
Abstract
We examined by immunocytochemistry the localization of the AP-1 family proteins c-Jun, JunB, JunD, c-Fos, FosB, Fra-1, and Fra-2 in rat incisor ameloblasts. Most of the antibodies against AP-1 family proteins, except for c-Fos-specific antibody, labeled ameloblast nuclei. The labeling intensity of the c-Jun, JunD, and Fra-2 antibodies was stronger than that of JunB, FosB, and Fra-1. Antibody reactivities of c-Jun, JunD, and Fra-2 were greatly enhanced during or after the transition zone. Furthermore, c-Jun antibodies labeled maturation ameloblasts in a cyclic pattern, which was correlated with ameloblast modulation. Disruption of ameloblast modulation by colchicine injection resulted in greatly decreased reactivity of the c-Jun antibody in the ameloblast nuclei of the maturation zone. Phospho-specific antibodies to c-Jun labeled ameloblast nuclei only weakly throughout the secretion, transition, and maturation zones. These results suggest that the stage-specific localization of AP-1 in ameloblasts is closely related to tooth enamel formation.
AP-1
Rodent incisors are a suitable model to learn about tooth enamel formation because the incisor teeth of adult animals include the entire range of ameloblasts where they proliferate, participate in enamel matrix formation and enamel maturation, and end their roles. In a secretion zone of enamel matrix formation, ameloblasts secrete several matrix proteins, a major component of which is amelogenin (Smith 1998). Although the promoter region of the bovine amelogenin gene is known to include an AP-1 binding motif, its regulatory role in enamel formation is unknown (Gibson et al. 1997). In the narrow transition zone between enamel matrix secretion and enamel maturation, a considerable number of ameloblasts die via apoptosis (Nishikawa and Sasaki 1995, 1996). However, the precise mechanisms of cell death are unknown. In the enamel maturation zone, two different types of ameloblasts are distinguished: the ruffle-ended ameloblast (RA) and the smooth-ended ameloblast (SA). Longer RA zones and shorter SA zones are repeated several times just after the transition zone up to the end of the maturation zone. That is called ameloblast modulation (Josephsen and Fejerskov 1977). Furthermore, the modulation moves as a wave in the incisal (erupting) direction more rapidly than tooth eruption speed. When this modulation wave is analyzed, it is estimated that an ameloblast repeats the RA–SA cycle 30–45 times in a maturation zone (Smith et al. 1987). Differences of morphology, calcium transport, and endocytosis between RA and SA appear to be associated with specific gene expression and the recruitment of proteins related to metabolism (Takano et al. 1982; Sasaki 1990; Kawamoto and Shimizu 1994; Smith 1998).
In this study, the localization of AP-1 family proteins in the ameloblasts was examined by immunohistochemistry to clarify the possible involvement of AP-1 in enamel secretion, transition, and enamel maturation.
List of AP-1 antibodies used in this study and reactivity to ameloblasts
Materials and Methods
Eleven male Wistar rats (7–8 weeks, 185–260 g) (Jcl Wistar; Clea Japan, Tokyo, Japan; institutional guidelines were followed) were used. The animals were perfused under sodium pentobarbital (Nembutal; Abbott, North Chicago, IL) anesthesia with 4% paraformaldehyde in 0.1 M phosphate buffer solution (PB), pH 7.2, via the left ventricle at room temperature (RT) for 10 min. They were immersed in the same fixative at 4C for 2 hr and the maxillary and mandibular incisors were dissected. After being washed with 0.1 M PB, the teeth were decalcified with 5% disodium EDTA solution adjusted to pH 7.3 with sodium hydroxide solution and were placed in a cold room for 3–4 weeks. Decalcified incisors were cut transversely into two segments. Both segments were further cut longitudinally (6–8-μm thick for light microscopy and 40–50-μm thick for electron microscopy) using the cryotome.
The antibodies used in this study were c-jun/AP-1(Ab-1) (lots D04353–3 and D04353–5) purchased from Oncogene Research Products (Cambridge, MA), p-c-Jun(KM-1), JunB(N-17), JunD(329), c-Fos(K-25), c-Fos(4), FosB(102), Fra-1(R-20), and Fra-2(Q-20) purchased from Santa Cruz Biotechnology (Santa Cruz, CA), and phospho-specific c-Jun(Ser 63)II purchased from New England Biolabs (Beverly, MA) (Table 1).
Cryosections on the glass microscopic slides were incubated with 0.6% hydrogen peroxide in 80% methanol to inhibit endogenous peroxidase activity at RT for 20 min. They were labeled with an appropriate primary antibody, described above, diluted with 1% bovine serum albumin in PBS (BSA-PBS) at RT for 1 hr or at 4C overnight. They were then labeled with HRP-conjugated anti-rabbit or anti-mouse IgG (Cappel; West Chester, PA) diluted 1:100 with 1% BSA-PBS at RT for 30 min. Some specimens were labeled with FITC-conjugated anti-rabbit IgG. HRP-labeled sections were incubated with a solution composed of 0.5 mg/ml diaminobenzidine (DAB) and 2 mg/ml (NH4)2Ni(SO4)2 6H2O in Tris-HCl (pH 7.6) at RT for 10 min, then with the DAB solution containing 0.005% hydrogen peroxide at RT for 2–12 min. Some of the specimens were developed with the AEC substrate kit (DAKO; Carpentia, CA). The sections were further labeled with 1 μg/ml Hoechst 33342 (Molecular Probes; Eugene, OR) to detect the transition zone (Nishikawa and Sasaki 1999a) or rhodamine–phalloidin (Molecular Probes) diluted 1:20 with PBS to detect ruffle-ended (RA) or smoothended ameloblasts (SA) (Nishikawa and Josephsen 1987).
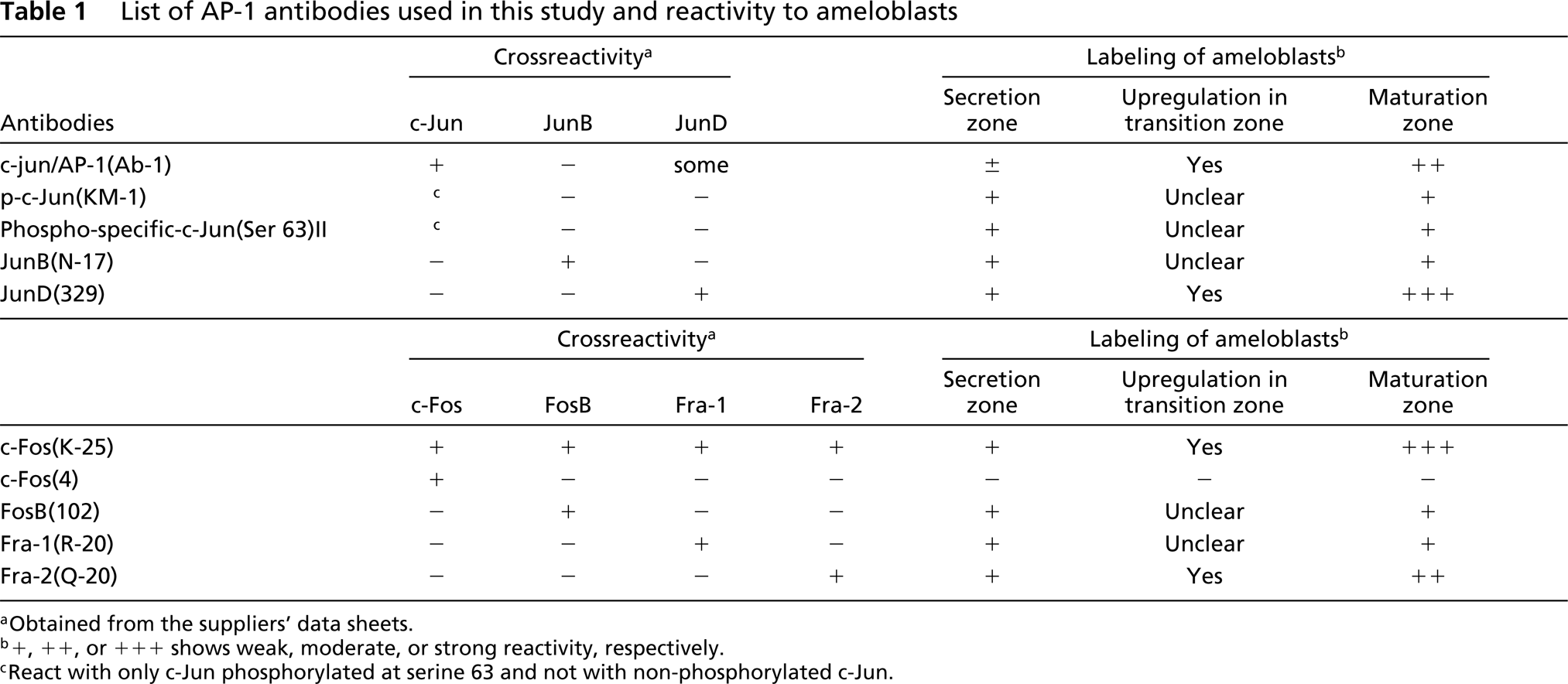
Immunocytochemical labeling of late secretion (S) and transition (T) ameloblasts by JunD(329) (
Immunocytochemical labeling of maturation ameloblasts by JunD(329) (
Procedures for pre-embedding immunoelectron microscopy were described elsewhere (Nishikawa and Sasaki 1999b). Briefly, thick cryosections fixed with paraformaldehyde and decalcified with EDTA were labeled with c-jun/AP-1(Ab-1) diluted 1:10 at 4C overnight, followed by HRP-conjugated anti-rabbit IgG at 4C overnight. They were incubated in a DAB-H2O2 solution and postfixed with 1% osmium tetroxide. Then they were embedded in Epon 812. Control sections were processed in the same way as described above, except that the primary antibody was replaced by 1% BSA-PBS. The positive reaction products of original electron micrographs were enhanced by NIH Image (developed at the National Institute of Health and available from the Internet by anonymous FTP).
For the colchicine experiment, five male Wistar rats (9 weeks, 300–310 g; Clea Japan) were used. Three rats were injected SC with a colchicine solution (Sigma–Aldrich Japan; Tokyo, Japan) (1.3 mg/kg bw; colchicine was dissolved with PBS in a concentration of 1 mg/ml) and two rats for control with the same amount of PBS alone. Eight hours later the animals were sacrificed as described above.
Results
Reactivity of Antibodies Against Jun and Fos Family Proteins in Ameloblasts
Most of the antibodies used in this study labeled ameloblast nuclei (Table 1). An exceptional case was c-Fos(4) antibody, which had no reactivity in the ameloblasts, whereas c-Fos(K-25), which has broad reactivity to Fos family proteins (Table 1), labeled ameloblasts. JunD(329), c-Fos(K-25), Fra-2(Q-20) and c-jun/AP-1(Ab-1) antibodies labeled maturation ameloblast nuclei strongly, but the labeling intensity of secretion ameloblasts was weak (Table 1; Figures 1–4). During or just after the transition zone, the labeling intensity drastically increased (Figures 1 and 4).
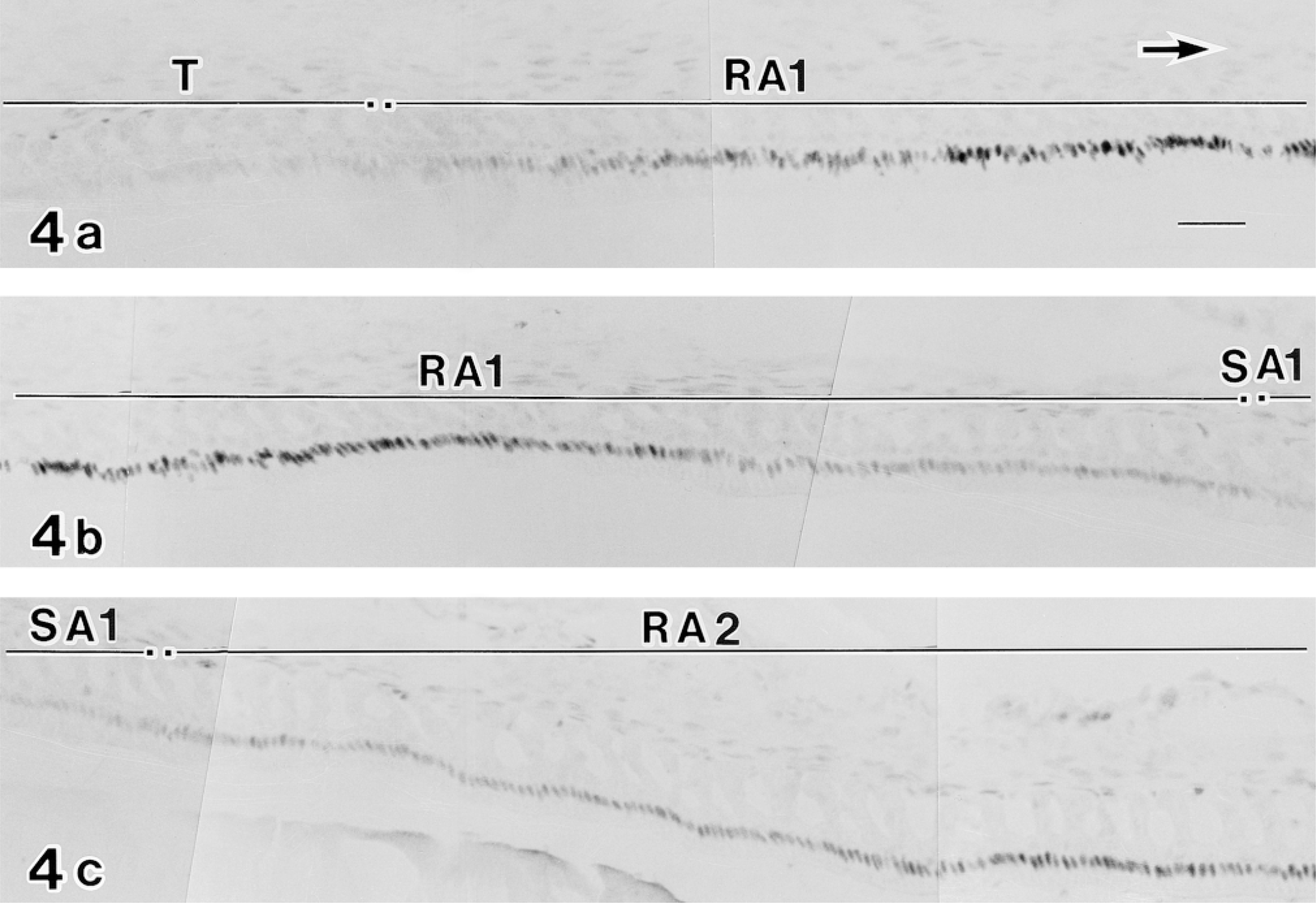
Labeling pattern of c-Jun/AP-1(Ab-1) antibody in ameloblasts from the proliferation to the early maturation zone. All micrographs were taken from the same section of an incisor. (
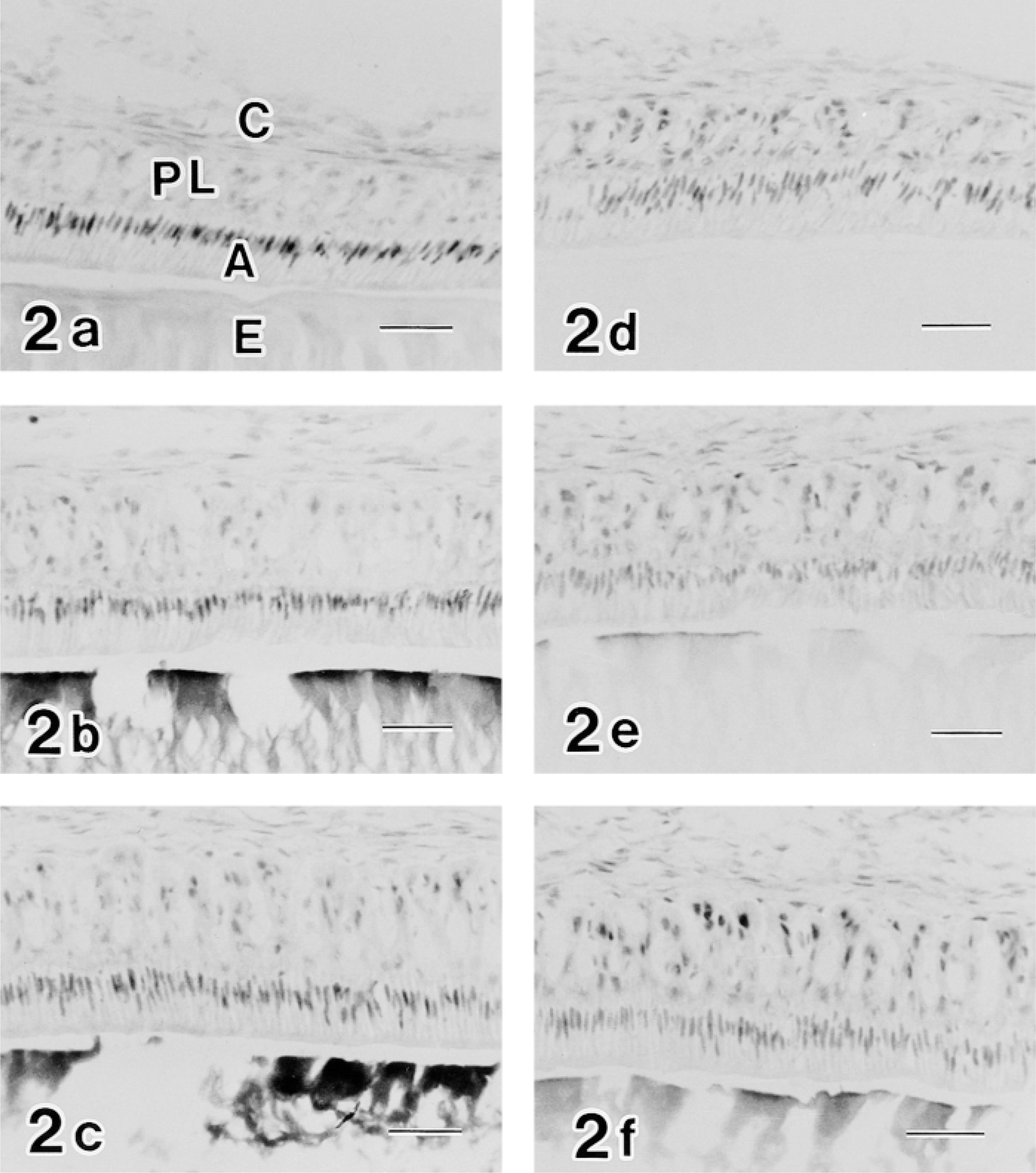
In the case of the c-jun/AP-1(Ab-1) antibody, the labeling pattern was complex. In the proliferation zone at the apical end of teeth, nuclei of several cells constituting the centrally located core of the stellate reticulum (SR) and the surrounding inner and outer enamel epithelia were positive for the c-jun/AP-1(Ab-1) antibody (Figure 3a). During differentiation, c-jun/AP-1(Ab-1)-positive epithelial cells were limited to the stratum intermedium (SI) and the SR (Figure 3b). In the early secretion zone, SI cells continued to be positive and the ameloblast nuclei were weakly labeled (Figure 3c). In the middle secretion zone most reactivity was lost (Figure 3d), whereas in the late secretion zone modest reactivity of ameloblast nuclei was observed (Figure 3e). The transition zone also exhibited low levels of c-jun/AP-1(Ab-1) reactivity (Figure 3f). Reactivity in maturation ameloblast nuclei, however, was greatly enhanced (Figures 3g and 4).
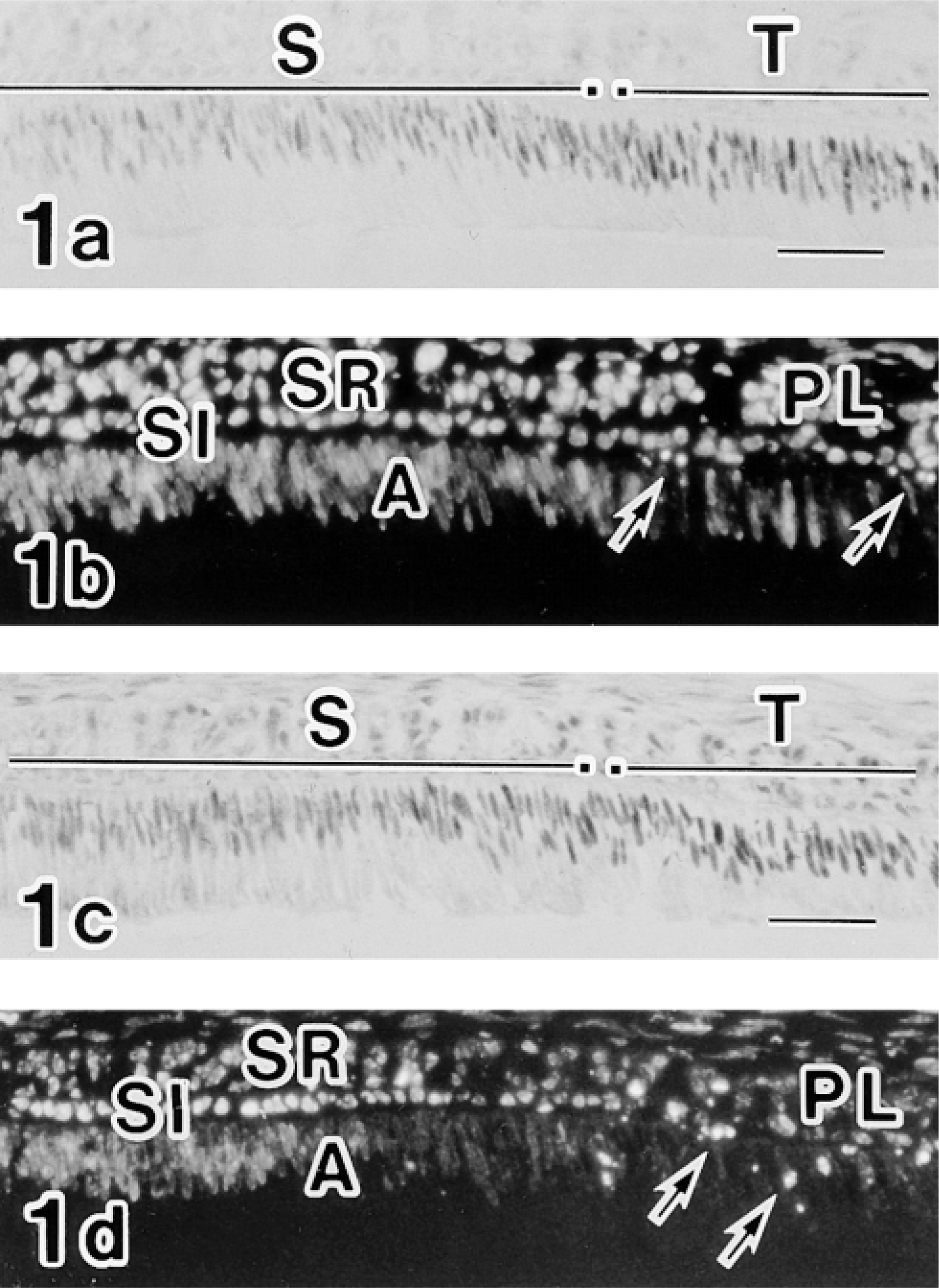
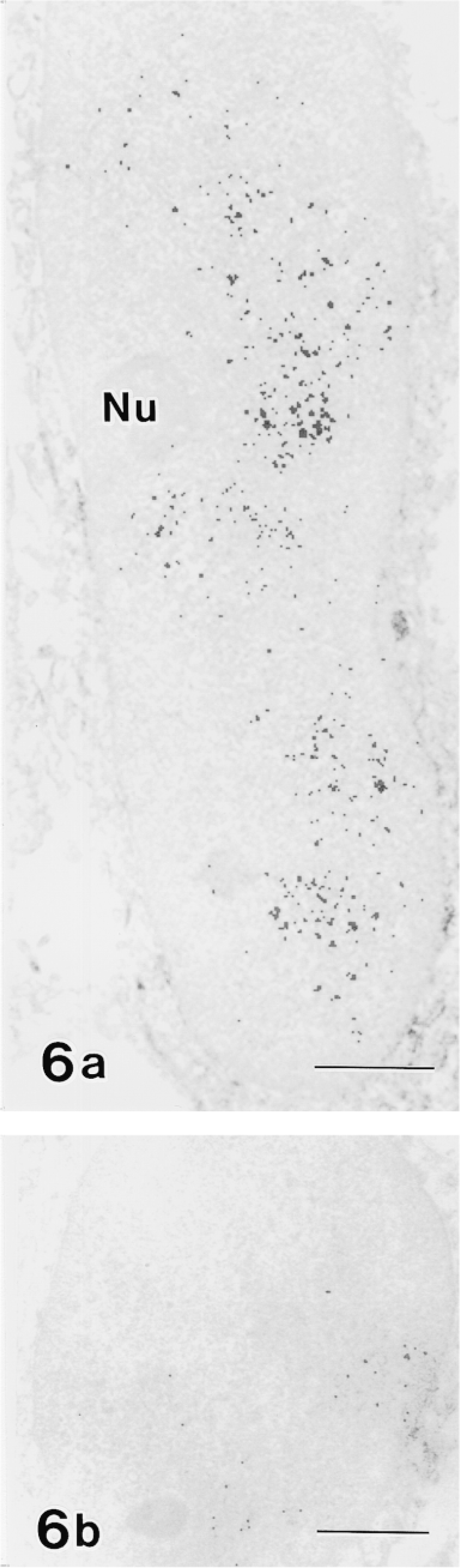
Moreover, the labeling of c-jun/AP-1(Ab-1) changed cyclically in the maturation zone. Whereas RAs were intensely labeled, SAs exhibited relatively weak labeling (Figure 4). In double-labeling experiments, maturation ameloblasts positive for c-jun/AP-1(Ab-1) exhibited intense labeling of rhodamine–phalloidin at the apical end, which confirmed that c-Jun-rich ameloblasts were RAs (Figure 5). This cyclic pattern repeated several times until ameloblasts underwent the pigment release stage, but JunD(329), c-Fos(K-25), and Fra-2(Q-20) antibodies did not exhibit the cyclic labeling pattern despite strong reactivity in the maturation zone. In reduced enamel epithelia, the labeling intensity of c-jun/AP-1(Ab-1) antibody decreased. These labeling patterns were identical in two different lots (D04353-3 and D04353-5) of c-jun/AP-1(Ab-1). The ultrastructure labeled by c-jun/AP-1(Ab-1) was interchromatin space (Figure 6).
Composite micrographs showing immunocytochemical labeling of the c-Jun/AP-1(Ab-1) antibody in the ameloblast nuclei of the early maturation zone (
p-c-Jun(KM-1) and phospho-specific c-Jun(Ser63)II antibodies, both of which react only to c-Jun phosphorylated at serine 63 and do not react with nonphosphorylated c-Jun, labeled ameloblast nuclei weakly in the secretion and maturation zones and provided no evidence of cyclic changes or increased labeling intensity in the maturation zone (Figures 7a and 7b).
JunB(N-17), FosB(102); and Fra-1(R-20) antibodies labeled ameloblasts (Figures 7c–7h). No cyclic changes of labeling intensity were observed, thus showing continuous labeling in the maturation zone. Compared with the overall labeling intensity of c-jun/AP-1(Ab-1), JunD(329), c-Fos(K-25), and Fra-2(Q-20) antibodies, that of JunB(N-17), FosB(102), and Fra-1(R-20) was weak.
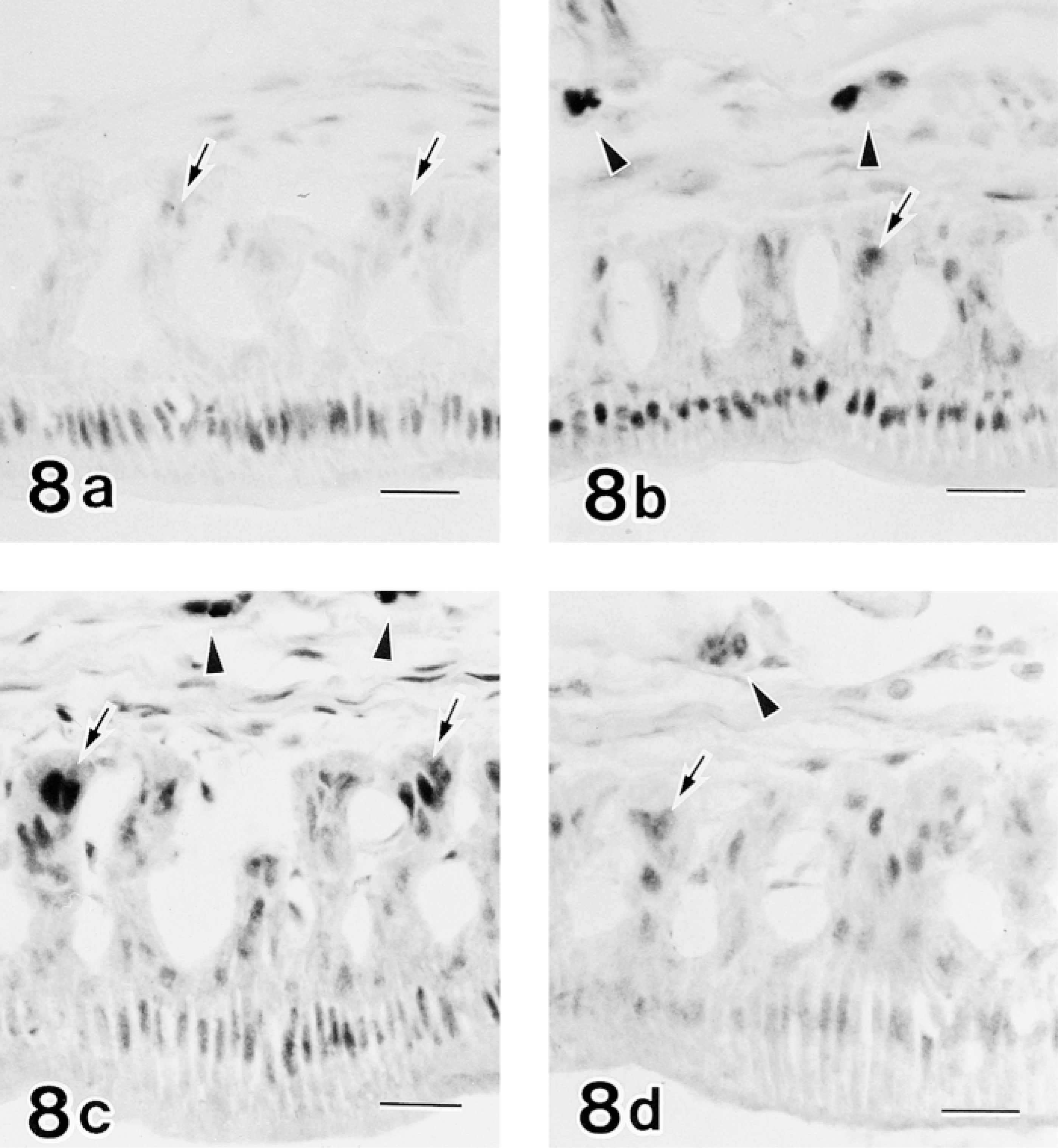
In the late maturation zone, long protruded processes of the papillary layer were attached to connective tissues. Epithelial cells constituting the apex of each process exhibited strong reactivity for c-jun/AP-1(Ab-1), JunD(329), c-Fos(K-25), and Fra-2(Q-20) (Figure 8). Osteoclasts located in the inner surface of the alveolar bone covering incisors were strongly positive for JunD(329), c-Fos(K-25), and Fra-2(Q-20), but not for c-jun/AP-1(Ab-1) (Figure 8).
Negative control sections processed without the primary antibody did not show any specific labeling. Positive control sections of keratinized oral epithelia or epidermis showed positive labeling of c-jun/AP-1(Ab-1) in epithelial cell nuclei, especially strong in the cell nuclei just before cornification, consistent with a previous report (Welter and Eckert 1995). JunD(329) labeled epithelial cell nuclei of basal and middle layers but did not label the cells just below the stratum corneum. C-Fos(K-25) and Fra-2(Q-20) labeled epithelia in a similar pattern to c-jun/AP-1(Ab-1), which was consistent with previous studies (Fisher et al. 1991; Byers et al. 1995).
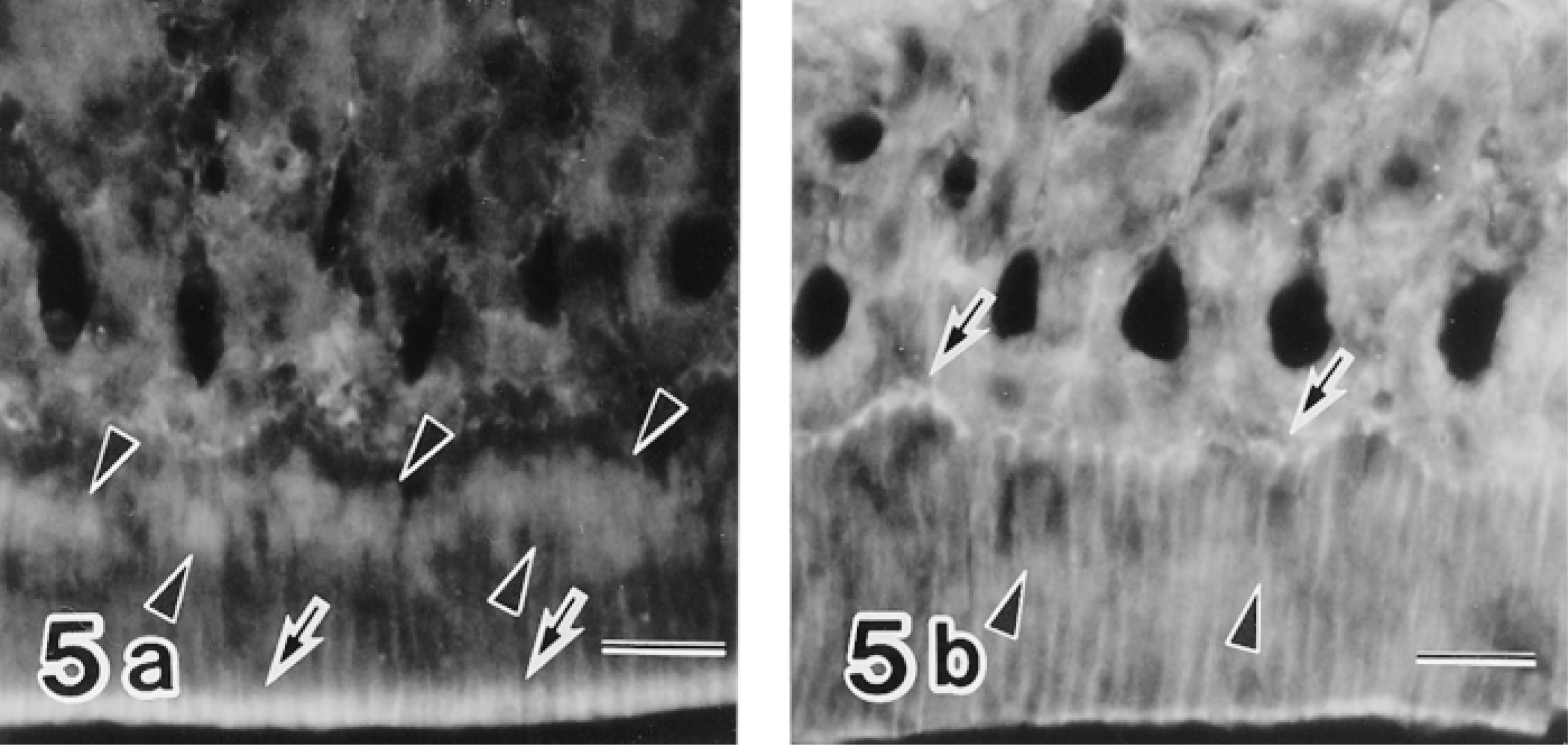
Immunofluorescence micrographs doubly labeled with the c-Jun/AP-1(Ab-1) antibody followed by FITC-conjugated anti-rabbit IgG and rhodamine–phalloidin, showing preferential labeling of RA nuclei (arrowheads in
Changes in the Reactivity of c-jun/AP-1 (Ab-1) in Maturation Ameloblasts of Colchicine-injected Rat Incisors
Colchicine experiments were conducted to determine the possible relationship between cyclic change of c-jun/AP-1(Ab-1) labeling and ameloblast modulation in the maturation zone. Eight hours after injection of 1.3 mg/kg colchicine, most RAs disappeared and SA-like cells became predominant, thus showing the disruption of cyclic ameloblast modulation (Figure 9). c-jun/AP-1(Ab-1) antibody reactivity in colchicine-injected specimens decreased significantly. Only small regions of maturation ameloblast nuclei were labeled by the c-jun/AP-1(Ab-1) antibody (Figure 11). Labelings of JunD(329) and c-Fos(K-25) were also disturbed, resulting in a smaller reacted area. Disturbance of AP-1 family protein reactivity was more severe in the early maturation zone than in the late maturation zone.
Specimens from control rats showed normal ameloblast modulation (Figure 10) and the same pattern of antibodies against Fos and Jun family proteins as those from normal rats.
Discussion
Various AP-1 family proteins were detected in the ameloblasts and epithelial cells constituting the stratum intermedium, stellate reticulum, and outer enamel epithelia, although the labeling patterns showed considerable variation. The present results support previous findings of the presence of Fos/Fra reactivity in the odontoblasts and ameloblasts of rat molars and incisors (Byers et al. 1995), and c-jun and junB expression in the odontoblasts and ameloblasts of rat molars (Kitamura and Terashita 1997; Kitamura et al. 1999). It is suggested that AP-1 is involved in the regulation of amelogenesis. Localization of the c-jun/AP-1(Ab-1) antibody in the proliferation zone may be related to the mitogenic activity of enamel organ cells. Weak reactivity of c-jun/AP-1(Ab-1), JunD(329), Fra-2(Q-20), and c-Fos(K-25) in the secretion zone may be related to the function of regulating enamel matrix formation.
As for the role of AP-1 in apoptosis, ameloblasts in the late secretion zone exhibited transient reactivity of the c-Jun/AP-1(Ab-1) antibody in their nuclei in this study. Although this transient increase may indicate a positive role of c-Jun in ameloblast apoptosis, the significance of the relationship is unclear.
Unexpected results included an increase of some AP-1 family proteins in the maturation zone. It clearly shows that there are more AP-1 proteins in maturation ameloblasts than in secretion ameloblasts, suggesting active gene transcription in maturation ameloblasts. Alternatively, it is possible but unlikely that all AP-1 member proteins detected in this study play a role in suppressing gene transcription (Rossi et al. 1998). The ultrastructural localization of the c-jun/AP-1(Ab-1) antibody in this study supported the transcriptional regulation of this protein, because transcription is considered to occur in the interchromatin space (Fakan 1994).
An interesting result was the cyclic labeling pattern of the c-jun/AP-1(Ab-1) antibody in maturation ameloblasts. A colchicine experiment was conducted to examine the relationship between the RA–SA modulation cycle and AP-1. Eight hours after SC injection of 1.3 mg/kg colchicine, most RAs disappeared. Instead, SA-like cells were predominant throughout the maturation zone in this study, which is consistent with previous reports (Akita et al. 1983; Ohya et al. 1993). In these teeth, the reactivity of the c-jun/AP-1(Ab-1) antibody was greatly reduced. Only a small number of ameloblast nuclei were weakly labeled. AP-1, especially c-Jun, may regulate the modulation cycle. C-Jun has been shown to be activated with phosphorylation on Ser63 and Ser73 by c-Jun N-terminal protein kinase (JNK) when UV irradiation or tumor necrosis factor (TNF) is applied to cells (Karin et al. 1997). On the other hand, calcium-regulated transcriptional activation of c-Jun and c-Jun-mediated G1 progression of the cell cycle do not require JNK activation (Cruzalegui et al. 1999; Wisdom et al. 1999). In this study, p-c-Jun(KM-1) and phospho-specific c-Jun(Ser63)II antibodies, which react only with c-Jun phosphorylated at Ser63, labeled ameloblast nuclei weakly in the secretion and maturation zones. Therefore, the enhanced labeling of the c-jun/AP-1(Ab-1) antibody in the maturation zone in this study may not be associated with phosphorylation.
(

Immunocytochemical labeling of secretion (
Immunocytochemistry of late maturation zone labeled with c-Jun/AP-1(Ab-1) (
In the ameloblast nuclei of the maturation zone, strong reactivity to JunD(329), c-Fos(K-25), and Fra-2(Q-20) antibodies was characteristic in this study. Considering the broad reactivity of the c-Fos(K-25) antibody to FosB, Fra-1 and Fra-2 antigens in addition to c-Fos antigen, no reactivity of the c-Fos(52) antibody to ameloblasts, and stronger reactivity of the Fra-2-specific antibody to the maturation ameloblasts than that of FosB and Fra-1, the major component of Fos family proteins in the maturation ameloblasts is concluded to be Fra-2. Therefore, the Fra-2/JunD heterodimer may be a predominant AP-1 member in the maturation ameloblasts. Another heterodimer may be Fra-2/c-Jun. In an in situ hybridization study of AP-1 in developing chicken gastrointestinal tracts, the predominant AP-1 species were reported to be fra-2 and junD (Matsumoto et al. 1998). Another report that examined the regulation of laminin α3A (laminin 5 subunit) gene transcription by transforming growth factor revealed that AP-1, especially Fra-2 and JunD, are involved in that process (Virolle et al. 1998).
In conclusion, c-Jun, JunD, and Fra-2 are major AP-1 components of ameloblasts during amelogenesis of rat incisors. Target genes for stimulation or suppression remain to be clarified.
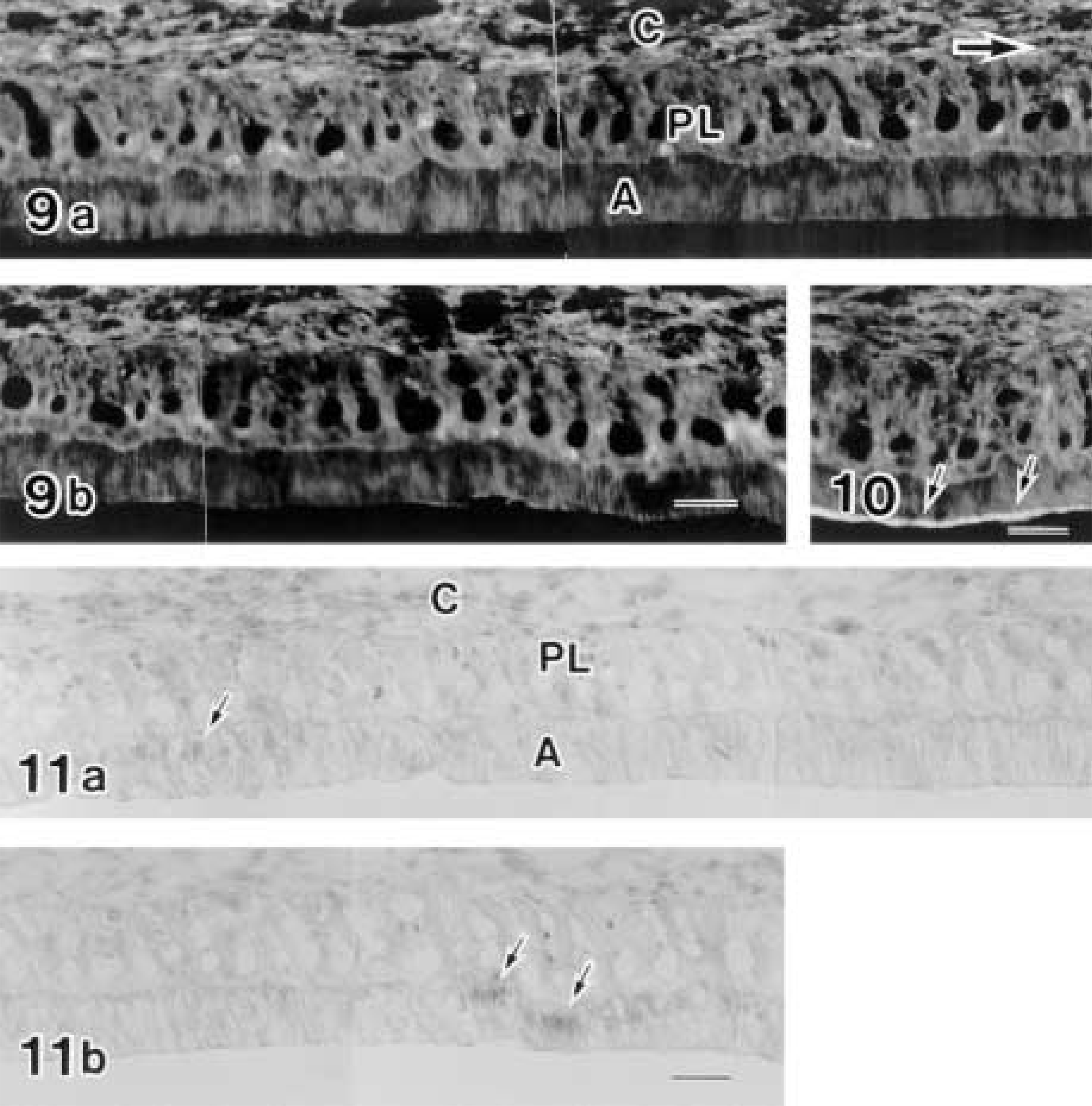
Early maturation zones of maxillary incisors from a colchicine-injected rat (Figures 9 and 11) and from a physiological saline-injected rat (Figure 10). Figures 9b and 11b are direct continuations of Figures 9a and 11a, respectively. Two sections of Figures 9 and 11 are obtained from an incisor and are close to each other at intervals of three 6-μm sections. Figure 9 shows rhodamine–phalloidin labeling and exhibits loss of apical concentration of F-actin, suggesting the loss of the ruffle-ended form of maturation ameloblasts. RAs from the control rat exhibit apical concentration of F-actin fluorescence (arrows in Figure 10). Figure 11 shows the labeling of the c-Jun/AP-1(Ab-1) antibody and exhibits only restricted positive reaction (arrows) in the ameloblast nuclei. Most ameloblast nuclei are devoid of labeling. Large arrow in Figure 9a shows incisal direction. C, connective tissue; PL, papillary layer; A, ameloblast. Bars = 50 μm
Footnotes
Acknowledgements
Supported in part by a Grant-in-Aid for scientific research (No. 11671824) from the Ministry of Education, Science, Sports and Culture of Japan.
I thank Dr Margaret R. Byers, University of Washington, Seattle for allowing me to examine her Fos/Fra-labeled specimens and for helpful discussion. For use of the facilities in the laboratory, acknowledgments are dedicated to Prof Fumie Sasaki.