Cytochemical Localization of Ouabain-sensitive,K + -dependent p -Nitrophenylphosphatase Activity in the Choroid Plexus of Normal and Reserpinized Guinea Pigs
Free accessBrief reportFirst published online August, 1998
Cytochemical Localization of Ouabain-sensitive,K + -dependent p -Nitrophenylphosphatase Activity in the Choroid Plexus of Normal and Reserpinized Guinea Pigs
SUMMARY High doses of reserpine induce depletion of biogenic amines. The K-NPPase activity of choroid plexus was determined after one-shot reserpine administration using cerium-based cytochemistry. In normal untreated animals, reaction product was found on the microvilli of the choroidal epithelium but was almost undetectable 3 and 7 days after reserpinization. At 20 days after reserpinization, however, it was detectable. These findings suggested that reserpine decreased the choroidal Na,K-ATPase activity, and that catecholamines might be essential to maintain normal choroidal Na,K-ATPase activity.
Ouabain-sensitive, K+-dependent adenosine triphosphatase (Na,K-ATPase) participates in the active transport of Na+ and K+ ions. Na,K-ATPase activity is high in the choroid plexus and is responsible for the production and maintenance of the cerebrospinal fluid (Masuzawa et al. 1981; Watts et al. 1991). Reserpine belongs to the family of rauwolfia alkaloids and induces the release of biogenic amines. Using a cerium-based method (Kobayashi et al. 1987), Kanoh et al. (1993) and Kanoh (1994) determined the cytochemical localization of ouabain-sensitive, K+-dependent, p-nitrophenylphosphatase (K-NPPase) activity in strial marginal cells after reserpine administration and found that the enzyme activity was undetectable. Kanoh (1997) also investigated the cytochemical localization of K-NPPase activity in the reserpinized facial nerve and reported that the reaction product was undetectable on the internodal axolemma. The aim of the present study was to determine if there are any changes in K-NPPase activity in the choroid plexus after reserpine administration.
The choroid plexus was obtained from 12 Hartly guinea pigs, weighing 400-500 g, with a normal Prayer's reflex at 3 days (n = 3), 7 days (n = 3), and 20 days (n = 2) after one-shot reserpine administrat on (10 mg/kg IP) and from untreated animals (n = 4). Under deep ketamine hydrochloride anesthesia (IM), the animals were perfused through the heart with a fixative containing 2% paraformaldehyde and 0.05% glutaraldehyde in 0.1 M cacodylate buffer (pH 7.4). After decapitation, the choroid plexi were dissected from the excised brain and immersed in the same fixative for 1 hr at 4C. The tissue samples were rinsed in 50 mM Tricine buffer (pH 7.5) for 15 min and then incubated in medium according to the cerium method (Kobayashi et al. 1987). After cytochemical incubation, the tissues were postfixed with 2% OsO4 in 0.2 M cacodylate buffer (pH 7.4) for 1 hr at room temperature. The specimens were then dehydrated through a graded series of alcohol solutions and propylene oxide and embedded in Spurr's epoxy resin. Uncontrasted ultrathin sections were then observed under a JEM-1200 electron microscope (JEOL; Tokyo, Japan). The animal use protocol was approved by the Institutional Animal Care and Use Committee of the Hyogo College of Medicine, and all experiments were performed in accordance with the guidelines of the Declaration of Helsinki.
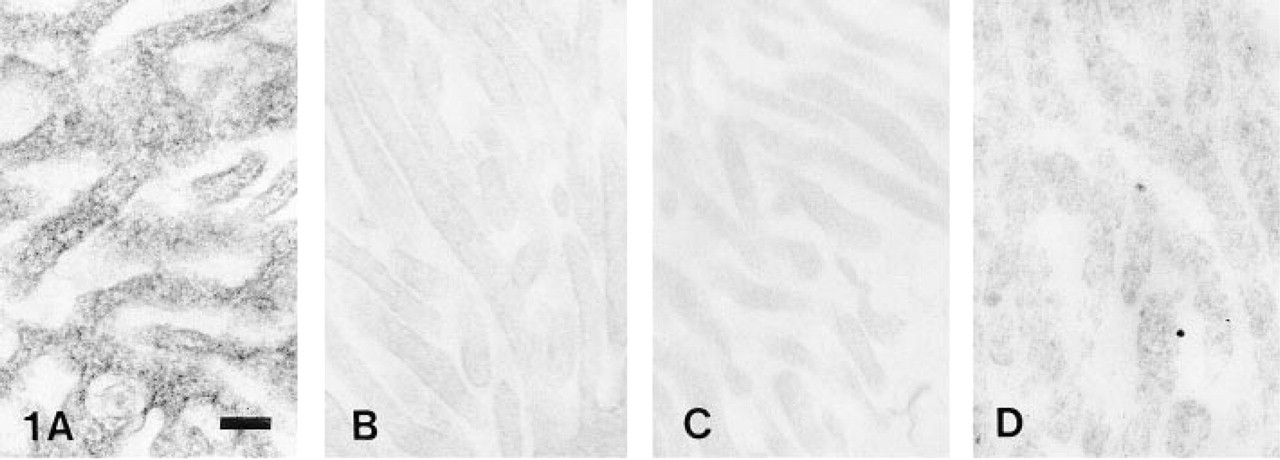
The cytochemical examination of K-NPPase activity revealed a fine granular reaction product that was found exclusively on the microvilli of normal choroidal epithelium (Figure 1A). In choroid plexus 3 days after reserpinization (Figure 1B) and 7 days after reserpinization (Figure 1C), enzyme reaction product was almost undetectable. However, at 20 days after reserpinization reaction product was detectable (Figure 1D). The formation of reaction product was almost completely inhibited when 10 mM ouabain was included in the medium (Figure 2A), and reaction product was undetectable in a substrate-free medium (Figure 2B), and in a medium in which K+ had been replaced with Na+ (Figure 2C).
(A) Normal animal. A fine granular reaction product of K-NPPase is distributed on the choroid plexus of a guinea pig. (B-D) Reserpine treatment (B) Day 3, (C) Day 7, and (D) Day 20. (B, C) Reaction product is almost suppressed. (D) Reaction product is detectable. Bar = 0.2 μm.
(A-C) Negative controls for K-NPPase activity. (A) Incubation with 10 mM ouabain. (B) Incubation in a substrate-free medium. (C) Incubation in a medium in which K+ had been replaced with Na+. Bar = 0.2 mm.
Controlling the CSF volume and electrolyte composition is necessary for normal neural functioning and is essential for maintaining physiological brain volume and intracranial pressure (Lindvall et al. 1978; Masuzawa et al. 1981; Watts et al. 1991). It is widely recognized that the choroid plexus plays a major role in the production and regulation of CSF secretion. The active transport of sodium across the choroidal epithelia has also been implicated as the driving force for CSF secretion. Several factors have been reported to affect CSF production. In infant rats, choroidal Na, KATPase activity matures rapidly during the first 2-3 weeks after birth (Parmelee and Johanson, 1989). Pollay et al. (1985) showed that ouabain almost completely inhibited choroidal Na,K-ATPase activity and decreased CSF formation by 70-80%. The present experimental results also demonstrated that choroidal K-NPPase activity was greatly decreased from Day 3 to Day 7 after reserpine administration, which was similar to the strial (Kanoh et al. 1993; Kanoh 1994) and facial nerve K-NPPase activity (Kanoh 1997). Because high doses of reserpine induced the depletion of catecholamines, these results indicate that catecholamines may be essential for the regulation and maintenance of normal choroidal Na,K-ATPase activity and CSF production.
Footnotes
Acknowledgements
Supported by a Grant-in-Aid (08671998) for Science Research from the Ministry of Education, Science and Culture of Japan.
References
1.
KanohN (1994) Reserpine inhibits the Na-K ATPase activity in the stria vascularis of the cochlea.Laryngoscope104: 197–200
2.
KanohN (1997) Cytochemical localization of ouabain-sensitive, K+-dependent, p-nitrophenylphosphatase activity in the facial nerve of reserpinized guinea pigs.J Histochem Cytochem45: 1129–1135
3.
KanohNKumoiTKobayashiTOkadaTSeguchiH (1993) Ultracytochemical localization of K-dependent, p-nitrophenylphosphatase activity in stria vascularis of reserpinized guinea pig.Acta Otolaryngol (Stockh)113: 142–145
4.
KobayashiTOkadaTSeguchiH (1987) Cerium-based cytochemical method for detection of ouabain-sensitive, potassium-dependent, p-nitrophenylphosphatase activity at physiological pH.J Histochem Cytochem35: 601–611
5.
LindvallMEdvinssonLOwmanCH (1978) Sympathetic nervous control of cerebrospinal fluid production from the choroid plexus.Science201: 176–178
6.
MasuzawaTSaitoTSatoF (1981) Cytochemical study on enzyme activity associated with cerebrospinal fluid secretion in the choroid plexus and ventricular ependyma.Brain Res222: 309–322
7.
ParmeleeJTJohansonCE (1989) Development of potassium transport capability by choroid plexus of infant rats.Am J Physiol256:R786–791
WattsAGWattsGSEmanuelJRLevensonR (1991) Cell-specific expression of mRNAs encoding Na,K-ATPase α- and β-subunit isoforms within the rat central nervous system.Proc Natl Acad Sci USA88: 7425–7429