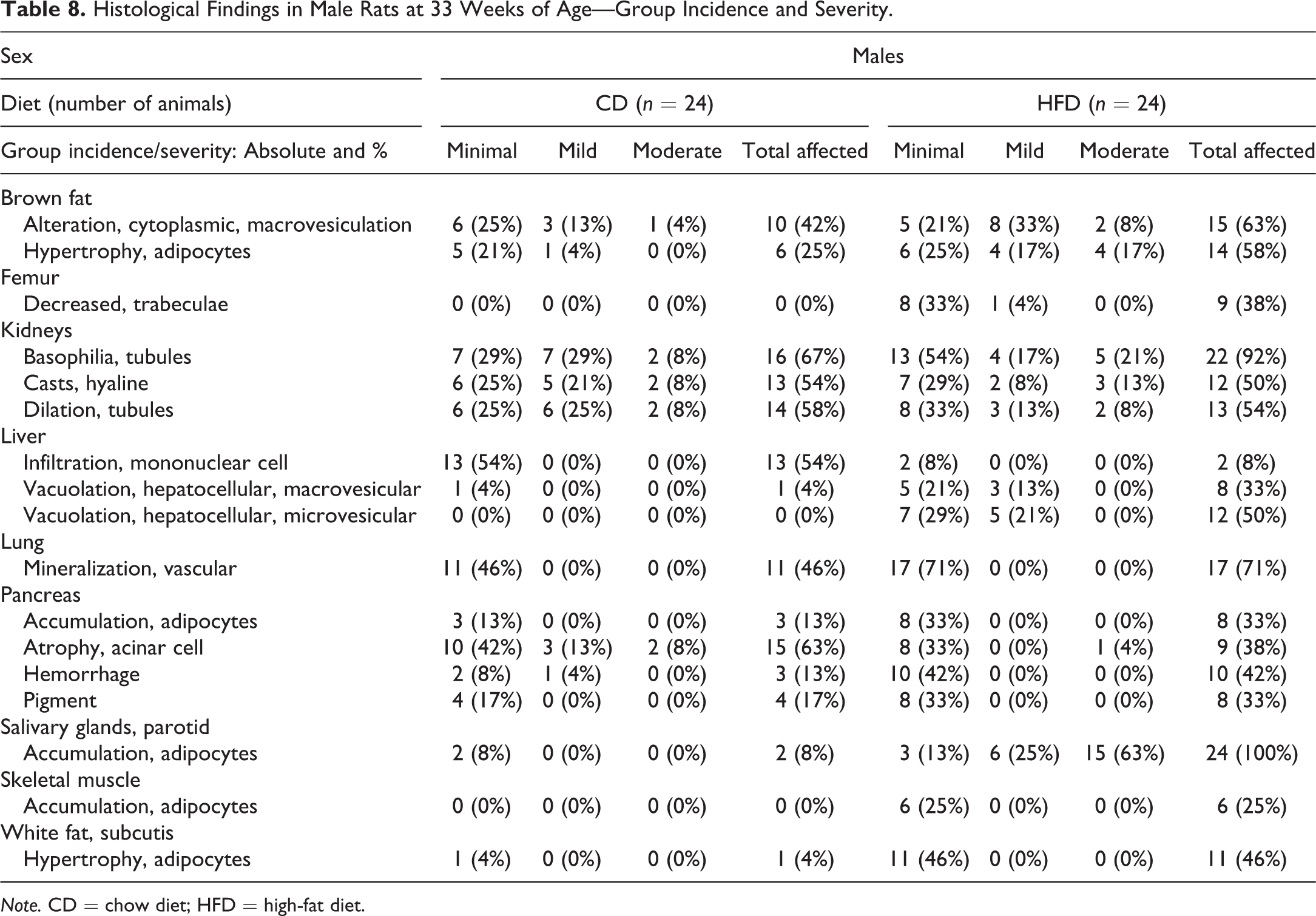
Note. CD = chow diet; HFD = high-fat diet.
HFD-fed rats of both sexes exhibited a higher incidence of cytoplasmic macrovesiculation and adipocyte hypertrophy in the BAT. These findings were mainly of minimal to mild severity, but up to moderate severity was observed in a small percentage of animals (Tables 7 and 8). This is consistent with the observed increase in the weight of this tissue in the HFD-fed animals (Table 1). The brown adipocytes were increased in size with enlargement of the cytoplasmic lipid droplets in the HFD group. Yet the cells still appeared multilocular (Figure 1A, B). Both sexes in the HFD group showed a higher incidence of minimal hypertrophy in the white adipocytes in the subcutaneous inguinal fat pad (Tables 7 and 8). This finding is consistent with the observed increase in the weight of this tissue in these animals (Table 1). Hypertrophy was not found in the two abdominal white fat pads (retroperitoneal and gonadal), in spite of the weight of these compartments being significantly higher in the HFD-fed animals (Table 1 and Online Supplemental Table 6A, B). Neither the brown nor white fat pad depots had differences in inflammatory cell infiltrates between the dietary groups for both sexes (Online Supplemental Table 6A, B). In the HFD group, a higher incidence and/or severity of white adipocyte accumulation occurred in the interstitial tissue in the pancreas and skeletal muscle in both sexes and the parotid salivary glands in the males (Tables 7 and 8). HFD feeding increased the incidence and severity of tubular basophilia in the kidneys in both sexes (Tables 7 and 8). HFD-fed females, but not males, exhibited a higher incidence and severity of kidney tubular dilation with hyaline casts (Tables 7 and 8; Figure 1C, D). These findings were of minimal to moderate severity. Tubular mineralization at the corticomedullary junction, mainly of minimal severity, was present in only the HFD-fed females (Table 7). Macro- and microvesicular hepatocellular vacuolation in the liver was observed in both sexes in the HFD group (Tables 7 and 8; Figure 2A, B). These findings were mainly of minimal to mild severity, but a small percentage of HFD-fed females exhibited moderate severity. The vacuolation was without zonal distribution and primarily observed in the caudate liver lobe. Minimal hepatocellular vacuolation was observed in only a single male in the CD group (Table 8). Oil Red O stain verified that the vacuoles contained neutral lipid droplets (Figure 2B, inset). The incidence of minimal focal mononuclear cell infiltration in the liver parenchyma was lower in both sexes fed the HFD (Tables 7 and 8).
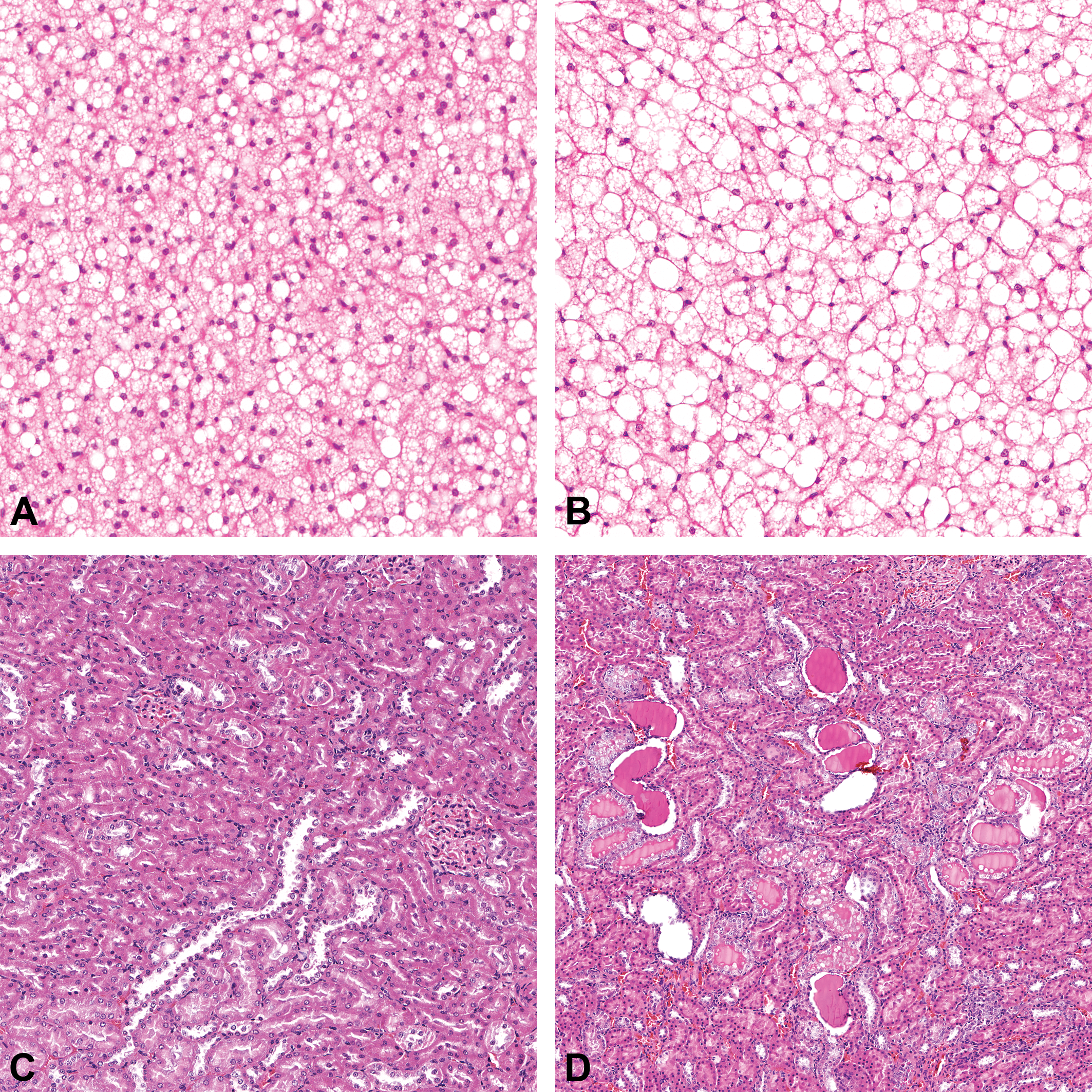
Representative images of histopathological findings in the BAT and kidney in rats fed either a CD or HFD for 29 weeks. (A, B) Micrographs (original objective 20×) of H&E-stained interscapular BAT in the CD-fed rat (A) showing normal multilocular brown adipocytes with small cytoplasmic lipid droplets versus HFD-fed animal (B) which shows that the adipocytes appeared increased in size (hypertrophy) with enlargement of the cytoplasmic lipid droplets (macrovesiculation); the cells still appear multilocular. (C, D) Micrograph of the H&E-stained kidney (original objective 8×) in the CD-fed rat (C) showing normal kidney structure versus a HFD-fed animal (D) showing tubular basophilia and tubular dilation with hyaline casts. BAT = brown adipose tissue; CD = chow diet; H&E = hematoxylin and eosin; HFD = high-fat diet.
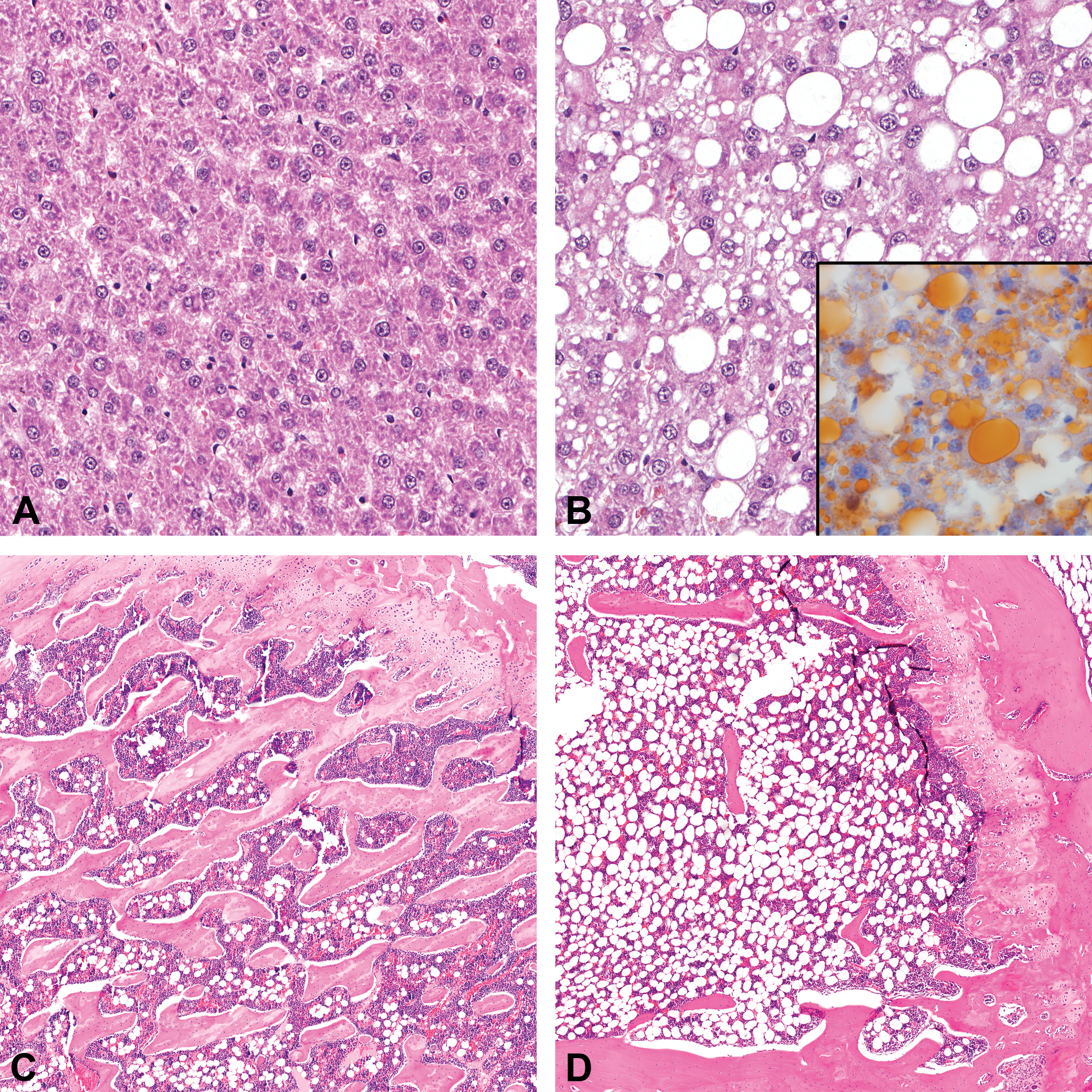
Representative images of histopathological findings in the liver and distal femur bone in 33-week-old CD- versus HFD-fed rats. (A, B) Micrographs of H&E-stained liver tissue (original objective 20×) from a CD-fed rat (A) showing normal hepatocytes with normal lipid content versus a HFD-fed animal (B) showing macro- and microvesicular hepatocellular vacuolation of Lobus Caudatus. Inset: Micrographs of Oil Red O stained vacuoles in the liver (original objective 60×) from a HFD-fed rat shown to contain neutral lipid droplets. (C, D) Micrographs of H&E-stained distal femur bone (original objective 2×) from a CD-fed rat (C) showing normal trabecular pattern versus a HFD-fed animal (D) showing decreased amount of trabeculae. CD = chow diet; H&E = hematoxylin and eosin; HFD = high-fat diet.
Focal vascular mineralization of minimal severity in the lung was observed at higher incidence in both sexes in the HFD group (Tables 7 and 8). HFD-fed males had a higher incidence of minimal hemorrhage and pigment deposition observed in the periphery of the pancreatic islets (Table 8). The pancreatic hemorrhage was observed in only one HFD-fed female (Table 7). A lower incidence and severity of atrophy of the pancreatic acinar cells were observed in the HFD-fed animals (Tables 7 and 8). Both HFD-fed sexes had minimal to mild decreased trabecular bone in the distal femur. These findings were most common in the HFD-fed males and not observed in the CD-fed animals (Tables 7 and 8; Figure 2C, D).
Correlational Analysis of Whole-body Fat Mass and Adipocyte-related Histological Changes
We compared animals with and without adipocyte-related histological findings (i.e., adipocyte hypertrophy and/or accumulation) in the various organs within the same dietary group. These analyses revealed that those HFD-fed rats exhibiting positive findings in brown fat, subcutaneous WAT, and skeletal muscle had higher whole-body fat mass (Figure 3A–H). In the CD-fed rats, only females exhibited an association of higher body fat with the presence of brown fat hypertrophy (Figure 3A). Venn diagrams were created to determine the degree of overlap of adipocyte-related histological findings in multiple organs in the CD- versus HFD-fed animals (Online Supplemental Figure 2A, B, D, and E). Only a few CD-fed rats in both sexes exhibited a histological finding in either two or three organs (Online Supplemental Figures 2A, D). Chronic HFD feeding resulted in the presence of up to four histological changes in the various organs within individual animals (Online Supplemental Figure 2B, E). These were highly correlated to body fat mass (Online Supplemental Figure 2C, F).
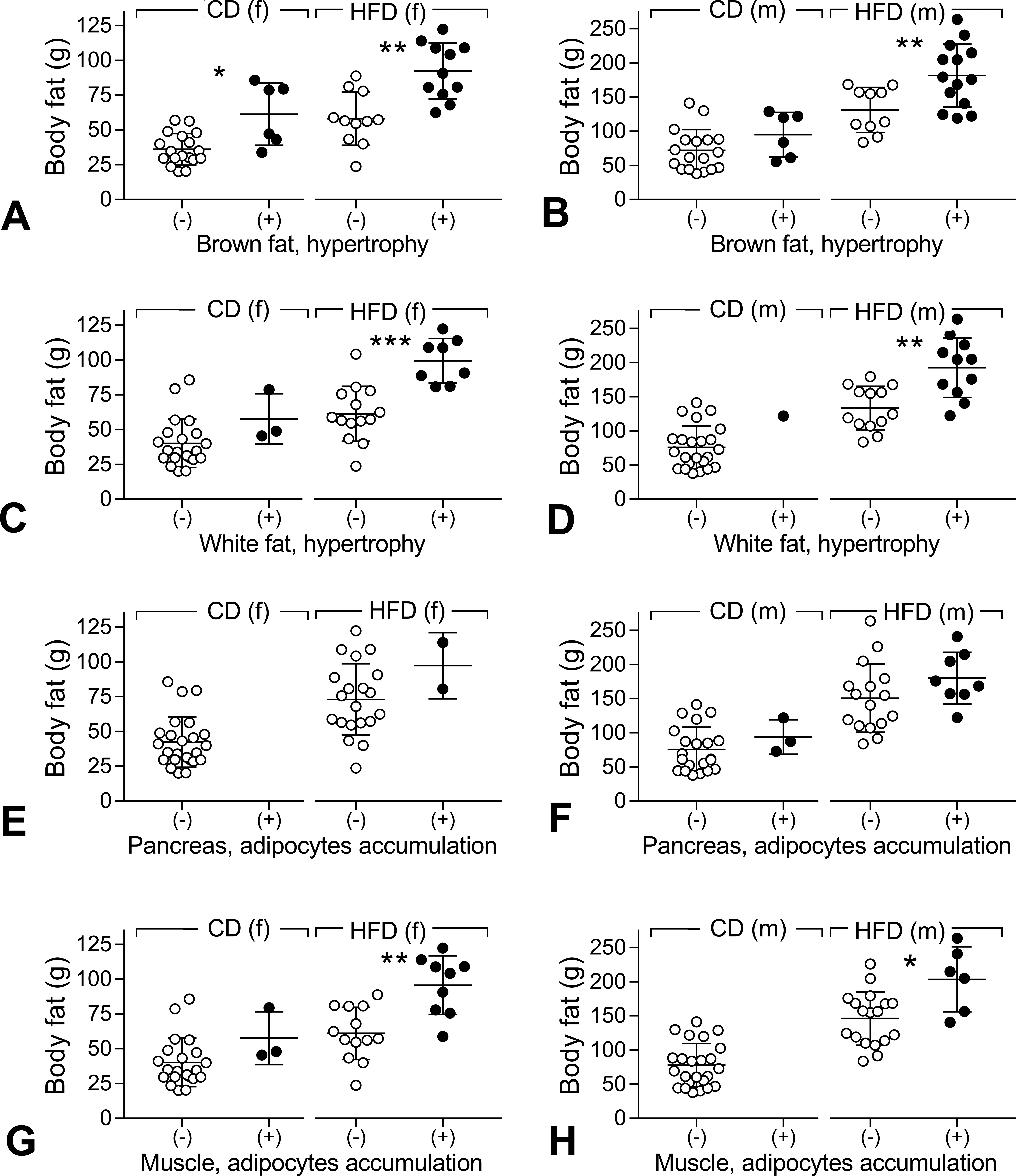
Impact of body adiposity on the incidence of adipocyte hypertrophy and/or accumulation in various organs in CD- versus HFD-fed rats at 33 weeks of age. (A–H) Scatterplot of body fat mass of females (f; left panel) and males (m; right panel) in relation to negative (−) or positive (+) histopathological findings in (A, B) brown fat, (C, D) subcutaneous white fat, (E, F) pancreas, and (G, H) skeletal muscle. Data are the means ± SD (n = 22–24 rats/dietary group). *p < .05. **p < .01. ***p < .001 by Mann-Whitney test (negative [−] vs. positive [+] within each dietary group). CD = chow diet; HFD = high-fat diet; SD = standard deviation.
Reproductive System
Next, we determined the impact of HFD feeding on estrous cycle regularity in female rats. Seventeen-week-old females exhibited regular estrous cycles of 4 to 5 day duration in 71% of CD-fed and 68% of HFD-fed rats (Online Supplemental Figure 3A). The relative numbers of animals with regular estrous cycles were slightly reduced by 30 weeks of age: only 58% of the CD-fed and 50% of the HFD-fed rats maintained a regular estrous cycle length (Online Supplemental Figure 3A). Overall, there were no differences detected in the ratio of regular to irregular cycle duration between the dietary groups (Online Supplemental Figure 3A). Thirty-week-old HFD-fed females exhibiting a higher degree of whole-body fat mass demonstrated irregular estrous cycles in contrast to the females with regular cycles within the same dietary group (Online Supplemental Figure 3C). The amount of body fat in the CD-fed females was similar between animals with either regular or irregular cycles (Online Supplemental Figure 3B, C). Next, we measured reproductive hormones in the plasma at study termination (33 weeks of age) to determine whether chronic HFD feeding adversely impacts these parameters. HFD feeding did not alter reproductive hormone levels in the female rats (Table 9). For the HFD-fed males, circulating levels of FSH and LH were both significantly attenuated by ∼38% (Table 9). Consistent with the lowered gonadotropin levels, HFD feeding induced a tendency for reduced plasma testosterone levels by 29% in the males (Table 9).
Effect of Diet-induced Obesity on Reproductive Hormones in Rats at 33 Weeks of Age.

Note. Data are the means ± SD (n = 22–24 rats/dietary group). CD = chow diet; FSH, follicle-stimulating hormone; HFD = high-fat diet; LH, luteinizing hormone; SD = standard deviation.
*** p < .001.
† p < .0001 (by Student’s t-test or Mann-Whitney test [CD vs. HFD]).
Epiphyseal Growth Plate of the Proximal Tibia Bone and Biomarkers of Bone Turnover
The HFD-fed females exhibited a 10% increase in the total height of the proximal tibia bone growth plate in contrast to the males. This was due to an increase in the proliferative and not the hypertrophic zone of chondrocytes (Online Supplemental Table 7). Circulating levels of IGF-1 were minimally reduced in the males (at all ages) and the females (at 33 weeks of age) in the HFD group (Online Supplemental Table 8). Histomorphometric examination of the metaphysis region directly below the tibia epiphyseal growth plate revealed a significant reduction in the TB VF by 36% in the HFD-fed males (Online Supplemental Table 7). TB VF was reduced by only 8% in the HFD-fed females and did not reach statistical significance. We next investigated highly specific and sensitive markers of bone turnover to determine the underlying cause for the observed trabecular bone loss. The volume fraction of osteoclast associated with the TB VF was not different between the dietary groups for both sexes (Online Supplemental Table 7). CTX-1 levels were minimally attenuated by 10% in the HFD-fed males at 33 weeks of age (Online Supplemental Table 8). Osteocalcin was significantly increased in the plasma of the HFD-fed females at all ages, which did not occur in the male rats (Online Supplemental Table 8).
Discussion
Obesity is associated with an elevated risk of morbidity and mortality owing to the development of obesity associated comorbidities such as cardiovascular disease, diabetes, and cancer (Abdelaal, le Roux, and Docherty 2017; Hruby and Hu 2015). This includes the metabolic syndrome, which is a clinical constellation of cardiovascular risk factors characterized by abdominal (visceral) obesity, insulin resistance ± glucose intolerance, atherogenic dyslipidemia, hypertension, and a prothrombotic and pro-inflammatory profile (Despres and Lemieux 2006; Grundy et al. 2004). Human epidemiological and animal studies indicate that obesity has a detrimental impact on bone and reproductive health, consequently leading to osteoporosis and associated increased risk for bone fracture (Cao 2011) and infertility (Jungheim et al. 2012). Yet there is a substantial proportion of the obese population that do not develop the metabolic abnormalities that commonly accompanies excess adiposity, a condition known as the “metabolically healthy obese” (Phillips 2013; Jung, Lee, and Song 2017; Hamer and Stamatakis 2012).
Genetic inheritance plays a strong contributing role in the etiology of human obesity. Monogenic forms of human obesity specifically caused by a single gene mutation, such as in key energy balance genes (e.g., loss of function mutations in genes encoding leptin, leptin receptor, or pro-opiomelanocortin), result in morbid obesity but occur very rarely in the human population (Barsh, Farooqi, and O’Rahilly 2000). Common human obesity is polygenic in nature and represents a complex interaction between multiple genes and the environment (Barsh, Farooqi, and O’Rahilly 2000). The cause of the obesity crises is the profound obesogenic environmental changes adopted by developed and developing countries. Specifically, this is characterized by the growing surplus availability of inexpensive, calorically dense, and nutrient-poor food coupled with a sedentary lifestyle (Hruby and Hu 2015).
Here we describe, for the first time to our knowledge, a longitudinal study characterizing the consequences of diet-induced obesity in SD rats up to 33 weeks of age. This includes standard toxicology parameters (i.e., clinical pathology and histopathology) and changes in metabolic, endocrine, bone, and reproductive parameters (all of which are commonly perturbed in human obesity). We selected the outbred SD rat model from Taconic Biosciences (NTac: SD) and fed these animals a HFD comprising 45% energy from fat and 17% energy from sucrose which mimics the human Western diet (Cordain et al. 2005) to achieve diet-induced obesity. We did not observe in these rats a clear segregation into a diet-induced susceptible and resistant subpopulation following exposure to a HFD unlike previous reports using the Charles River SD rat strain (Levin et al. 1997; Levin and Dunn-Meynell 2000). Our finding in the current study that the weight gain ranges were normally distributed and did not follow a bimodal segregation is in agreement with the expected distribution of a polygenic trait in the outbred rat population (Archer et al. 2003; Mercer and Archer 2005). Taken together, these key features of the DIO rat model closely mimics the obesity susceptibility of the human population on a Western diet (Blundell and Cooling 2000).
Consistent with previous evidence (Woods et al. 2003; Posey et al. 2009), 29 weeks of HFD feeding led to the manifestation of obesity as evidenced by a modest (8% for females vs. 9% for males) but nonetheless significant increase in body weight. Contrasting reports show that >10% increase in body weight can be achieved in both male and female SD rats obtained from the same vendor (Taconic, Denmark), if provided long-term ad-lib access to a candy supplemented feeding paradigm or a high fructose-HFD (Axelsen et al. 2010; Raun et al. 2007). Nevertheless, there are a myriad of attributing factors that can confound DIO rodent studies (e.g., differences in diet, environmental conditions, strain, and vendor; Kleinert et al. 2018). One confounder of the current toxicology study design that potentially hindered the robust HFD-induced weight gain may be related to the rats undergoing multiple procedures (e.g., blood sampling, estrous cycle measurements, and body composition analysis). Altogether, this limitation must be seriously considered, when designing a toxicology study that includes the use of a DIO rodent model and body weight differences as a primary end point (Kleinert et al. 2018).
However, using body weight as a surrogate measure for ascertaining whether the obese phenotype is achieved, per se, is misleading and can underestimate the degree of obesity developed in the HFD-fed rats (Woods et al. 2003). Specifically, the combined absolute weights of the WAT depots were on average 55% and 92% heavier in the HFD-fed females and males, respectively. These findings are consistent with the 2-fold increase in whole-body fat mass, in contrast to the CD-fed rats. The positive energy balance following chronic HFD feeding in the rats may also be related to the significant reduction in thyroid hormone levels, TSH, and T4. Specifically, the attenuation of these circulating hormones may indicate a disruption of the hypothalamic–pituitary–thyroid axis, which is a key neuroendocrine pathway involved in the stimulation of the basal metabolic rate (Mullur, Liu, and Brent 2014). Contrary to previous publications (Tannenbaum et al. 1997; Everds et al. 2013), chronic HFD feeding did not appear to induce a stress response and the subsequent activation of the hypothalamic–pituitary–adrenal axis. This is supported by evidence that plasma ACTH levels were not increased in both sexes.
The HFD-fed rats developed hyperleptinemia and hyperinsulinemia, and these hormonal levels positively correlated with the degree of whole-body fat mass and white fat pad depots. These findings are similar to those observed in obese humans (Considine et al. 1996; Bagdade, Bierman, and Porte 1967) and rodents (Posey et al. 2009; Woods et al. 2003), suggesting the development of systemic resistance to these hormones in the setting of long-term HFD feeding. Moreover, the observation in this study that the degree of insulin resistance (HOMA-IR) and adiposity is positively correlated agrees with previous evidence that excess adiposity plays a significant role in the development of insulin resistance in humans (Garg 2004) and rodents (Gao, Ma, and Liu 2015; Posey et al. 2009). Yet the HFD-fed animals never developed diabetes. This is consistent with previous findings in DIO SD rats and attributed to compensatory increases in both pancreatic β-cell mass and glucose-stimulated insulin secretion (Raun et al. 2007; Marques et al. 2016; Madsen et al. 2010). Circulating levels of adiponectin, which increases insulin sensitivity, were found to be significantly higher in the HFD-fed males and agree with previous studies in DIO rodents (Marques et al. 2016; Patsch et al. 2011; Raun et al. 2007). These findings contradict the well-documented decrease observed in obese humans (Kern et al. 2003). It is conceivable that the insulin sensitizing effect of adiponectin may be impaired in the setting of obesity owing to the development of “adiponectin resistance” (Scheid and Sweeney 2014).
HFD-fed females were found to exhibit a mild elevation in plasma TG levels, consistent with a previous report (Aubin et al. 2008), and were coupled with a minor reduction in TC levels. The positive correlation of TG levels with HOMA-IR in the females suggests that the mild hypertriglyceridemia observed in the HFD group may be driven by the insulin-resistant state (Ginsberg 2000). HFD-fed males had transient and minor fluctuations in plasma lipid levels relative to CD-fed rats and had no diet-related differences by 33 weeks of age. This corroborates the study by Marques et al. (2016) who reported similar findings in SD male rats fed a 45% HFD for 17 weeks. Yet chronic HFD feeding did not induce overt dyslipidemia, especially in the males (which are the most susceptible [Palmisano et al. 2018]), in contrast to other studies (Axelsen et al. 2010; Madsen et al. 2010). These divergent outcomes may be explained by not only dietary differences but also an age-related effect—that is, whereby lipid levels were also observed to steadily increase with advancing age in the CD-fed rats. This can confound significant differences between the dietary groups and is a consequence of the ad-lib overfeeding state in normal control animals (Keenan et al. 2005; Martin et al. 2010). Altogether, these data demonstrate that 29 weeks of feeding a HFD induce obesity in both sexes and are associated with metabolic derangements (hyperleptinemia, hyperinsulinemia, insulin resistance, and mild hypertriglyceridemia [only females]).
The changes in blood hematology, clinical chemistry, and urinalysis parameters were minor, and several alterations were considered related to differences in the macro- and micro-nutrient composition of the CD versus HFD. Specifically, the observed significant decrease in circulating Mg levels in the HFD group throughout the study is attributed, in part to the lower content of this micronutrient in the HFD (∼0.06%) with respect to the CD (0.20%). Factoring in the 27% reduction in average daily food intake (g) in the HFD-fed versus the CD-fed animals (data not shown) indicates that daily Mg intake was 78% less for the HFD group. Likewise, the 2-fold increase in circulating levels of SDH by 20 and 33 weeks of age in the HFD-fed animals may also be diet related. The HFD is enriched in sucrose (unlike the CD), which can provide the glucose substrate to fuel SDH activity. Intracellular SDH facilitates the process of de novo lipogenesis by using the glucose-derived substrate sorbitol to synthesize endogenous fructose (via the polyol pathway). This rapidly depletes ATP and consequently blocks fatty acid oxidation in the liver (Jensen et al. 2018). There was a corresponding reduction in circulating levels of the ketone body, β-HB (an important byproduct of β-oxidation) in HFD-fed animals. This latter finding may reflect an influence on mitochondrial function and consequent decrease in hepatic fatty acid oxidation (Chao et al. 2014).
The urine of both sexes in the HFD group was more acidic in contrast to the alkaline urine of the CD-fed animals, which reflect dietary differences. The CD is cereal-based (which lacks animal protein), whereas the HFD-fed animals consumed a diet enriched in casein (milk protein), resulting in a lower urinary pH (Breslau et al. 1988). Urinary crystals, urates, epithelial cells, and bacteria were observed at lower incidence in the HFD group and also considered to be attributed to differences in urinary pH and diet (Whalan 2015).
Obesity is associated with a hypercoagulable state and increased risk for thrombotic disorders (Kaji et al. 2013; Darvall et al. 2007; De Pergola and Pannacciulli 2002). APTT and PT were marginally shorter in the HFD-fed animals for both sexes. Contrary to previous studies (Kaji et al. 2013; El-Gendy and Abbas 2014; Dallak et al. 2018; Sakr, Abbas, and Haidara 2018), HFD-fed rats exhibited a decrease in both FIB and PLT counts, which may reflect a low-grade consumption of both parameters owing to a mild hypercoagulable state associated with chronic HFD feeding. These findings suggest that the HFD-fed animals may have a higher tendency for increased blood clotting. However, the changes in the coagulation parameters were considered to be minor, and microscopic histological examination of all organs did not reveal any signs of thrombus formation. Collectively, HFD-fed rats exhibited small alterations in several clinical pathology parameters which are most likely related to dietary differences and not associated with any organ/tissue damage.
HFD feeding in the males led to a decrease in both absolute and adjusted epididymides weight, which may indicate a decrease in sperm content (Creasy et al. 2012). There were no correlating histological findings to this weight finding. However, tendencies toward lower circulating testosterone levels were observed due to the reduction in the gonadotrophic hormones, FSH, and LH, which corroborates previous findings (Olivares et al. 2010). This finding is consistent with a previous report (Vidal and Whitney 2014), which shows that compounds that lower testicular testosterone result in the attenuation of the epididymal weight. A similar trend was not observed in the prostate and the seminal vesicles. An observed decrease in the adjusted weights of the spleen, liver, salivary glands (both sexes), and brain (females) without any differences in the absolute weights may merely reflect a lack of correlation between the organ weight and the body weight in the HFD-fed animals.
Microscopic examination of BAT and subcutaneous WAT revealed a higher incidence of adipocyte hypertrophy in the HFD-fed animals. These findings were consistent with the observed increase in the fat pad tissue weights and were highly correlated with the degree of whole-body fat mass. The macrovesicular appearance of the brown adipocytes in the HFD group is in accordance with previous publications (Shimizu et al. 2014; Betz and Enerback 2011) and may reflect reduced thermogenic activity within this tissue. It is plausible that this is related to a decrease in the activity of the mitochondrial uncoupling protein 1.
In response to the excessive caloric intake and the consequent weight gain induced by long-term HFD feeding, white fat pad depots expand via hyperplasia, hypertrophy, or both (Chusyd et al. 2016; Cinti 2012), which function as a metabolic sink to buffer the energy surplus (Frayn 2002). Our finding of hypertrophy in the subcutaneous WAT may be related to a lower metabolic activity in this tissue as previously reported (Sackmann-Sala et al. 2012). WAT depot expansion associated with aging or HFD feeding in rodents is indicative of a more insulin-resistant tissue and concomitant-reduced capacity as a metabolic sink and a marker of increased risk for developing type 2 diabetes (Frayn 2002; Sierra Rojas et al. 2016; Gao, Ma, and Liu 2015). Although it is well-documented that HFD feeding in rats leads to inflammation in the white fat depots (Cinti 2012; Hill, Reid Bolus, and Hasty 2014), there were no differences in inflammatory cell infiltrates in the adipose tissues between the dietary groups.
Ectopic lipid deposition in the various nonadipose organs, which commonly occurs in the setting of obesity (Snel et al. 2012; Gao, Ma, and Liu 2015), was found to occur at a higher incidence in the HFD-fed animals. Specifically, intercellular adipocyte accumulation was observed in the pancreas, skeletal muscle, and parotid salivary glands, and intracellular lipid accumulation was observed in the liver, ranging from minimal to moderate severity. The HFD-fed animals with higher body fat mass exhibited adipocyte-related histological findings (adipocyte hypertrophy and/or accumulation) in multiple organs (i.e., BAT, subcutaneous WAT, and muscle). Taken together, our findings agree with previous reports that demonstrate that the degree of adiposity correlates with adipocyte size and ectopic fat deposition (van Harmelen et al. 2003; Sinha et al. 2002).
Microscopic examination of the liver revealed minimal to mild hepatocellular vacuolation for both sexes in the HFD group. This was not accompanied by cell damage in hepatocytes, bile duct epithelium, or inflammatory cell infiltration. The liver enzymes ALAT and ASAT that are abnormally altered in the setting of liver damage (Whalan 2015; Giannini, Testa, and Savarino 2005; McDyre et al. 2018) were not increased in both sexes in the HFD-fed rats. This is consistent with a previous report in the DIO SD rat strain (Marques et al. 2016). This raises the likelihood that the observed minor increases in total bilirubin and SDH levels in the HFD-fed animals may not represent a marker of potential liver injury/cholestasis. Contrasting previous reports show that HFD feeding in rodents can lead to lipid droplets within the periportal zone (Tetri et al. 2008; Sampey et al. 2011), while the present study did not find any zonal distribution of lipid droplets in the liver. This fatty change was more pronounced in the caudate liver lobe. Hence, this may be related to inherent differences between the various liver lobes with respect to the distribution of the hepatic artery and portal vein branches that supply nutrient-rich blood (Malarkey et al. 2005) and the heterogeneous distribution of metabolic enzymes (Matsubara, Touchi, and Ogawa 1982). Liver lipid accumulation in the HFD-fed animals did not correlate with the degree of whole-body fat mass (data not shown), suggesting that the underlying cause may be related to other factors (e.g., diet, metabolic changes), independent of obesity. Both sexes fed the HFD exhibited a lower incidence of minimal focal mononuclear cell infiltration in the liver parenchyma. The underlying cause for this is unknown, but the finding is a common background finding, especially in older rats and may be incidental (Suttie, Leininger, and Bradley 2018).
Both sexes in the HFD group exhibited increased incidence of minimal to moderate tubular changes in the kidneys (tubular basophilia and/or tubular dilation with hyaline casts). These findings indicate that long-term HFD feeding increases the workload on the kidney. This agrees with a previous report (Altunkaynak et al. 2008), which revealed that SD rats fed HFD for 3 months developed histopathological changes in the kidney; albeit, these findings were more severe than observed in the current study. HFD-fed females exhibited tubular mineralization at the corticomedullary junction. Female rats are especially sensitive for kidney mineralization, which occurs as a spontaneous lesion (Greaves 2012). This kidney finding may be associated with the attenuated circulating levels of Mg secondary to lower dietary intake of this micronutrient (Massy and Drueke 2012; Reeves, Nielsen, and Fahey 1993). A higher incidence of minimal focal vascular mineralization was observed in the lung in both sexes in the HFD group. Although this is a very common background finding in rats (McInnes 2011), it is currently speculated that the higher occurrence in the HFD-fed rats may be similarly related to reduced Mg levels (Nicoll, Howard, and Henein 2015) as the kidney mineralization findings.
HFD-fed males exhibited a higher incidence of hemorrhage and pigment deposition within the periphery of the pancreatic islets in contrast to the CD-fed controls. These alterations in the pancreatic islets have also been observed in aging male rats (Imaoka, Satoh, and Furuhama 2007). Furthermore, the study by Imaoka, Satoh, and Furuhama (2007) demonstrates that pancreatic islet hemorrhage occurred at a higher incidence in male rats with lower levels of circulating estrogen. However, these authors reported in a later study (Imaoka et al. 2009) that treating aging male SD rats with weekly subcutaneous estradiol for 20 weeks did not inhibit the occurrence of these hemorrhagic lesions. Furthermore, feeding aging male rats a chronic high fat/protein diet versus the standard CD did not impact the onset and character of the pancreatic islet alterations (Imaoka, Satoh, and Furuhama 2007). We conclude that the higher occurrence of pancreatic islet hemorrhage in the HFD-fed male rats may be incidental.
Microscopic and histomorphometric examination of the distal femur and the proximal tibia bone, respectively, revealed that the trabecular bone was decreased in both sexes in the HFD group and at a higher incidence in the males. Our findings contradict the traditional view that obese individuals are protected against age-related bone loss owing to increased mechanical loading conferred by the higher body weight (otherwise known as the Mechanostat theory; Reid 2002; Shapses and Sukumar 2012; Iwaniec and Turner 2016). In both humans (Gower and Casazza 2013) and rodents (Gautam et al. 2014; Patsch et al. 2011; Cao, Gregoire, and Gao 2009; Cao and Picklo 2015; Dong et al. 2016; Inzana et al. 2013), excessive fat mass can impair the ability of the skeleton to fully adapt to the increased mechanical load. Consequently, this leads to detrimental effects on bone mass, as well as the structural and mechanical properties of the cortical and trabecular bone. Whether HFD feeding adversely impacts the former and latter bone parameters cannot be ascertained from the routine light microscopy evaluation performed in this study (which is the standard procedure used in regulatory toxicity studies). This requires the employment of highly sensitive ex vivo bone scanning techniques (e.g., microcomputed tomography) together with biomechanical testing (Smith, Varela, and Samadfam 2017) and awaits further study.
Trabecular bone loss induced by HFD feeding may be attributed to reduced bone formation, increased resorption, or a combination thereof (Gautam et al. 2014; Patsch et al. 2011; Cao, Gregoire, and Gao 2009). Our investigation found that the volume of CD-68 positive osteoclast associated with the trabecular bone within the metaphysis region of the proximal tibia was not different in the HFD-fed animals. Additionally, there were no changes in circulating CTX-1 levels, a marker of osteoclast activity, which is released during the resorption of the bone matrix (Smith, Varela, and Samadfam 2017). These findings suggest that osteoclast-mediated resorption cannot account for the trabecular bone loss. It can be speculated that osteoblast-mediated bone formation must be reduced. However, this bone turnover parameter could not be measured in the present study owing to technical issues and awaits further investigation. The HFD-fed females (unlike the males) exhibited higher circulating levels of osteocalcin, a highly sensitive marker of bone formation and osteoblast number (Smith, Varela, and Samadfam 2017). This finding raises the likelihood that osteoblast activity may be higher in these animals and thus potentially offset the negative effects of HFD feeding on bone. This results in less pronounced trabecular bone loss in the female rats, although this warrants further study.
Several mechanisms have been proposed that would account for the negative impact of obesity on bone. This includes a decrease in muscle mass and function, reduced physical activity, and deficiencies in essential bone regulatory hormones and nutrients (Shapses and Sukumar 2012; Shapses and Cifuentes 2015; Iwaniec and Turner 2016). Albeit lean muscle mass was significantly reduced in the HFD-fed males, spontaneous locomotor activity was not measured in the animals to assess for impaired physical activity and therefore is a caveat of the present study. Ex vivo biomechanical testing on bone can be used as a surrogate measure to ascertain whether bone strength and fracture resistance were undermined by HFD-induced obesity in the rats (Smith, Varela, and Samadfam 2017) and awaits further investigation. Perturbations in calcium and phosphorus homeostasis can negatively impact bone health (Smith, Varela, and Samadfam 2017). However, circulating levels of these essential minerals were not different between the dietary groups and therefore are not considered underlying factors. The observed (and sex specific) deleterious changes in the pituitary/thyroid (FSH, LH, ACTH, TSH, and T4), gonadal (testosterone), metabolic (insulin and IGF-1, a growth factor involved in linear bone growth and remodeling, [Kawai and Rosen 2010; Yakar et al. 2002]), and adipocyte-derived hormones (leptin, adiponectin) in the HFD-fed animals may serve as important contributing factors to the trabecular bone loss. These aforementioned bone-regulatory hormones are intimately associated with skeletal growth, maturation, and turnover (Fernandes, Goncalves, and Brito 2017; Cao 2011; Iwaniec and Turner 2016). Therefore, this is a hypothesis that should be tested in future studies.
Conclusion
We herein provide a comprehensive set of toxicology data in the DIO SD rat model that indicates the applicability of this model for future toxicity studies for two reasons: (1) chronic HFD feeding in the polygenic SD rat model led to the development of an obese phenotype and associated metabolic abnormalities that mimics, to a certain extent common human obesity; and (2) the histological examination reveals that these HFD-fed animals developed only mild obesity-associated disease manifestations and spontaneous background findings, comparable to that of the normal control animals. Collectively, our data suggest that the DIO SD rats are obese but otherwise generally healthy. These findings raise the possibility that the HFD-induced obese phenotype in the SD rat model most likely resembles metabolically healthy obese humans who virtually lack the development of obesity associated comorbidities (Phillips 2013; Jung, Lee, and Song 2017; Hamer and Stamatakis 2012). Therefore, our data highlight the usefulness of the DIO rat model as a highly valuable adjunct to the conventional healthy animal model in the safety evaluation of weight-loss drugs in support of phase I and II clinical trials (which typically enroll metabolically healthy obese and overweight human subjects; U.S. Department of Health and Human Services. Food and Drug Administration, Center for Drug Evaluation and Research 2007). Furthermore, employing DIO rodents in the toxicity testing of anti-obesity compounds may also help better delineate the exaggerated pharmacology-related findings associated with weight loss from the true drug-related toxicities (Everds et al. 2013). Taken together, we conclude that the DIO SD rat model that mimics the human obese condition may potentially bridge the gap of the unmet need for improving predictability and understanding of toxicities encountered with anti-obesity compounds (Morgan et al. 2013).
Supplemental Material
Supplemental Material, DS1_TPX_10.1177_0192623318803557 - The Effect of Diet-induced Obesity on Toxicological Parameters in the Polygenic Sprague-Dawley Rat Model
Supplemental Material, DS1_TPX_10.1177_0192623318803557 for The Effect of Diet-induced Obesity on Toxicological Parameters in the Polygenic Sprague-Dawley Rat Model by Jennifer M. Rojas, Florian Bolze, Inger Thorup, Jette Nowak, Charlotte M. Dalsgaard, Mikala Skydsgaard, Line O. Berthelsen, Kevin A. Keane, Henrik Søeborg, Ingrid Sjögren, Jes T. Jensen, Johannes J. Fels, Hanne K. Offenberg, Lærke W. Andersen, and Majken Dalgaard in Toxicologic Pathology
Supplemental Material
Supplemental Material, DS2_TPX_10.1177_0192623318803557 - The Effect of Diet-induced Obesity on Toxicological Parameters in the Polygenic Sprague-Dawley Rat Model
Supplemental Material, DS2_TPX_10.1177_0192623318803557 for The Effect of Diet-induced Obesity on Toxicological Parameters in the Polygenic Sprague-Dawley Rat Model by Jennifer M. Rojas, Florian Bolze, Inger Thorup, Jette Nowak, Charlotte M. Dalsgaard, Mikala Skydsgaard, Line O. Berthelsen, Kevin A. Keane, Henrik Søeborg, Ingrid Sjögren, Jes T. Jensen, Johannes J. Fels, Hanne K. Offenberg, Lærke W. Andersen, and Majken Dalgaard in Toxicologic Pathology
Supplemental Material
Supplemental Material, DS3_TPX_10.1177_0192623318803557 - The Effect of Diet-induced Obesity on Toxicological Parameters in the Polygenic Sprague-Dawley Rat Model
Supplemental Material, DS3_TPX_10.1177_0192623318803557 for The Effect of Diet-induced Obesity on Toxicological Parameters in the Polygenic Sprague-Dawley Rat Model by Jennifer M. Rojas, Florian Bolze, Inger Thorup, Jette Nowak, Charlotte M. Dalsgaard, Mikala Skydsgaard, Line O. Berthelsen, Kevin A. Keane, Henrik Søeborg, Ingrid Sjögren, Jes T. Jensen, Johannes J. Fels, Hanne K. Offenberg, Lærke W. Andersen, and Majken Dalgaard in Toxicologic Pathology
Supplemental Material
Supplemental Material, DS4_TPX_10.1177_0192623318803557 - The Effect of Diet-induced Obesity on Toxicological Parameters in the Polygenic Sprague-Dawley Rat Model
Supplemental Material, DS4_TPX_10.1177_0192623318803557 for The Effect of Diet-induced Obesity on Toxicological Parameters in the Polygenic Sprague-Dawley Rat Model by Jennifer M. Rojas, Florian Bolze, Inger Thorup, Jette Nowak, Charlotte M. Dalsgaard, Mikala Skydsgaard, Line O. Berthelsen, Kevin A. Keane, Henrik Søeborg, Ingrid Sjögren, Jes T. Jensen, Johannes J. Fels, Hanne K. Offenberg, Lærke W. Andersen, and Majken Dalgaard in Toxicologic Pathology
Supplemental Material
Supplemental Material, DS5_TPX_10.1177_0192623318803557 - The Effect of Diet-induced Obesity on Toxicological Parameters in the Polygenic Sprague-Dawley Rat Model
Supplemental Material, DS5_TPX_10.1177_0192623318803557 for The Effect of Diet-induced Obesity on Toxicological Parameters in the Polygenic Sprague-Dawley Rat Model by Jennifer M. Rojas, Florian Bolze, Inger Thorup, Jette Nowak, Charlotte M. Dalsgaard, Mikala Skydsgaard, Line O. Berthelsen, Kevin A. Keane, Henrik Søeborg, Ingrid Sjögren, Jes T. Jensen, Johannes J. Fels, Hanne K. Offenberg, Lærke W. Andersen, and Majken Dalgaard in Toxicologic Pathology
Supplemental Material
Supplemental Material, DS6_TPX_10.1177_0192623318803557 - The Effect of Diet-induced Obesity on Toxicological Parameters in the Polygenic Sprague-Dawley Rat Model
Supplemental Material, DS6_TPX_10.1177_0192623318803557 for The Effect of Diet-induced Obesity on Toxicological Parameters in the Polygenic Sprague-Dawley Rat Model by Jennifer M. Rojas, Florian Bolze, Inger Thorup, Jette Nowak, Charlotte M. Dalsgaard, Mikala Skydsgaard, Line O. Berthelsen, Kevin A. Keane, Henrik Søeborg, Ingrid Sjögren, Jes T. Jensen, Johannes J. Fels, Hanne K. Offenberg, Lærke W. Andersen, and Majken Dalgaard in Toxicologic Pathology
Supplemental Material
Supplemental Material, DS7_TPX_10.1177_0192623318803557 - The Effect of Diet-induced Obesity on Toxicological Parameters in the Polygenic Sprague-Dawley Rat Model
Supplemental Material, DS7_TPX_10.1177_0192623318803557 for The Effect of Diet-induced Obesity on Toxicological Parameters in the Polygenic Sprague-Dawley Rat Model by Jennifer M. Rojas, Florian Bolze, Inger Thorup, Jette Nowak, Charlotte M. Dalsgaard, Mikala Skydsgaard, Line O. Berthelsen, Kevin A. Keane, Henrik Søeborg, Ingrid Sjögren, Jes T. Jensen, Johannes J. Fels, Hanne K. Offenberg, Lærke W. Andersen, and Majken Dalgaard in Toxicologic Pathology
Supplemental Material
Supplemental Material, DS8_TPX_10.1177_0192623318803557 - The Effect of Diet-induced Obesity on Toxicological Parameters in the Polygenic Sprague-Dawley Rat Model
Supplemental Material, DS8_TPX_10.1177_0192623318803557 for The Effect of Diet-induced Obesity on Toxicological Parameters in the Polygenic Sprague-Dawley Rat Model by Jennifer M. Rojas, Florian Bolze, Inger Thorup, Jette Nowak, Charlotte M. Dalsgaard, Mikala Skydsgaard, Line O. Berthelsen, Kevin A. Keane, Henrik Søeborg, Ingrid Sjögren, Jes T. Jensen, Johannes J. Fels, Hanne K. Offenberg, Lærke W. Andersen, and Majken Dalgaard in Toxicologic Pathology
Supplemental Material
rojas_supplemental_fig1 - The Effect of Diet-induced Obesity on Toxicological Parameters in the Polygenic Sprague-Dawley Rat Model
rojas_supplemental_fig1 for The Effect of Diet-induced Obesity on Toxicological Parameters in the Polygenic Sprague-Dawley Rat Model by Jennifer M. Rojas, Florian Bolze, Inger Thorup, Jette Nowak, Charlotte M. Dalsgaard, Mikala Skydsgaard, Line O. Berthelsen, Kevin A. Keane, Henrik Søeborg, Ingrid Sjögren, Jes T. Jensen, Johannes J. Fels, Hanne K. Offenberg, Lærke W. Andersen, and Majken Dalgaard in Toxicologic Pathology
Supplemental Material
rojas_supplemental_fig2 - The Effect of Diet-induced Obesity on Toxicological Parameters in the Polygenic Sprague-Dawley Rat Model
rojas_supplemental_fig2 for The Effect of Diet-induced Obesity on Toxicological Parameters in the Polygenic Sprague-Dawley Rat Model by Jennifer M. Rojas, Florian Bolze, Inger Thorup, Jette Nowak, Charlotte M. Dalsgaard, Mikala Skydsgaard, Line O. Berthelsen, Kevin A. Keane, Henrik Søeborg, Ingrid Sjögren, Jes T. Jensen, Johannes J. Fels, Hanne K. Offenberg, Lærke W. Andersen, and Majken Dalgaard in Toxicologic Pathology
Supplemental Material
rojas_supplemental_fig3 - The Effect of Diet-induced Obesity on Toxicological Parameters in the Polygenic Sprague-Dawley Rat Model
rojas_supplemental_fig3 for The Effect of Diet-induced Obesity on Toxicological Parameters in the Polygenic Sprague-Dawley Rat Model by Jennifer M. Rojas, Florian Bolze, Inger Thorup, Jette Nowak, Charlotte M. Dalsgaard, Mikala Skydsgaard, Line O. Berthelsen, Kevin A. Keane, Henrik Søeborg, Ingrid Sjögren, Jes T. Jensen, Johannes J. Fels, Hanne K. Offenberg, Lærke W. Andersen, and Majken Dalgaard in Toxicologic Pathology
Footnotes
Acknowledgments
The authors would like to thank Kenneth S. Nielsen, Leon Jacobsen, Louise Nielsen, Helle Wagner, Nikolai K. Jensen, Joan H. Rasmussen, Anette H. Mast, and Dorte Larsen for their expert technical assistance during the in-life study phase, necropsy, and the histology processing and analysis. The authors would also like to thank Brian Lauritzen for providing insightful and constructive feedback on the manuscript. Finally, the authors gratefully acknowledge Hanne Paltoft from CiToxLAB Scantox A/S for providing expert technical assistance in the clinical pathology analysis.
Author Contributions
Authors contributed to conception or design (JR, LA, MD); data acquisition, analysis, or interpretation (JR, FB, IT, JN, CD, MS, LB, KK, HS, IS, JJ, JF, HO, LA, MD); drafting the manuscript (JR, FB); and critically revising the manuscript (JR, FB, IT, JN, CD, MS, LB, KK, HS, IS, JJ, JF, HO, LA, MD). All authors gave final approval and agreed to be accountable for all aspects of work in ensuring that questions relating to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Declaration of Conflicting Interests
The author(s) declared no potential, real, or perceived conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publications of this article: This study was funded by the Novo Nordisk R&D STAR Programme (JR, FB) and by Novo Nordisk A/S.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.