
Abstract
Dendritic cells in the enamel organ of rat incisors were examined with immunocytochemistry using an anti-cystatin C antibody for immature dendritic cells and macrophages, OX6 for MHC Class II, ED1 for macrophages and dendritic cells, and ED2 for macrophages. Single cells positive for anti-cystatin C appeared in the enamel organ in zones at which ameloblasts secrete enamel matrix proteins. They were also present in transition and enamel maturation zones. In addition, ameloblasts, osteocytes, and osteoclasts were labeled by anti-cystatin C. ED1 and ED2 immunocytochemistry revealed that there was no macrophage population in the enamel organ of secretion, transition, or enamel maturation zone. A double labeling study showed that most anti-cystatin C-positive cells in the enamel maturation zone were also positive for OX6, whereas anti-cystatin C-positive and OX6-negative cells were prevalent in the secretion zone. The results suggest that immature dendritic cells penetrate the enamel organ of the secretion zone and begin to mature in the zones of transition and enamel maturation.
Keywords
D
During amelogenesis in rat incisors, ameloblasts secrete enamel matrix proteins, forming an enamel layer. The same ameloblasts also contribute to enamel maturation, which is accompanied by organic matrix loss and increased mineralization. A narrow zone called the transition zone lies between the secretion and maturation zones. A cell kinetic study of rat incisors (bw 100 g and 300 g) revealed that it takes 6.5–7.5 days for an ameloblast to pass the secretion zone, less than 1 day to pass the transition zone, and 12–22 days to pass the maturation zone (Smith and War-shawsky 1977). During the long maturation zone of rat incisors, a number of dendritic cells can be observed by using anti-MHC Class II antibody (OX6) immunohistochemistry, although their functional roles are still unknown (Takano et al. 1996). Several OX6-positive macrophage-like cells are observed, and some of them ingest apoptotic fragments in the transition zone, where 25% of the ameloblasts are lost by apoptosis (Nishikawa and Sasaki 1995, 1996, 1998; Smith and Warshawsky 1977). Two questions are raised. When do dendritic cells penetrate into the enamel organ? Are the macrophage-like cells in the transition zone the same kind of cells as the dendritic cells in the maturation zone?
In this study, antibodies against cystatin C known to be endogenous cysteine protease inhibitors were used to address these questions and ascertain the nature of OX6+ cells in the enamel organ. Cystatin C was shown to be abundant in immature dendritic cells, and therefore the proteolytic processing of the MHC Class II protein complex and antigen proteins was inhibited. On the other hand, cytostatin C was decreased in mature dendritic cells where protein processing occurs, and antigen–MHC Class II complexes were expressed in the plasma membrane (Pierre and Mellman 1998).
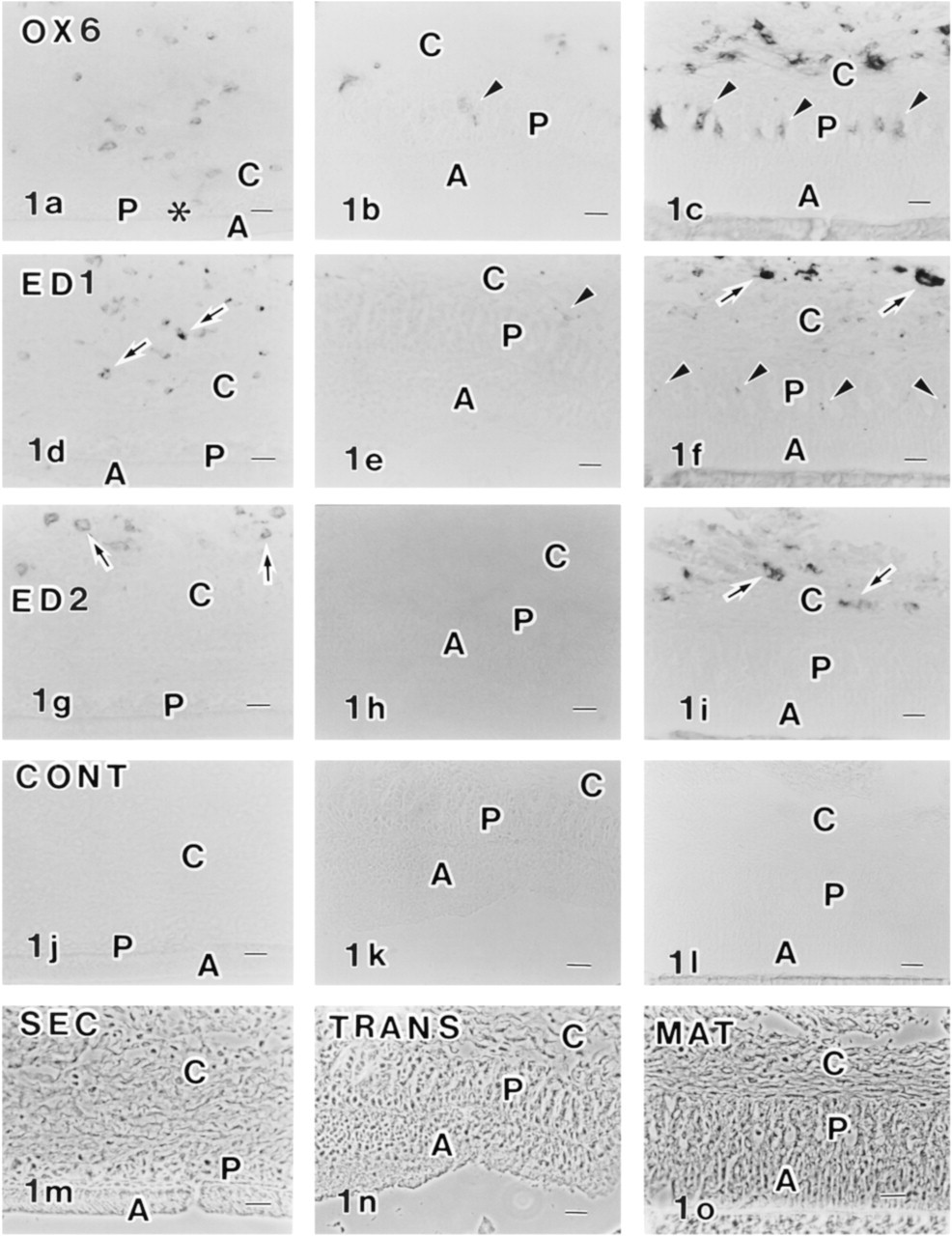
Immunocytochemical localization of OX6 (
Materials and Methods
Three male Wistar rats (280–290 g) (Jcl Wistar; Clea Japan, Tokyo, Japan; institutional guidelines were followed) were sacrificed by diethyl ether inhalation and maxillary and mandibular incisors were dissected. The labial side of each tooth was scraped with a razor blade. Tissue pieces containing secretion, transition, and early maturation ameloblasts were immediately frozen in a cryotome (HM505E; Microm, Walldorf, Germany). Cryosections (6–8 μm thick) were laid on glass microscopic slides and fixed with pure acetone at 4C for 5 min. The immunoperoxidase labeling method was described previously (Nishikawa and Sasaki 1999b). Briefly, the sections were labeled with OX6, ED1, or ED2 antibodies (Serotec; Oxford, UK) diluted 1:100, 1:50, or 1:50, respectively. They were then labeled with HRP-conjugated anti-mouse IgG (Cappel; West Chester, PA) diluted 1:100, followed by development with an AEC substrate chromogen system (AEC; 3-amino-9-ethylcarbazole as chromogen; DAKO, Carpinteria, CA). Control experiments were processed in the same way as described above except that the primary antibody was replaced by 1% bovine serum albumin in phosphate-buffered saline (BSA–PBS).
Procedures for pre-embedding immunoelectron microscopy for the OX6 antibody were described previously (Nishikawa and Sasaki 1996). Briefly, aldehyde-fixed and EDTA-decalcified incisors were cryosectioned (40–50 μm thick) in the cryotome. The sections were labeled with OX6 antibody diluted 1:100, followed by HRP-conjugated anti-mouse IgG (Cappel) diluted 1:100. They were then processed with diaminobenzidine (DAB) and H2O2 and embedded in Epon 812 (TAAB; Redding, UK). Some specimens were labeled by gold-conjugated anti-mouse IgG (10 nm; Amersham, Poole, UK) instead of HRP-conjugated anti-mouse IgG.
For immunocytochemistry of the anti-cystatin C antibody (Upstate Biotechnology; Lake Placid, NY), 11 male Wistar rats (185–260 g; Jcl Wistar, Clea Japan) were used. This anti-human cystatin C antibody has been shown to react specifically with murine 13–15-kD cystatin C (Pierre and Mellman 1998) and rat cystatin C (supplier's data sheet). The animals were perfused under sodium pentobarbital (Nembutal; Abbott, North Chicago, IL) anesthesia with 4% paraformaldehyde in 0.1 M phosphate buffer solution (PB, pH 7.2, via the left ventricle at room temperature (RT) for 10 min. They were immersed in the same fixative at 4C for 2 hr. The maxillary and mandibular incisors were dissected. After being washed with 0.1 M PB, the teeth were decalcified with 5% disodium EDTA solution adjusted to pH 7.3 with sodium hydroxide solution and were placed in a cold room for 3–4 weeks. Decalcified incisors were cut transversely into two segments. Both segments were further cut longitudinally (6–8 μm thick for light microscopy and 40–50 μm thick for electron microscopy) using the cryotome.
For light microscopy, cryosections were labeled with anti-cystatin C antibody diluted 1:100 with 1% BSA–PBS at RT for 60 min and then labeled with FITC-conjugated anti-rabbit IgG (Cappel) diluted 1:50 with 1% BSA–PBS at RT for 30 min. Most of the cryosections were triply labeled with anti-cystatin C followed by FITC-conjugated anti-rabbit IgG, with OX6 antibody followed by rhodamine-conjugated anti-mouse IgG (Cappel), and with Hoechst 33342 (1 μg/ml in PBS) (Molecular Probes; Eugene OR) for DNA. Some sections were triply labeled with anti-cystatin C, ED1, and Hoechst 33342. For numerical analysis, the number of anti-cystatin C+ cells and OX6+ cells in the enamel organ of the secretion zone or transition plus early maturation zone were counted along the long axis of the incisors using double labeled sections. The length of the secretion zone for counting was 2.7–4.7 mm (n = 6 sections, maxillary and mandibular incisors from two rats) and that of the transition plus early maturation zone was 2.4–4.6 mm (n = 6 sections, maxillary and mandibular incisors from two rats). Some sections were labeled with anti-OX62 antibody (MRC OX62, MCA1029G; Serotec) for some cells with dendritic shape and γδ T-cells, and with anti-CD45R (HIS24; Serotec) for pan-B-cells. Some acetone-fixed and undecalcified sections were double labeled with anti-cystatin C antibody and anti-desmoplakin 1 and 2 antibody (Clone DP1&2–2.15; Progen Biotechnik, Heidelberg, Germany) (Nishikawa et al. 1988) diluted 1:10, followed by FITC-conjugated anti-rabbit IgG and rhodamine-conjugated anti-mouse IgG diluted 1:50.
For immunoelectron microscopy, thick cryosections fixed with paraformaldehyde and decalcified with EDTA were labeled with anti-cystatin C diluted 1:100 with 1% BSA–PBS at 4C overnight, followed by HRP-conjugated anti-rabbit IgG (Cappel) diluted 1:100 with 1% BSA–PBS at 4C overnight. After being fixed with 1% glutaraldehyde solution for 1 hr, the sections were incubated in a DAB solution (0.2 mg/ml in 0.05 M Tris-HCl buffer, pH 7.6) at RT for 30 min and then immersed in DAB plus 0.005% H2O2 at RT for 5 min. They were postfixed with 1% osmium tetroxide and embedded in Epon 812. Ultrathin sections were lightly stained with lead citrate and examined with a JEOL 1200 EXII electron microscope (JEOL; Tokyo, Japan). Control sections were processed in the same way as described above, except that the primary antibody was replaced by 1% BSA–PBS. Procedures for conventional electron microscopy have been described elsewhere (Nishikawa and Sasaki 1995).
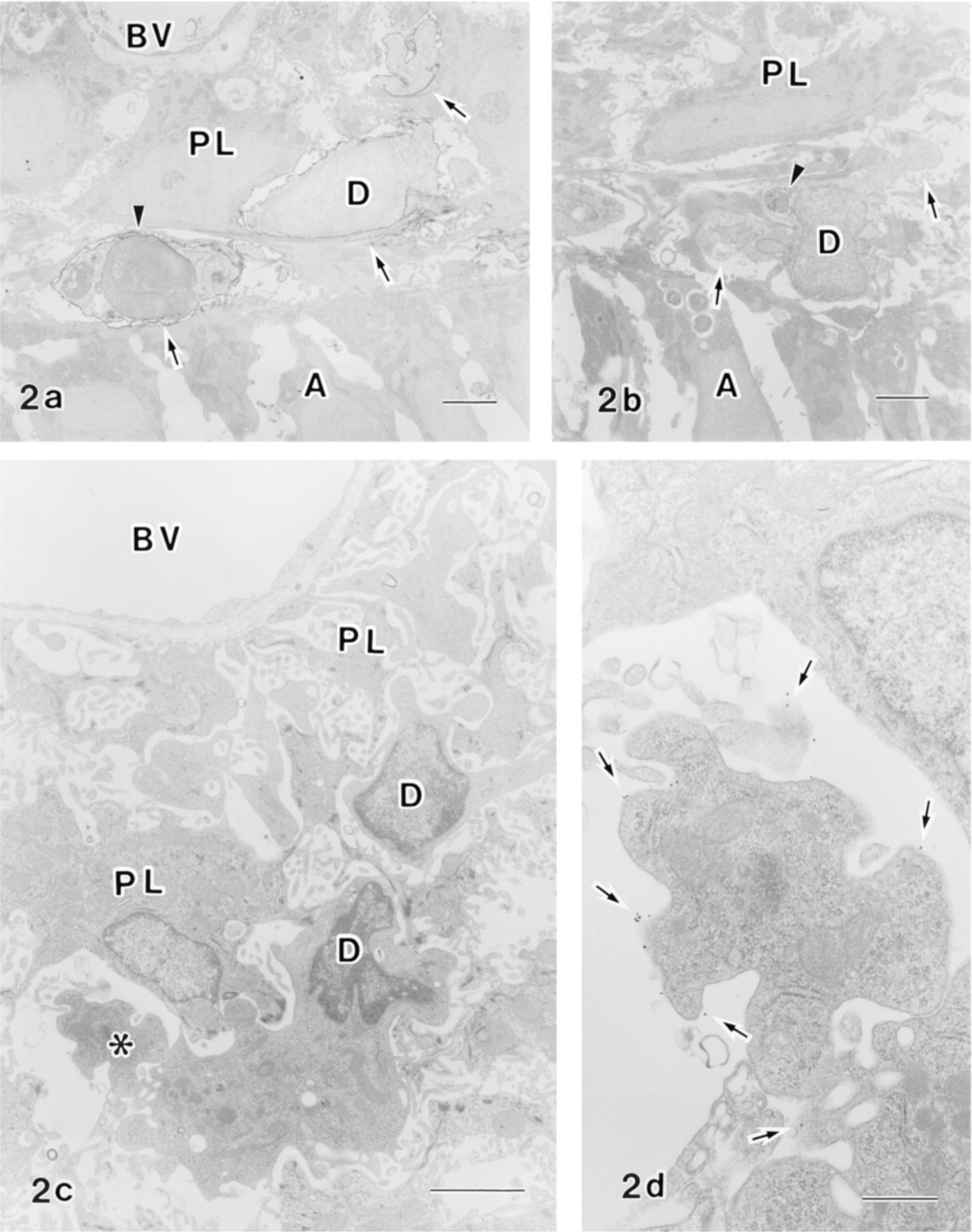
Immunoelectron micrographs of dendritic cells in the papillary layer of the transition zone (
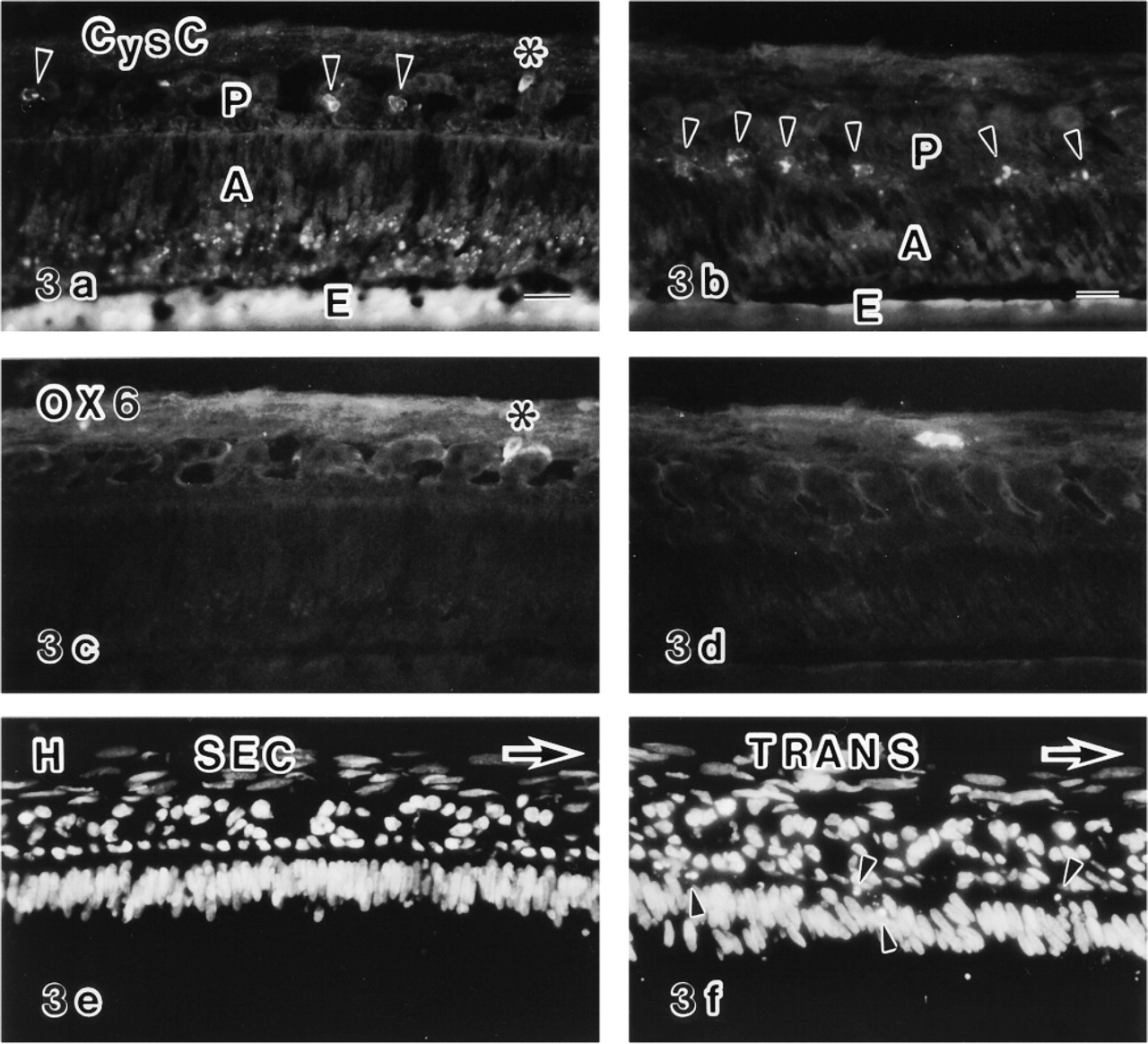
Triple labeled sections of enamel organs and overlying connective tissues of the secretion zone (SEC) (
Results
OX6+ Cells in Transition and Maturation Zones Are Dendritic Cells
OX6+, ED1 +, and ED2+ cells were scattered in connective tissues overlying the enamel organ in the secretion, transition, and maturation zones (Figures 1a–1i). In the enamel organ of the secretion zone, however, no OX6+, ED1 +, and ED2+ cells were present (Figures 1a, 1d, and 1g). OX6+ or ED1+ cells began to appear in the enamel organ of the transition zone (Figures 1b and 1e; and see Nishikawa and Sasaki 1996). They were localized in the papillary layer. In the maturation zone, OX6+ and ED1+ cells were abundant in the papillary layer (Figures 1c and 1f), which is consistent with a previous report (Takano et al. 1996). Double labeling of enamel organ in the maturation zone by OX6 and ED1 revealed the presence of doubly positive cells. Because ED2+ cells were not present in the enamel organ in any of the three zones (Figures 1g–1i), these OX6+ and ED1+ cells are considered to be dendritic cells. Control sections showed no specific labeling (Figures 1j–1o). Anti-CD45R antibody for B-lymphocytes did not react with the enamel organ except for the positive reaction of round cells in the nearby blood vessels. An anti-OX62 antibody that was recently shown to recognize the αE2-integrin of some dendritic cells and intraepithelial dendritic T-cells (Brenan and Rees 1997) strongly labeled particular cell populations in connective tissues of oral mucosa but did not label dendritic cells in the enamel organ.
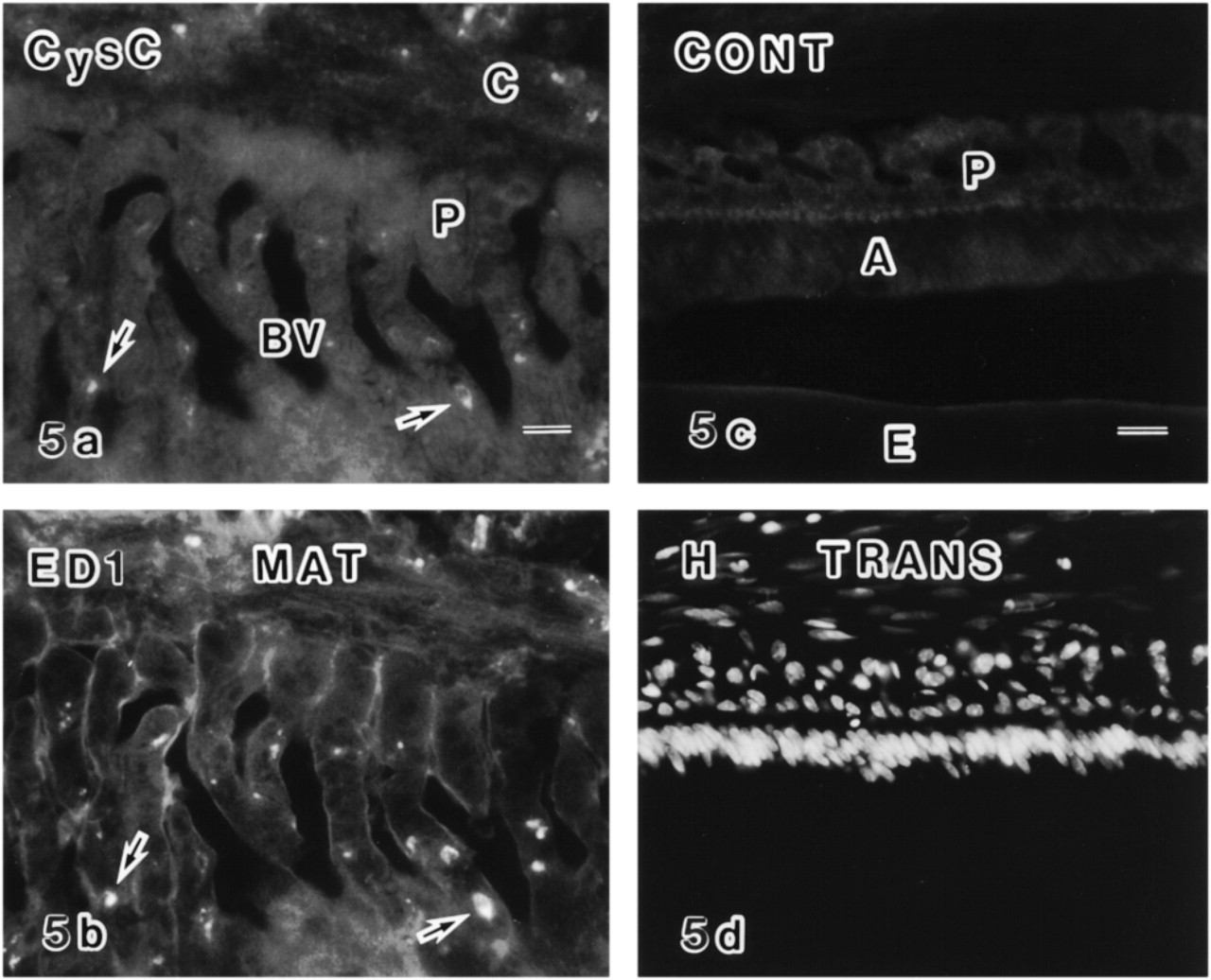
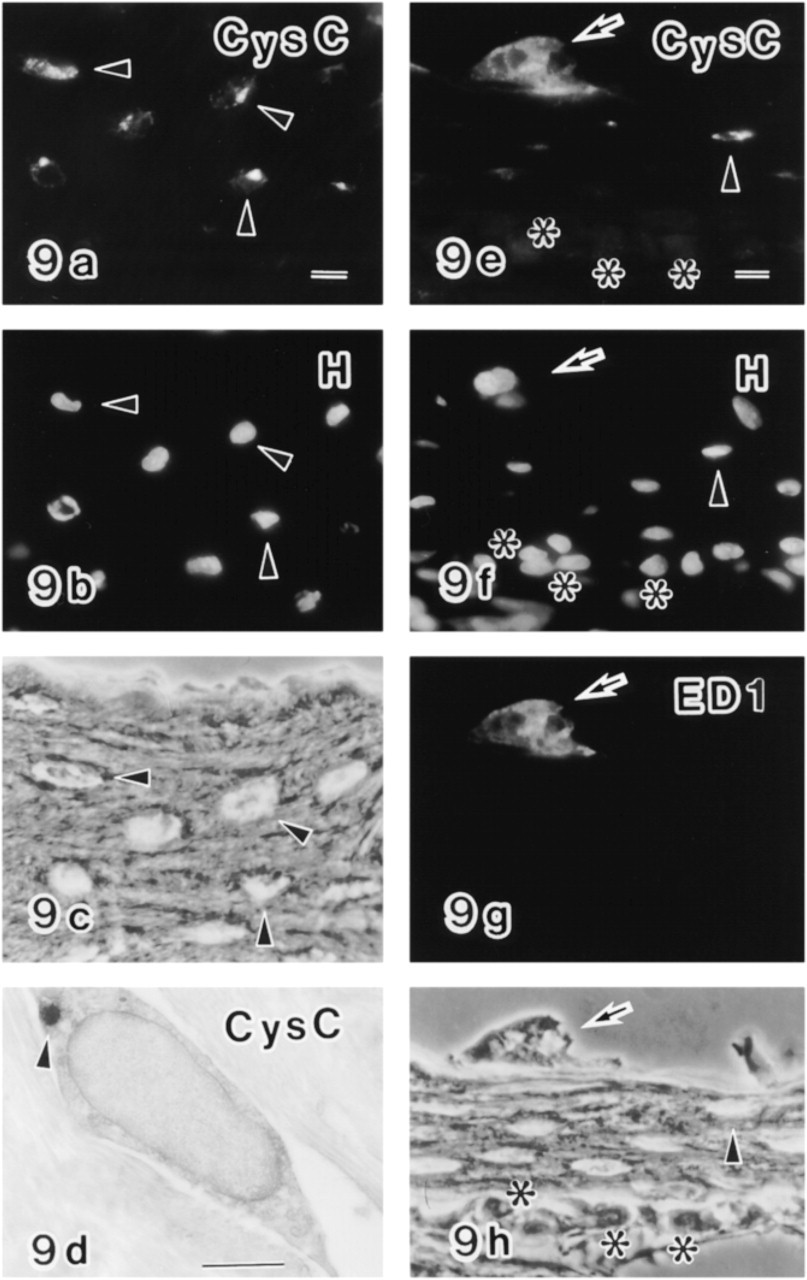
A double labeled section of the enamel organ of the early maturation zone (MAT) labeled with anti-cystatin C (CysC) (
OX6 immunoelectron microscopy revealed that OX6+ dendritic cells and OX6– non-epithelial cells with a “dendritic shape” that were morphologically indistinguishable from OX6+ cells were observed in the transition zone (Figures 2a and 2b). These OX6–non-epithelial cells are considered to be dendritic cells. OX6+ dendritic cells with a highly complex shape were observed in the maturation zone (Figures 2c and 2d).
Anti-cystatin C Labels Immature and Maturing Dendritic Cells in Enamel Organs
In the secretion zone, anti-cystatin C+ and OX6– cells were sparsely distributed in the enamel organ (Figures 3a, 3c, and 3e). These cells had ovoid profiles and were preferentially located in the stellate reticulum (Figure 3a). Most of these cells were distributed in the intercellular spaces of the stellate reticulum and fewer cells were attached to the inner surface of the blood vessels in light microscopy of Epon-embedded semi-thin sections. To determine the nature of the cells positive for anti-cystatin C, conventional electron and correlated anti-cystatin C immunoelectron microscopy were performed. Conventional electron microscopy revealed that these ovoid cells had a relatively smaller area of cytoplasm, with sparse cytoplasmic organelles and a greater proportion of heterochromatin in the nuclei, and thus have a monocyte-like appearance (Figure 6f). These cells exhibited no desmosomes or tonofilaments (Figure 6f). By Immunoelectron microscopy showed the labeled structures to be endoplasmic reticulum, including the nuclear membrane and vesicles of various sizes (Figure 6a). To confirm the non-epithelial origin of anti-cystatin C-positive cells, double labeling experiments with anti-desmoplakin 1 and 2 for desmosomes and anti-cystatin C antibodies were performed. The results showed that the anti-cystatin C-positive cells lacked anti-desmoplakin 1- and 2-positive fluorescent dots at their periphery (Figures 6g and 6h).
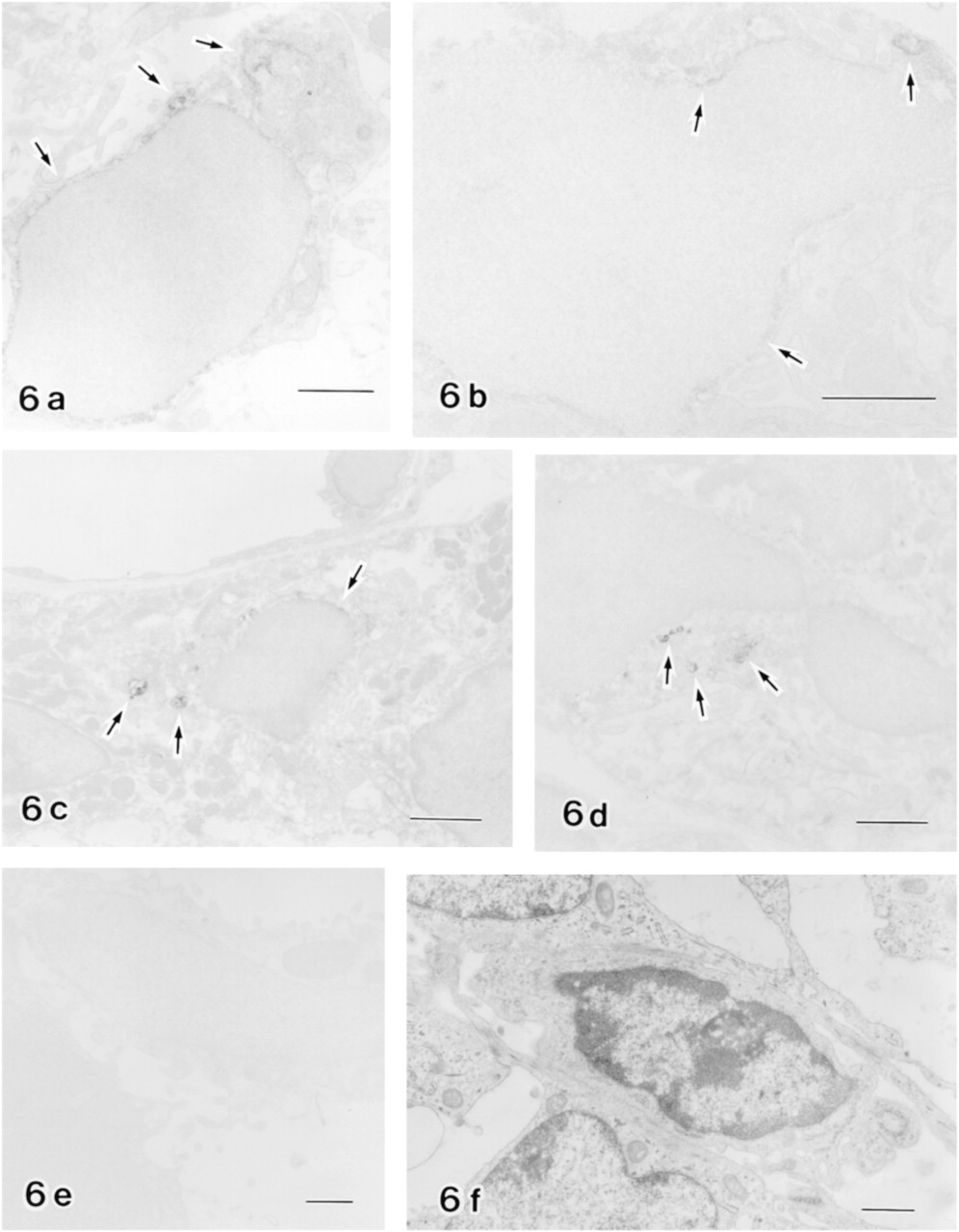
Dendritic cells in enamel organs. Immunoelectron micrographs of dendritic cells in the secretion zone (
In the transition zone, an increased number of anti-cystatin C+ cells were distributed in the papillary layers (Figures 3b and 4a). Some anti-cystatin C+ cells were also positive for the OX6 antibody (Figures 4a, 4d, and 4g), but the labeling intensity of OX6 was weak (Figure 4d) compared with OX6+ cells in the maturation zone (Figures 4e and 4f). Some of the anti-cystatin C+ cells were negative for OX6 (Figures 3b, 3d, and 3f). The structures labeled by anti-cystatin C on an electron microscopic level were endoplasmic reticulum and unknown vesicles (Figure 6b).
In the maturation zone, a number of OX6+ dendritic cells were distributed in the enamel organ. Most were also positive for the anti-cystatin C antibody in double labeled specimens (Figures 4b, 4c, 4e, 4f, 4h, and 4i), although some were singly positive. Double positive cells were present until the late maturation zone. Double labeling of the enamel organ in the maturation zone with anti-cystatin C and ED1 showed that two different antigens were co-localized (Figures 5a and 5b). The anti-cystatin C antibody labeled endoplasmic reticulum and some unknown vesicles of non-epithelial cells in the enamel organ (Figures 6c and 6d). Control sections without anti-cystatin C showed no specific labeling (Figures 5c, 5d, and 6e).
Using double labeled sections with anti-cystatin C and OX6, the number of positive cells per unit length of the enamel organ along the longitudinal axis of the incisor was counted. Based on a comparison between the secretion zone and the transition and maturation zones, anti-cystatin C-positive cells increased moderately in the more advanced zone (p < 0.01, Student's t-test), whereas OX6+ cells were rarely seen in the secretion zone but drastically increased in the transition and maturation zones (p < 0.001) (Figure 7).
Anti-cystatin C Labels Ameloblasts, Osteocytes, and Osteoclasts
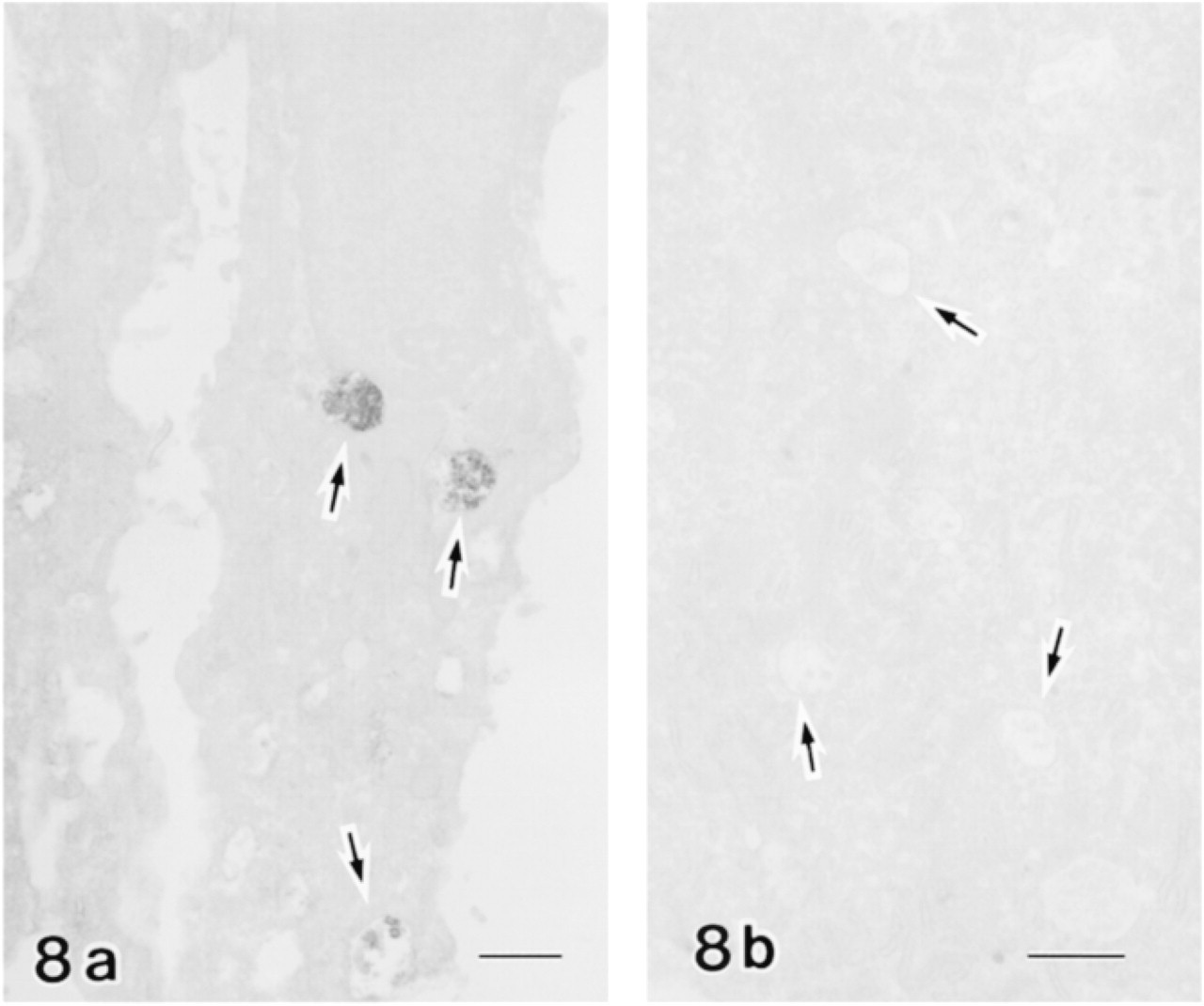
Anti-cystatin C+ dots were observed in the supranuclear region of ameloblasts in the late secretion to early maturation zones but not in the late maturation zone (Figures 3 and 4). The organelles labeled were multivesicular bodies (Figure 8). In alveolar bone covering an incisor, older osteocytes and osteoclasts were positive for anti-cystatin C, but osteoblasts and newly formed osteocytes were only faintly labeled (Figure 9). Some unknown vesicles in the osteocytes were labeled by anti-cystatin C in electron microscopy (Figure 9d). Osteoclasts were double labeled with anti-cystatin C and ED1, but osteocytes and osteoblasts were not labeled by ED1 (Figures 9e–9h).
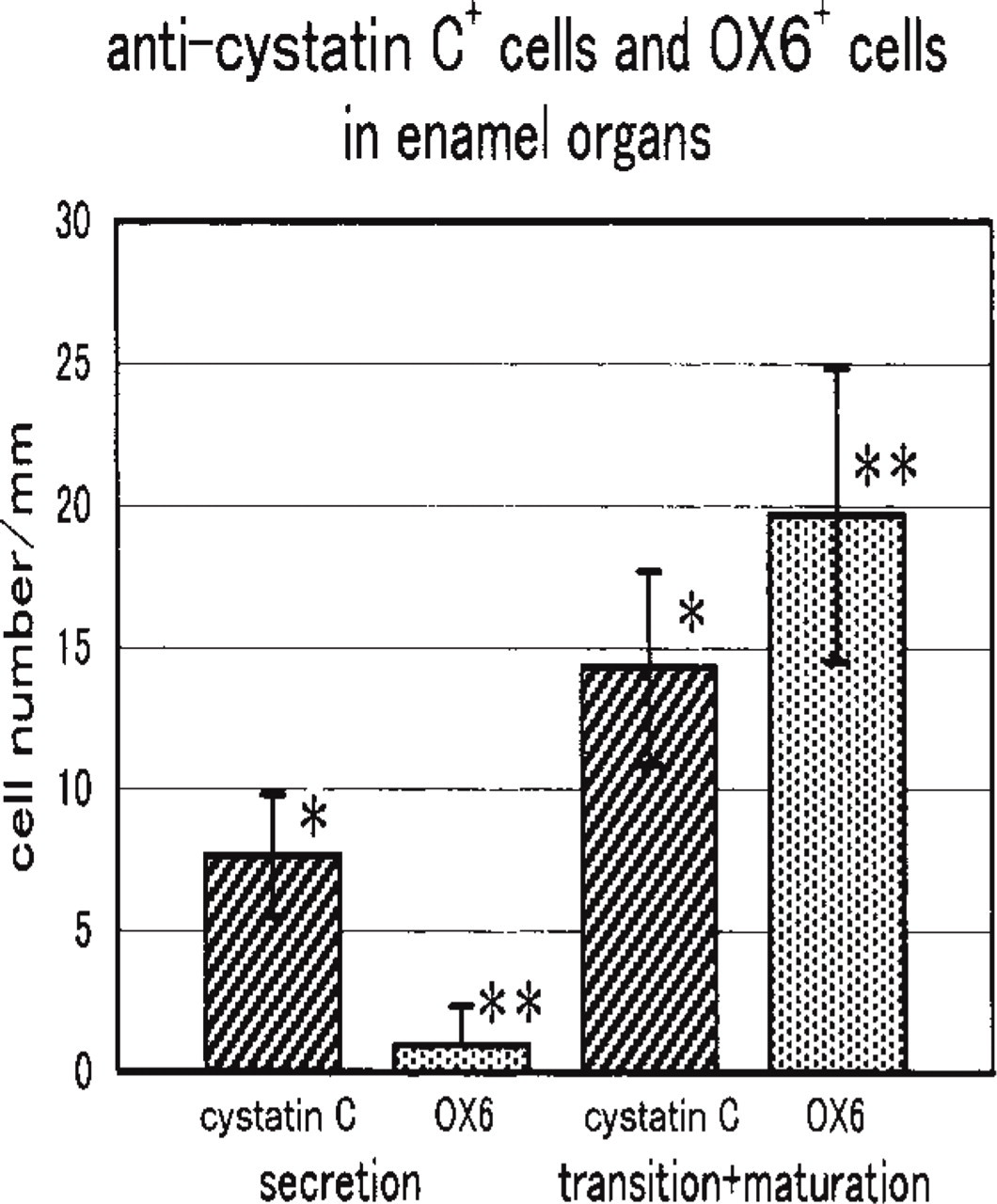
Numerical analysis of anti-cystatin C+ cells and OX6+ cells in the enamel organ of the secretion zone and transition zone plus early maturation zone. Cell number was counted along the long axis of incisors using sections labeled doubly with anti-cystatin C and OX6. Anti-cystatin C+ cells increase from the secretion zone to the transition plus early maturation zone, whereas OX6+ cells are rare in the secretion zone and increase drastically in the transition plus early maturation zone. Bars show standard deviation. ∗p < 0.01; 7 p < 0.001 (Student's t-test).
Discussion
Dendritic cells are the most professional and specialized antigen-presenting cells. They are distributed in a variety of tissues in vivo, stimulate quiescent, naive, and memory B- and T-lymphocytes and induce tolerance by deletion of self-reactive thymocytes and anergy of mature T-cells (Banchereau and Steinman 1998; Mellman et al. 1998). Although Langerhans cells in the skin were visualized in the nineteenth century, characterization of the dendritic cells began only 25 years ago (Banchereau and Steinman 1998). It has been shown that the dendritic cells are related to blood monocytes and tissue macrophages but are clearly from a distinct cell lineage (Shortman and Maraskovsky 1998).
Mature dendritic cells are stellate cells with long spiny or sheet-like extensions and exhibit high surface MHC Class II molecules and low endocytic activity, whereas immature dendritic cells exhibit high intracellular MHC Class II molecules and high endocytic activity in vitro (Banchereau and Steinman 1998). However, intracellular MHC Class II antigens are difficult to detect in tissues in vivo. The ultrastructure of the dendritic cells exhibits stellate morphology, eccentric irregular nuclei, and cytoplasm with ruffled projections. The cytoplasm contains phagosomes, multivesicular bodies, small vesicles, and a less-developed Golgi apparatus (Matsuno et al. 1996). The peripheral cytoplasm looks clear because there are only few cell organelles in this area (Sato et al. 1998).
Dendritic cells undergo maturation via antigen peptide loading on MHC Class II molecules in specialized MHC Class II-rich compartments, and present the complexes on the cell surface. To form antigen–MHC Class II complexes, both antigen and pro-formed MHC Class II must be cleaved by lysosomal proteases, a candidate for which is cathepsin S (Pierre and Mellman 1998). Proteolytic activities of the proteases in immature and mature dendritic cells have been reported to be regulated by endogenous protease inhibitors such as cystatin C. In the immature state, cystatin C inhibits cathepsin S activity, and in the mature state a decreased amount of cystatin C liberates its activity, whereas the amount of cathepsin S is constant in immature and mature dendritic cells (Pierre and Mellman 1998).
In this study, dendritic cells in the enamel organ of the secretion, transition, and maturation zones were examined by using an anti-cystatin C antibody and an OX6 antibody directed to MHC Class II antigen.
It has been shown that during amelogenesis there are “macrophages” or macrophage-like cells in the enamel organ of secretion, transition, and maturation zones (Jessen and Moe 1972; Josephsen and Fejerskov 1977; Kallenbach 1970, 1978; Nishikawa and Joseph-sen 1987; Nishikawa and Sasaki 1996). Takano et al. (1996) revealed that they are dendritic cells and that a number of OX6+ dendritic cells are distributed in the enamel organ of the maturation zone. This study and a previous report (Nishikawa and Sasaki 1999a; Takano et al. 1996) revealed that because there is no macrophage marker (ED2-reactive antigen), there are no OX6+ epithelial cells in the enamel organ, and because there is no B-cell marker, OX6+ cells observed in the enamel organs of both transition and maturation zones were dendritic cells.
To determine whether or not these dendritic cells are a mature form, anti-cystatin C and OX6 double labeling was performed. Anti-cystatin C-positive cells were observed in the enamel organs of the secretion, transition, and maturation zones by light microscopic immunocytochemistry. Unexpectedly, in the enamel organ of the secretion zone anti-cystatin C-positive cells were present and were negative for OX6 antibody, whereas in the enamel organs of the transition and maturation zones some of the anti-cystatin C-positive cells were also double positive for OX6 antibody. Double positive cells in the transition and maturation zones may be maturing dendritic cells instead of fully matured cells, because matured Langerhans cells in epithelia have mostly lost cystatin C antigenicity (Pierre and Mellman 1998).
Anti-cystatin C immunoelectron microscopy of early maturation ameloblasts (
On the basis of conventional electron microscopy and anti-cystatin C immunoelectron microscopy, anti-cystatin C+ cells in the secretion zone also appear to be immature dendritic cells that do not yet express cell surface MHC Class II antigens. They do not contain desmosomes or tonofilaments, thus demonstrating their non-epithelial origin. Double labeling of the enamel organ with anti-cystatin C and anti-desmoplakin 1 and 2 antibodies for desmosomes confirmed that the anticystatin C+ cells were of non-epithelial origin. In anti-cystatin C immunoelectron microscopy, the organelles labeled were the nuclear membrane, the endoplasmic reticulum, and some vesicles. These vesicles should be phagolysosomes, because ED1 co-localized with anti-cystatin C (Dijkstra et al. 1985). Cystatin C is also known to be synthesized by macrophages (Chapman 1991), which secrete it via a constitutive pathway. Therefore, in tissues containing macrophages as well as dendritic cells, it is difficult to evaluate the nature of anti-cystatin C+ cells. Fortunately, in this study, the enamel organ did not contain macrophages. The results of the present study suggest that in the secretion zone, immature dendritic cells are present, possibly penetrate the enamel organ from nearby blood vessels, endocytose antigens in the transition zone, and become maturing cells in the maturation zone. The increase of OX6+ and anti-cystatin C double positive dendritic cells and several anti-cystatin C single positive cells in the maturation zone may further imply penetration of a new population of dendritic cells into the enamel organ. Alternatively, several types of non-epithelial cells may be present in different zones during amelogenesis.
During enamel maturation, a considerable number of ameloblasts are lost, as well as the transition zone (Smith and Warshawsky 1977). Furthermore, atrophic cytoplasm encircled by the annulated gap junctions is extensive in the papillary layer of the maturation zone (Kallenbach 1978; Sasaki and Garant 1986). In the transition zone, the dendritic cells phagocytose apoptotic cell fragments and endocytose anti-amelogenin-positive material (Nishikawa and Sasaki 1996, 1999a). The dendritic cells may be related to reconstitution of the enamel organ during amelogenesis.
An unexpected result was the presence of anti-cystatin C+ vesicles in the ameloblasts. These vesicles appear to be multivesicular bodies. Considering that ameloblasts develop a lysosomal system during the period of secretion to the maturation zone and therefore exhibit cathepsin B, one of the cysteine proteases (Smith 1979; Al Kawas et al. 1996), it is reasonable to assume that cystatin C regulates cathepsin B in the lysosomal body, especially during the secretory stage. Calvarial bone tissues, including osteocytes and osteoblasts, and an osteoblastic cell line are shown to produce cystatin C and are believed to inhibit the activity of osteoclastic proteolytic enzymes (Lerner et al. 1997). The distribution pattern of anti-cystatin C in the alveolar bone observed in this study is consistent with this line of evidence. Osteoclasts have been shown to express cysteine proteases and those such as cathepsin L are believed to participate in bone matrix degradation (Tezuka et al. 1994; Kakegawa et al. 1993). The presence of anti-cystatin C in the osteoclasts suggests that this inhibitor is involved in regulation of bone resorption.
Immunocytochemical localization of anti-cystatin C (CysC) (
In conclusion, immature dendritic cells positive for anti-cystatin C may already be present in the enamel organ of the secretion zone, may increase in number in the transition and maturation zones, and may also begin to express MHC Class II antigens at their plasma membranes. Some immunological responses may be strongly related to the duration of amelogenesis, and those remain to be clarified.
Footnotes
Acknowledgements
Supported in part by a Grant-in-Aid for scientific research (no. 11671824 to SN) from the Ministry of Education, Science, Sports and Culture of Japan.