
Abstract
In this study we investigated the distribution of a recently cloned polyspecific organic anion transporting polypeptide (Oatp2) in rat brain by nonradioactive in situ hybridization histochemistry and immunofluorescence microscopy. The results demonstrate that Oatp 2 is expressed in brain capillary and in plexus epithelial cells. At the blood-brain barrier (BBB), Oatp 2 expression could be co-localized with the endothelial marker vWF (von Willebrand factor) but not with the astrocyte marker GFAP (glial fibrillary acidic protein). In choroid plexus epithelial cells, Oatp 2 could be localized to the basolateral cell pole, whereas the first member of the Oatp gene family of membrane transporters to be cloned (Oatp 1) co-localized with the α1subunit of Na,K-ATPase at the apical plasma membrane domain. Because Oatp 1 and Oatp 2 have been previously shown to mediate transmembrane transport of a wide variety of amphipathic organic compounds, including many drugs and other xenobiotics, the histochemical localization of Oatp 2 at the BBB and of Oatp 1 and Oatp 2 in the choroid plexus imply a role for these transporters in the active exchange of amphipathic solutes between the blood, brain, and cerebrospinal fluid compartments.
T
Recently, a novel multispecific organic anion transporting polypeptide (Oatp2) with an exceptionally high affinity for digoxin (Km ~0.24 μM) has been cloned from rat brain (Noé et al. 1997). Oatp 2 belogns to a growing family of multispecific drug transporters that are also expressed in liver and kidney, indicating that they play an important role in overall drug disposition (Meier et al. 1997). The best-characterized member of the Oatp gene family of membrane transporters is Oatp 1, which represents an ∼80-kD protein (Bergwerk et al. 1996), accepts a broad range of amphipathic substrates, including bile acids, steroid and glutathione conjugates, organic anionic dyes, leukotriene C4, GSH, peptidomimetic drugs, and even certain organic cations (Meier et al. 1997; Li et al. 1998), functions as an X− /HCO3 − and/or X− /glutathione (GSH) antiporter (Satlin et al. 1997; Li et al. 1998), and is localized at the basolateral plasma membrane domain of hepatocytes (Bergwerk et al. 1996), the brush border of kidney S3 tubular epithelial cells (Bergwerk et al. 1996), and the apical plasma membrane of the choroid plexus (Angeletti et al. 1997). In contrast, the exact cellular localization of Oatp 2 has not yet been determined. This study demonstrates that Oatp 2 is expressed at BBB endothelial cells and at the basolateral pole of choroid plexus epithelial cells, indicating that it may participate in the active exchange of drugs (e.g., digoxin) between the brain interstitium, the cerebrospinal fluid, and the blood plasma compartments.
Materials and Methods
Animals
Male Sprague-Dawley rats (SUT:SDT) weighing 200-250 g were obtained from the Institute für Labortierkunde, University of Zurich (Zurich, Switzerland) and kept under standard conditions.
Riboprobes
Full-length antisense and sense Oatp 2 cRNAs were labeled with digoxigenin. The 3.6-kb long probes were transcribed in vitro from an Oatp 2 cDNA ligated into the Uni-ZAP XR vector (Noé et al. 1997). The vector was first linearized with restriction enzymes and transcribed with T7 or T3 RNA polymerases for antisense and sense probes, respectively. The transcription process was carried out with the nonradioactive RNA labeling kit (Boehringer; Mannheim, Germany) according to the manufacturer's recommendations. The transcripts were later shortened by limited alkaline hydrolysis to 200-250 nucleotide fragments (Cox et al. 1984).
Antibodies and Western Blotting
Antisera were raised in rabbits against a fusion protein containing the last 40 amino acids of Oatp 1 (Eckhardt et al. 1999) and against a synthetic peptide consisting of the 15 terminal amino acids at the carboxy termi nus of Oatp 2 (648-662) coupled to KLH (keyhole limpet hemocyanin). Rabbits were immunized as described (Stieger et al. 1994). Western blots were performed with basolateral rat liver plasma membranes for Oatp 1 (Eckhardt et al. 1999) and Oatp 2 (Reichel et al. unpublished observations) and with crude rat brain membranes (see legend to Figure 1).
In Situ Hybridization Histochemistry (ISHH)
After decapitation of the animals the brains were removed and frozen immediately on dry ice. Cryostat sections (12 μm) were mounted on glass slides coated with 3-aminopropyltriethoxysilane (Sigma; St Louis, MO) and stored at −80C until use. The sections were fixed with 2% paraformaldehyde in PBS for 20 min, acetylated in 0.1 M triethanolamine hydrochloride containing 0.25% acetic anhydride, and prehybridized for 2-3 hr at room temperature (RT) with hybridization buffer containing 50% formamide, 5 × standard saline citrate (SSC), 5 × Denhardt's solution, 250 μg/ ml yeast tRNA, and 500 μg/ml hering sperm DNA. Then the sections were hybridized overnight at 53C with the digoxigenin-labeled antisense or sense probe dissolved in the hybridization buffer at a concentration of about 1 μg/ml. Sections were then sequentially washed at 53C with SSC, using a final concentration of 0.1 × SSC/50% formamide for 20 min. Thereafter, the sections were processed for immunodetection with anti-DIG-alkaline phosphatase (Boehringer) and with nitrotetrazolium blue and X-phosphate as the color substrate (Boehringer) according to the manufacturer's instructions. Sections were incubated at RT for 1 h with the anti-DIG-alkaline phosphatase at a dilution of 1:5000 in 100 mM Tris-HCl (pH 7.5), 150 mM NaCl, and 1% blocking reagent. After several washings, additional incubations were performed with nitrotetrazolium blue/X-phosphate at a dilution of 1:50 and 1 mM levamisole (Sigma) in 100 mM Tris-HCl (pH 9.5), 100 mM NaCl, and 50 mM MgCl2 at RT overnight in the dark.
Immunofluorescence Staining
To more definitively localize Oatp 2 in brain capillary endothelial cells, cryostat sections subjected to ISHH analysis were also processed for immunofluorescence labeling with antibodies against von Willebrand factor (vWF; Sigma), an endothelial cell marker, and against glial fibrillary acidic protein (GFAP; Dako, Glostrup, Denmark), a marker for astrocytes. The sections were incubated overnight at 4C with the perspective antibody diluted in PBS containing 2% normal goat serum and 0.2% Triton X-100. The concentration of the rabbit anti-vWF was 1:400 and for the mouse antiGFAP 1:5000. Finally, the sections were washed with PBS and incubated for 30 min at RT with the Cy3-labeled secondary antibody (1:300) (Jackson Immunoresearch; West Grove, PA).
Immunoperoxidase Staining
Rats were anesthetized with sodium pentobarbital (40 mg/kg
Double Immunofluorescence Staining
Fresh-frozen and paraformaldehyde (2%)-fixed (20 min) cryostat sections (12 μm) were incubated overnight at 4C in PBS containing 2% normal serum, 0.2% Triton X-100, and the primary antibodies against Oatp 1 (1:500), Oatp 2 (1:5000), GFAP (1:5000), (α1 subunit of Na, K-ATPase (2 μg/ml; Upstate Biotechnology, Lake Placid, NY), and the P-glycoprotein (antibody C219, 1:50; Signet Laboratories, Dedham, MA). After washing in PBS, the sections were incubated at RT with affinity-purified secondary goat antibodies (Jackson Immunoresearch) labeled with Cy2 (1:100) and Cy3 (1:300). The immunofluorescently stained sections were analyzed by confocal laser microscopy (MRC 600; Bio-Rad Laboratories, Hercules, CA) using dual-channel illumination with simultanous image recording for the two different fluorochromes. The pictures were processed by IMARIS software (Bitplane; Zurich, Switzerland). Control experiments for anti-Oatp 2 specificity were performed by preincubating the antibody with increasing concentrations (3-5 μg/ml) of the peptide used for immunization. To control for crossreactivity by the secondary antibodies, one of the primary antibodies was omitted during the incubation.
Results
Immunoblot Analysis of the Oatp 2 Antiserum
As shown in Figure 1, the developed Oatp 2 antiserum reacted with two narrow protein bands of crude rat brain membranes (Figure 1, left lane). However, whereas the 66-kD protein remained constant, the 76-kD protein band disappeared after preabsorption of the Oatp 2 antibody in the presence of an excess of the oligopeptide used for immunization (Figure 1, right lane). A similar narrow 76-kD brain Oatp 2 protein band has very recently also been reported by others using a different antibody (Kakyo et al. 1999). Furthermore, our Oatp 2 antiserum did not recognize Oatp 1 in separate Western blot analysis of rat liver basolateral plasma membrane proteins, and both Oatp 1 and Oatp 2 demonstrated a distinct distribution pattern in immunofluorescent studies of rat liver (Eckhardt et al. 1999; Reichel et al. unpublished observations). Taken together, these data strongly indicate that the antiserum raised is specific for Oatp 2. Furthermore, the close correspondence between the size of the immunodetected brain Oatp 2 and the theoretical molecular mass calculated from the originally cloned Oatp 2 (∼73 kD; Noé et al. 1997) suggests that glycosylation of the native brain Oatp 2 is minor.
Localization of Oatp 2 in Brain Capillary Endothelial Cells
We first used ISHH to localize Oatp 2 mRNA in rat brain tissue. As shown in Figures 2A-2C, positive hybridization signals were associated exclusively with endothelial cells of cerebral capillaries. In contrast to the antisense probe, no positive hybridization signal was obtained with the sense probe (Figure 2A, inset), thus supporting the specific reactivity of the antisense ribo-probe with Oatp 2 mRNA. The ISHH-positive capillary endothelial cells were scattered diffusely over all brain regions, with the exception of the pineal gland, pituitary gland, subfornical organ, median eminance, and choroid plexus (not shown). Similar results were obtained at the protein level, where the Oatp 2 antibody also selectively reacted with the capillary endothelium (Figures 2D and 2E). Finally, after preabsorption of the Oatp 2 antiserum with the antigenic peptide used for immunization, the immunopositive reactivity of the brain capillary endothelium was completely lost (Figure 2F), again demonstrating the specificity of the Oatp 2 antiserum and supporting the idea that the preabsorption sensitive 76-kD protein band in Figure 1 corresponds to Oatp 2 in brain capillary endothelium cells.
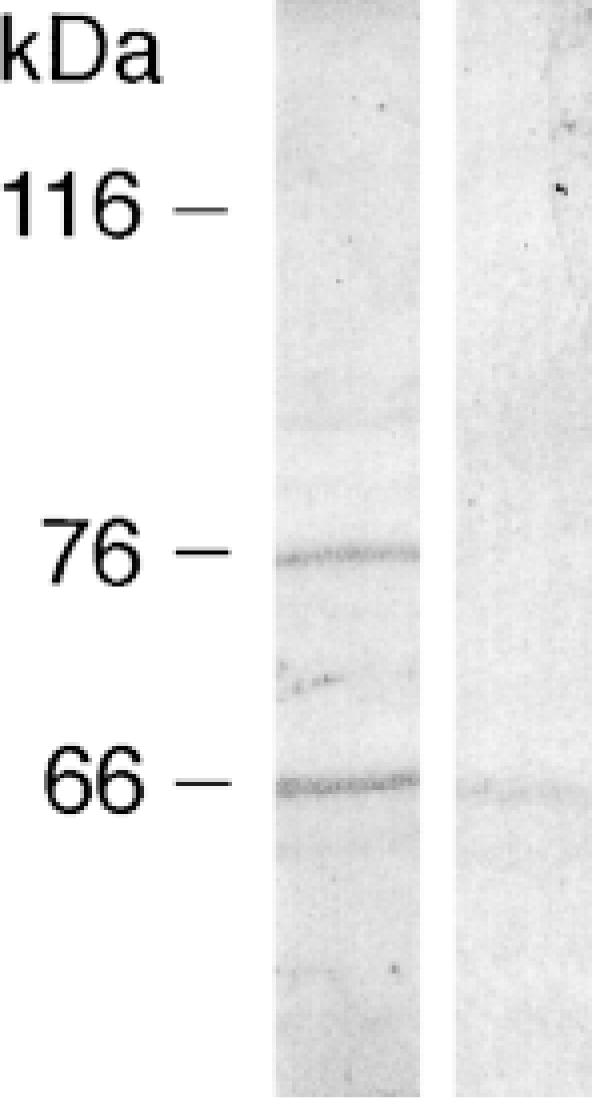
Western blot of crude rat brain membrane proteins with the Oatp 2 antiserum. Crude rat brain membranes were isolated as described (Wenzel et al. 1997). Membrane proteins were separated on 7.5% SDS-polyacrylamide minigels (30 μg protein per lane) and transferred to nitrocellulose membranes. Blots were blocked for 1 hr at RT in 10 mM Tris (pH 8.0), 150 mM NaCl, and 0.05% Tween (TBST) containing 5% blocker (nonfat dry milk; Bio-Rad). Blots were then incubated overnight at 4C with native Oatp 2 antiserum (dilution of 1:5000 in TBST containing 5% blocker) (left lane) or with Oatp 2-antiserum preincubated with 10 μg/ml of the peptide used for immunization (preabsorption of antibodies) (right lane). Blots were washed three times with 20 mM Tris (pH 7.5), 60 mM NaCl, 2 mM EDTA, 0.4% Triton X-100, 0.4% deoxycholate, and three times with TBST. They were then incubated with horseradish peroxidase-conjugated goat anti-rabbit IgG (Pierce; Rockford, IL) diluted at 1:5000 for 1 hr at RT. Immunoreactivity was visualized with the enhanced chemiluminescence method (NEN Life Science Products; Boston, MA) according to standard procedures.
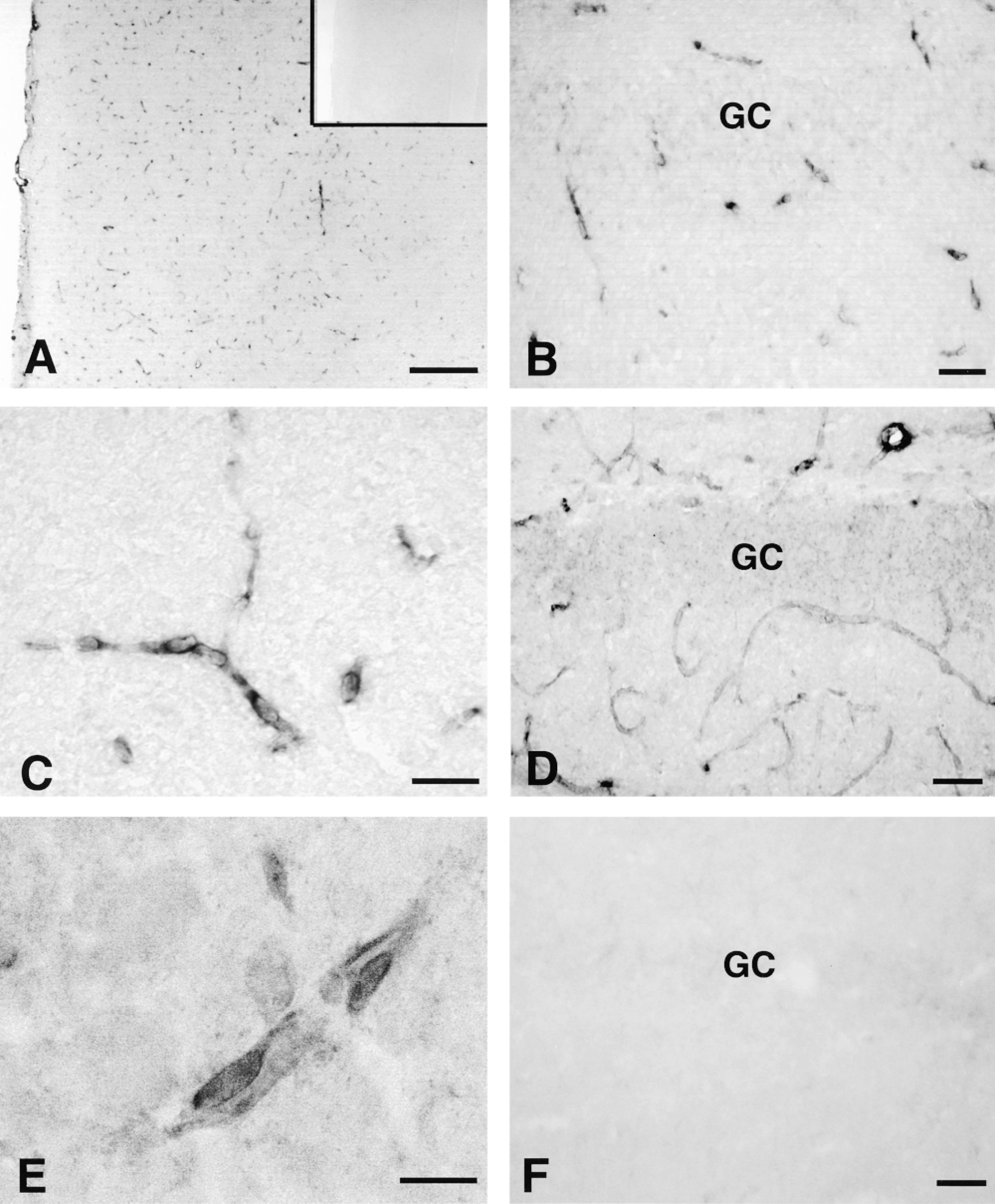
Localization of Oatp 2 in rat brain capillary endothelial cells by nonradioactive ISH (A-C) and immunohistochemistry (D,E). Sections of cerebral cortex (A) and hippocampal formation (B,C) were hybridized with the digoxigenin-labeled antisense probe for Oatp 2 mRNA. Inset in A shows an adjacent control section hybridized with the sense probe. (D) Immunoperoxidase staining of the hippocampal formation with the Oatp 2 antibody. (E) Higher magnification of immunopositive brain capillary endothelial cells. (F) Negative immunostaining after preabsorption of the Oatp 2 antiserum with the antigenic peptide used for immunization. GC, granular cell layer. Bars: A = 100 μm; B,D,F = 40 μm; C = 25 μm; E = 10 μm.
To more definitely discriminate between Oatp 2 expression in capillary endothelial cells and in astrocytes, double labeling experiments were performed that combined in situ hybridization for Oatp 2 mRNA with immunofluorescence staining for the endothelial marker vWF or the astrocyte marker GFAP. These experiments revealed that most, if not all, cells expressing Oatp 2 mRNA were immunopositive for vWF and vice versa (Figure 3A) and that positivity for Oatp 2 mRNA did not correlate with immunopositivity for GFAP, indicating that astrocytes do not express Oatp 2 (Figure 3B). In addition, double immunofluorescence staining with Oatp 2 and GFAP antibodies combined with confocal microscopic analysis indicated that Oatp 2 is localized on both the abluminal and the luminal domain of brain capillary endothelial cells (Figures 3C-3F). Furthermore, Figure 3E shows the intimate contacts between the brain capillaries and the surrounding GFAP-positive astrocyte endings that serve as ideal markers for the brain side of the blood vessels. In contrast, the blood-oriented luminal side of brain capillary endothelium is characterized by its expression of mdrla or P-glycoprotein (Stewart et al. 1996; Beaulieu et al. 1997). With the P-glycoprotein antibody C219, Figure 3F confirms that P-glycoprotein is selectively expressed at the luminal membrane, whereas Oatp 2 is localized on the entire surface domain of brain capillary endothelial cells. However, it should be noted that additional localization of Oatp 2 in intracellular vesicles of endothelial cells cannot be excluded at present.
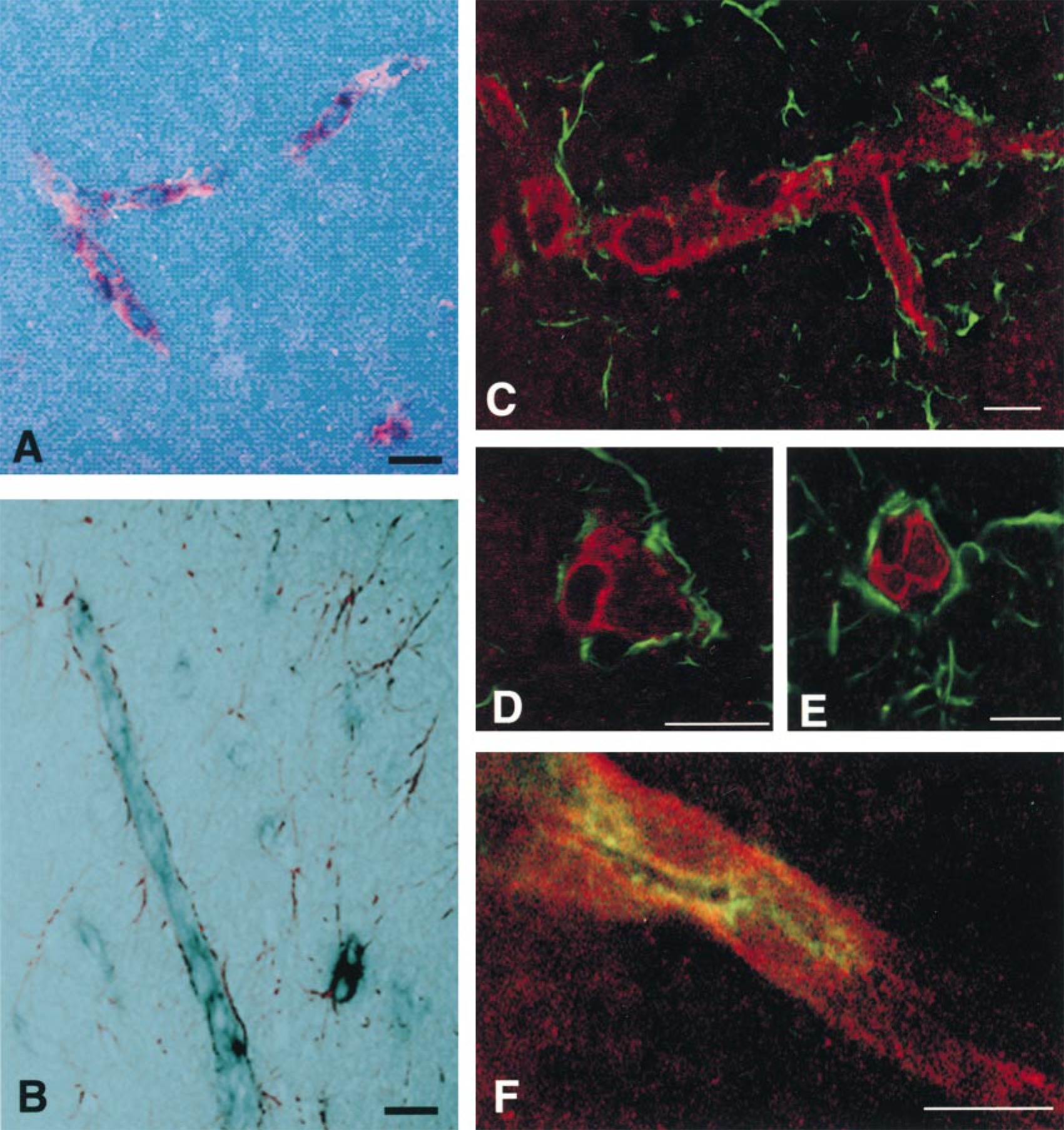
Cellular localization of Oatp 2 by double labeling techniques combining ISH and immunofluorescence staining. All pictures were analyzed by confocal laser microscopy and superimposition of the digitalized images. (A,B) The black color represents the hybridization signal of the Oatp 2 mRNA and the red color depicts the immunopositivity for the endothelial marker vWF (A) and the astrocyte marker GFAP (B). (C-E) Double immunostaining with the antibodies to Oatp 2 (red) and to GFAP (green). Oatp 2 is expressed only in endothelial cells and not in astrocytes. GFAP immunoreactivity indicates the brain (abluminal) side of the capillary endothelium. (F) Double immunostaining (yellow) of brain endothelial cells with antibodies to Oatp 2 (red) and to P-glycoprotein (C219; green). Whereas P-glycoprotein is selectively localized on the luminal side, Oatp 2 is present on the entire surface of rat brain endothelial cells. Bars = 10 μm.
Localization of Oatp 2 Compared to Oatp 1 in Choroid Plexus Epithelial Cells
Oatp 2 mRNA was also detected in the choroid plexus. However, in contrast to the BBB capillary endothelium, Oatp 2 mRNA (Figure 4A) and Oatp 2 protein (Figure 4B) were detected only in choroid plexus epithelial cells. Moreover, the immunofluorescence staining indicated that Oatp 2 is selectively expressed at the basolateral pole of choroid plexus epithelial cells (Figure 4B). This basolateral Oatp 2 expression is further shown in Figure 5, in which the apical membrane of the choroid plexus epithelial cells was selectively labeled with an antibody against the α1-subunit of the Na, K-ATPase (Marrs et al. 1993). In contrast to the apical co-localization of Na, K-ATPase and Oatp 1 (Figure 5F) (Angeletti et al. 1997), Oatp 2 immunoreactivity was localized to the opposite cell pole of choroid plexus epithelial cells (Figures 5A and 5C). Therefore, the basolateral Oatp 2 staining most probably represents the interdigitated basolateral plasma membrane of choroid plexus epithelial cells, although additional localization of Oatp 2 in intracellular vesicles cannot be excluded at this level of magnification. Nevertheless, the polar localization of Oatp 1 and Oatp 2 in choroid plexus epithelial cells again demonstrates the specificity of the antibodies used and contrasts to their expression in the liver, in which they are both localized at the basolateral plasma membrane of hepatocytes (Eckhardt et al. 1999; Kakyo et al. 1999; Reichel et al. unpublished observations).
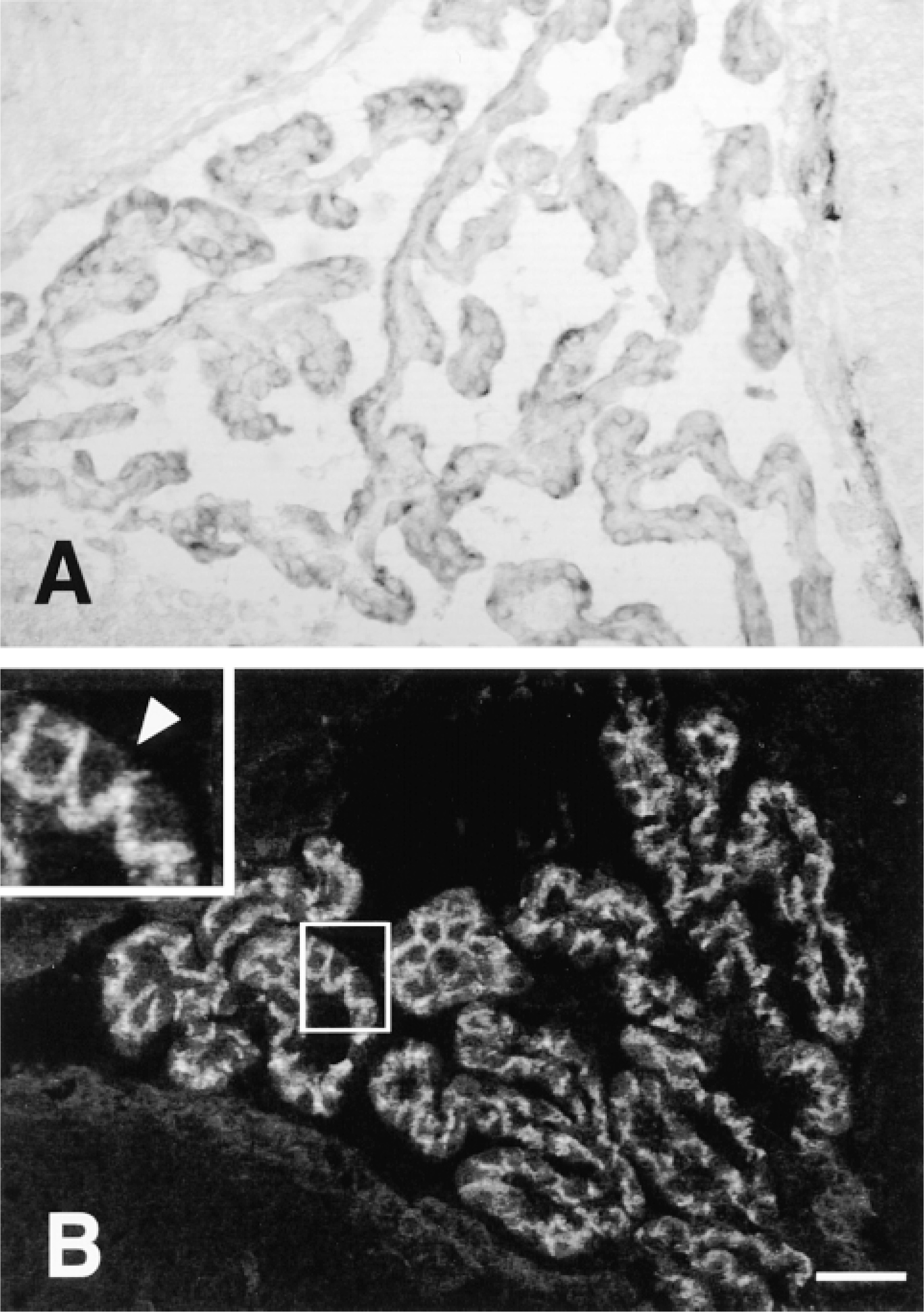
Localization of Oatp 2 in the choroid plexus. (A) ISH with the antisense probe for Oatp 2 mRNA. (B) Immunofluorescence staining for Oatp2. The boxed area has been enlarged to indicate more clearly the putative selective basolateral expression of Oatp 2. Arrow points to the apical membrane of epithelial cells. Bar = 20 μm.
Discussion
This study, for the first time, localizes the multispecific organic anion transporter Oatp 2 at the BBB and demonstrates a polar distribution of Oatp 1 and Oatp 2 in the choroid plexus epithelium.
The BBB barrier is formed by a tight endothelium lining the cerebral microvessels. Effective tight junctions restrict solute uptake across the paracellular pathway to small water-soluble compounds and make transcellular routes necessary for uptake of larger organic solutes into brain tissue. These transcellular routes have been divided into a diffusional pathway for lipidsoluble agents, specific transport systems for essential nutrients such as glucose, amino acids, and purines, and receptor-mediated and adsorptive endocytosis for hormones and plasma proteins, respectively (Abbott and Romero 1996). Although passive diffusion can account for brain penetration of many nonionic very lipid-soluble drugs, ionized compounds require carrier-mediated translocation across the plasma membrane of the BBB endothelium (Spector 1990). Our study demonstrates that in addition to the “renal-like” p-aminohippurate system (Wong et al. 1993), the BBB endothelium also localizes the “liver-like” organic anion carrier Oatp 2 (Figures 2 and 3). This polyspecific transporter can mediate transmembrane transport of a wide range of amphipathic substrates, including many drugs, drug conjugates, and small peptides (Meier et al. 1997; Noé et al. 1997; Kakyo et al. 1999). Its high expression at the surface of BBB endothelial cells (Figure 3), together with its high affinity for the cardiac glycoside digoxin (Noé et al. 1997), indicates that Oatp 2 might be responsible for the extensive intracerebral accumulation of digoxin in brain tissue of mdr 1a (- /-) knockout mice (Mayer et al. 1996; Schinkel et al. 1995,1996). Furthermore, Oatp 2 appears to mediate uptake of certain centrally active peptides, such as the δ1-opioid receptor agonist [
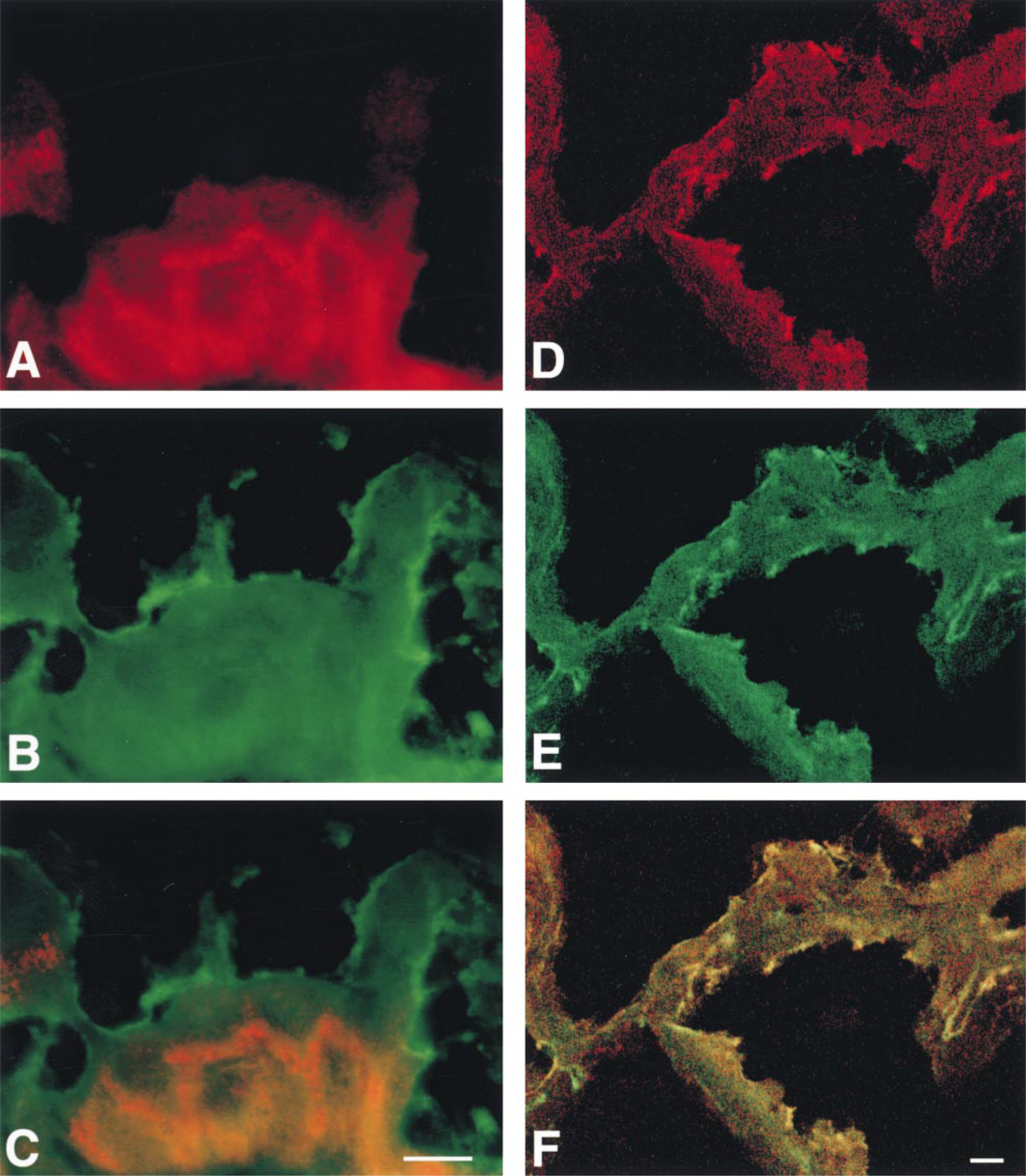
Polar expression of Oatp 2 (A-C) and Oatp 1 (D-F) at choroid plexus epithelial cells, as visualized by their double fluorescence staining with the apically localized α1-subunit of Na, K-ATPase (Marrs et al. 1993). (A) Staining for Oatp 2. (B,E) Staining for Na,K-ATPase. (C) Superimposition of A and B showing localization of Oatp 2 (red) at the opposite (basolateral) cell pole compared to Na,K-ATPase (green). (D) Staining for Oatp 1. (F) Superimposition of D and E showing co-localization of Oatp 1 and Na,K-ATPase at the apical plasma membrane of choroid plexus epithelial cells (yellow). Bars = 10 μm.
Interestingly, Oatp 2 is also localized at the basolateral cell pole of choroid plexus epithelial cells (Figures 4 and 5). The polar localization of Oatp 1 and Oatp 2 at the apical and basolateral surface domains, respectively, indicates that these two transporters may serve complementary functions in the transport of amphipathic organic anions into and/or out of the cerebrospinal fluid. Previous in vitro studies have provided evidence for the presence of at least one “liver-like” organic anion transporter in the choroid plexus of mammalian brain (Barany 1973a). Interestingly, the transport kinetics and substrate specificity of this (these) “liver-like” transport system(s) (Barany 1973b, 1975) correspond closely to the transport properties of Oatp 1 and Oatp 2 (Meier et al. 1997; Noé et al. 1997; Kakyo et al. 1999), indicating that these Oatps are responsible for “liver-like” transport properties of choroid plexus epithelial cells. Furthermore, because Oatp 1 mediates low-affinity transport of GSH and high-affinity transport of leukotriene C4 (Li et al. 1998), it could be involved in the apical secretion of GSH into CSF (Anderson et al. 1989) in exchange for leukotriene C4 uptake from cerebrospinal fluid (Spector and Johanson 1989). Whether and to what degree Oatp 2 exhibits similar efflux and/or uptake functions at the basolateral plasma membrane of choroid plexus epithelial cells remains to be determined.
In contrast to a recent study by Abe and co-workers (1998), we found no evidence for expression of Oatp 2 mRNA in hippocampal neuronal cells (Figure 2). A similar discrepancy between their study and ours also exists with respect to Oatp 2 mRNA expression in the cerebellar cortex (our negative data; not shown). The most probable cause of these discrepancies lies in the different ISHH methodology used. Whereas Abe and co-workers used a radiolabeled antisense cRNA probe, we used a digoxigenin-labeled antisense probe and a different ISH protocol. It is noteworthy, however, that in preliminary experiments with a radiolabeled probe we also found strong positivity in hippocampal and cerebellar neuronal cells. This neuronal labeling was judged as unspecific because similar positive labeling was also found with the sense probe. Furthermore, the neuronal cell labeling collapsed with the digoxigenin-labeled antisense probe, whereas the strong labeling of the brain capillary and choroid plexus epithelial cells remained constant independently of the labeling methods used. Finally, no immunopositivity of hippocampal and/or cerebellar neuronal cells was found with the Oatp 2 antiserum used. Hence, although this study presents strong evidence for Oatp 2 expression in the brain capillary endothelium and in the choroid plexus epithelium at both the mRNA and the protein level, definitive demonstration of additional expression of Oatp 2 in neuronal cells requires further investigation.
In conclusion, we have localized Oatp 2 at the BBB endothelium and demonstrate a polar distribution of Oatp 1 and Oatp 2 at the apical and basolateral cell poles of choroid plexus epithelial cells. At both sites, the expression of members of the Oatp gene family of membrane transporters can explain some phenotypic transport activities previously identified in functional in vivo and in vitro studies. The results demonstrate that more carriers than hitherto assumed (Abbott and Romero 1996) exist at the BBB and in the choroid plexus for amphipathic organic anion and drug transport into and out of the brain and the cerebrospinal fluid, respectively. Because these carriers must be taken into account in the targeting of therapeutic drugs to the brain tissue, further delineation of the exact functional role, directionality of transport, and structure/ activity relationships of Oatp 1 and Oatp 2 under various physiological and pathophysiological conditions is important for a better understanding of their “gatekeeping” functions at their strategic localization between the blood and brain compartments.
Footnotes
Acknowledgements
Supported by the Swiss National Science Foundation (grant 31-045536.95) and the Olga Mayenfisch Foundation, Zurich, Switzerland.
We thank Dr H.U. Luder for allowing us to use the technical facilities of the Division of Oral Structural Biology of the University Centre for Dental Medicine, Zurich, Switzerland.