
Abstract
Immunohistochemistry employing a new hapten antibody that detects the SKL sequence and its variants of the PTS1 C-terminus of peroxisomal enzymes was attempted to visualize peroxisomes across species. Rabbits were immunized with the SKL sequence coupled with KLH, between which an arm molecule was interposed. IgG fractions of antisera were affinity-purified against the hapten and employed for immunochemical analyses and immunoelectron microscopy. The specificity of the antibody was examined by immunoblot analyses for various purified enzymes of rat liver peroxisomes and by dot-blot analyses inhibited by SKL peptide and its variants. Various animal and plant tissues were subjected to immunoelectron microscopy with the protein A-gold technique. The antibody reacted with various enzymes in the peroxisome with the SKL motif. The affinity of the antibody for tripeptides, which varied depending on their structures, was higher for SKL than for its variants. Hepatic and renal peroxisomes of vertebrates, peroxisomes in the fat body of an insect, and the cotyledon of a plant were visualized by immunoelectron microscopy. Immunohistochemistry employing this SKL antibody may provide specific staining that can detect peroxisomes across different species.
P
Histochemical detection of peroxisomes has been accomplished most commonly by the DAB reaction for catalase (Fahimi 1969) and by cerium methods for various oxidases, including D-amino acid oxidase, L-α-hydroxyacid oxidase, and urate oxidase (van den Munckhof 1996). Immunoelectron microscopy, especially the protein A-gold technique, successfully visualized the locations of various kinds of enzymes and proteins in peroxisomes and clarified the relationship between the ultrastructure and its constituents (Usuda et al. 1995). Immunohistochemical detection requires an antibody for each kind of antigen to be visualized. Because distinct species differences exist for metabolic systems and for most peroxisomal proteins in general, it would be expedient to be able to apply the antibody to other species than that from which the antigen was purified.
Recently, the transport system of peroxisomal matrical enzymes, i.e., how they translocate through the peroxisomal membrane after they are synthesized on free ribosomes (Miura et al. 1984; Lazarow and Fujiki 1985; Osumi and Fujiki 1990), has come to be understood in principle. The mechanism of transport was first clarified in vivo and in vitro in firefly luciferase and rat fatty acid β-oxidation enzymes (Gould et al. 1989; Miyazawa et al. 1989). At least two types of targeting signal exist for matrix enzymes, i.e., the extreme carboxyl end of peroxisomal protein, the Ser-Lys-Leu-COOH (SKL) sequence, and the internal position of the N-terminus of the presequence (Purdue and Lazarow 1994). The former is called the PTS1 signal and the latter the PTS2 signal. Proteins with these signals are transported into peroxisomes by binding to their respective PTS receptors or to docking proteins of the peroxisome translocation apparatus (Rachubinski and Subramani 1995). It now appears that this mechanism of transport of peroxisomal matrix proteins is conserved among eukaryotes, including animals and plants (Gould et al. 1989, 1990; Keller et al. 1991; Miura et al. 1992; Gietl 1996).
The new antibody was raised against the tripeptide of SKL. The antibody was characterized for its specificity, and immunoelectron microscopic detection of peroxisomes across different species was attempted with it.
Materials and Methods
Tissues
Electron microscopic analysis was conducted on liver tissues of various animals: Wistar rats (Rattus rattus), slider tortoise (Pseudemys scripta collirostris), bullfrog (Rana catesbeiana), and rainbow trout (Salmo gaird nerii), the fat body of a cicada (Oncotympana maculaticollis), and the cotyledon of a soybean (Glycine max), and on rat and bovine (Bos taurus) kidney tissues. Rats were fed a normal or a 1% DOP diet. Peroxisome fractions obtained by sucrose gradient from normal and DOP-treated rat liver tissues were prepared for immunoblot analysis (de Duve et al. 1955).
Preparation of Antibodies
A peptide, cysteine-aminocaproic acid-serine-lysine-leucine, was synthesized by the solid-phase method, purified by reverse-phase HPLC, and the sequence confirmed by amino acid analysis. SKL is the C-terminus of the deduced amino acid sequence of acyl-CoA oxidase, based on the rat gene (Miyazawa et al. 1987). Aminocaproic acid (Acp) was employed as a hydrophobic arm molecule with a carbon chain length of C5. The purified peptide was coupled to keyhole limpet hemocyanin (KLH) through cysteine at the -SH side chain by m-maleimidobenzoyl-N-hydroxysaccinimid ester (Royer-Pokora et al. 1986) so as not to interfere with -COOH and -NH2 groups of amino acids. New Zealand White rabbits were immunized with the antigen KLH-SKL using complete and incomplete Freund's adjuvant. The IgG fraction of the antisera was affinity-purified on CH-Sepharose coupled with the polypeptide. The antibody was designated as SKL-Ab.
The antibody for acyl-CoA oxidase was prepared by immunizing rabbits with its protein purified from rat liver tissues fed a 1% DOP diet as previously described (Osumi et al. 1980).
Preparation of Peroxisomal Enzymes and Peptides
Acyl-CoA oxidase (AOX), enoyl-CoA hydratase/3-hydroxyacyl-CoA dehydrogenase (PH), 3-ketoacyl-CoA thiolase (PT), urate oxidase (UOX), D-amino acid oxidase (DAOX), and catalase (CAT) were purified from liver or kidney tissues of normal or DOP-treated rats (Watanabe and Suga 1978; Furuta et al. 1980, 1986; Miyazawa et al. 1980, 1989; Osumi et al. 1980; Usuda et al. 1986). Firefly luciferase was purchased from Boehringer Mannheim (Tokyo, Japan). The C-terminus of each enzyme is as follows, AOX, PH, and firefly luciferase, SKL; UOX, SRL; DAOX,SHL; CAT, ANL; and PT, PGN. Peptides of SKL, SRL, SHL, AKL, ANL, and PGN were synthesized by the solid-phase method, purified by reverse-phase HPLC, and confirmed by amino acid analysis. Those peptides other than PGN are examples of the C-terminus of the deduced amino acid sequence of rat peroxisomal enzymes with PTS1 C-termini (Furuta et al. 1986; Ishii et al. 1987; Miyazawa et al. 1987; Reddy et al. 1988; Gould et al. 1990; Jiang et al. 1997; Konno 1998). PGN is the C-terminus of the deduced amino acid sequence of PT, which does not possess PTS1 in the C-terminus but has the PTS2 targeting signal in the N-terminus (Hijikata et al. 1987; Tsukamoto et al. 1994).
Immunoblot Analyses
The purified rat peroxisomal enzymes, AOX, PH, PT, UOX, DAOX, and CAT, firefly luciferase, and peroxisome fractions were subjected to 12% SDS-PAGE electrophoresis and immunoblot analysis was performed as previously described (Johkura et al. 1998), using SKL-Ab and secondary antibody coupled with horseradish peroxidase. The reaction was visualized using 4-chloro-1-naphthol in the presence of hydrogen peroxide for 10 min. Immunoblot of AOX was also performed using its antibody.
Inhibition Assay
Dot-blot analyses were performed by inhibiting the reaction between SKL-KLH antigen and SKL-Ab with the synthetic peptides. The synthetic tripeptides were dissolved in 10 mM potassium phosphate, (pH 7.5)/150 mM NaCl. The KLH-SKL dissolved in 10 mM potassium phosphate (pH 7.5)/150 mM NaCl/1% SDS was applied at a concentration of 50 ng/ml to small (1 cm × 1 cm) squares of nitrocellulose membrane as focused spots in a volume of 1 ml. Each nitrocellulose square was stained by the same procedure as for the immunoblot analysis. The squares were stained with the mixture of SKL-Ab (2.5 mg/ml) and the tripeptides at various concentrations. After the staining, each square was dried and examined by densitometry. The images were digitized with a computer (Apple Computer; Cupertino, CA) equipped with a scanner (Hewlett Packard Japan; Tokyo, Japan). They were analyzed with Ultimage software (Image and Measurement; Tokyo, Japan). The density was corrected for background density measured outside each spot and was expressed relatively as percent inhibition in terms of the control, i.e., the background density was set as 100% inhibition and the density of the reaction products obtained in the absence of any tripeptide was set as 0% inhibition. The percent inhibition was plotted against the log concentration of tripeptides (Kabat 1961) and the approximate line was calculated.
Immunoelectron Microscopy
Tissue processing and immunostaining were done basically as reported previously (Bendayan and Reddy 1982; Usuda et al. 1988a). Small pieces of tissue (approximately 1 mm × 0.5 mm × 0.5 mm) were fixed in 4% paraformaldehyde/0.1% glutaraldehyde/100 mM sodium phosphate, pH 7.4, at 4C, for 12 hr. After rinsing in a cold solution consisting of 100 mM lysine/100 mM sodium phosphate (pH 7.4)/150 mM NaCl and dehydration in a graded series of cold ethanol, they were embedded in Lowicryl K4M at −20C (Roth et al. 1981). Ultrathin sections were immunostained with SKL-Ab and a protein A-gold solution of 15-nm gold particles (EY Laboratories; San Mateo, CA). Specimens were observed with a Hitachi H-700 electron microscope at an accelerating voltage of 150 kV after staining with uranyl acetate and lead citrate.
Results
Immunoblot Analysis
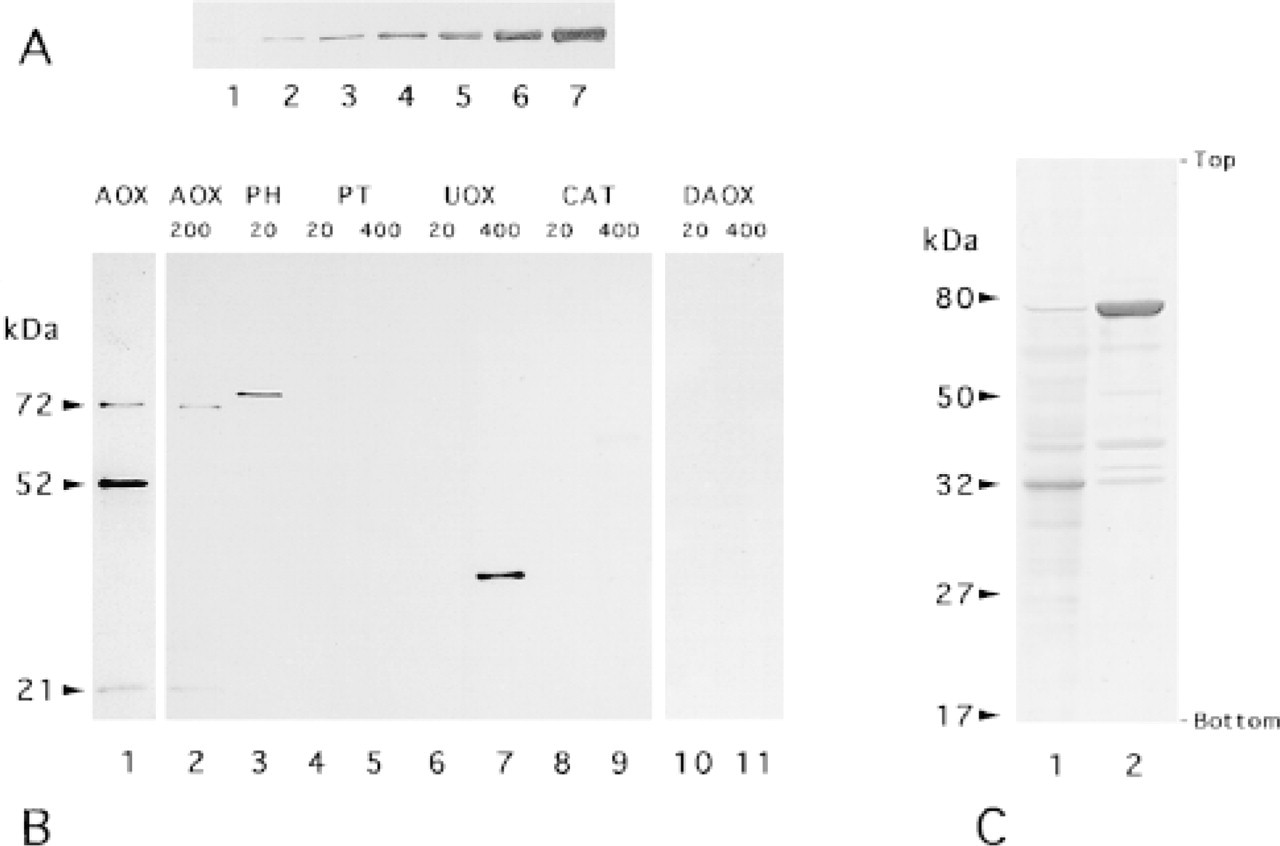
(
Figure 1A shows the detection of various amounts of firefly luciferase by SKL-Ab. The signal was slight with an amount of 8 ng and clearly visible with amounts over 16 ng. Figure 1B shows the reactivity of SKL-Ab with various rat peroxisome-specific enzymes under the same conditions as for luciferase. AOX consists of three subunits, Subunit A (MW 72 kD), Subunit B (MW 51 kD), and Subunit C (MW 21 kD). Whereas Subunits A and C were detected by SKL-Ab when 200 ng was applied, Subunit B was not. In contrast, all three subunits of AOX were detected by anti-AOX antibody. PH was detected clearly at an amount of 20 ng. UOX was not detectable with 20 ng but was detected clearly with 400 ng. CAT was not detectable with 20 ng and was detected very weakly with 400 ng. PT and DAOX were not detected when 20 ng or 400 ng was applied. The immunoblots of peroxisome fractions of normal and DOP-treated livers showed more than 15 signals. The most distinct signals corresponded to the molecular weight of UOX and PH for fractions of normal and DOP-treated livers, respectively (Figure 1C).
Inhibition Assay
Figure 2A shows the results of the inhibition of the reaction between SKL antigen and SKL-Ab by synthetic tripeptides. The density of the reaction products decreased considerably in parallel with the increase in the concentration of SKL. The reaction became negative at the maximal concentration of 1000 μg/ml. The density of the reaction product also decreased along with the increase in the concentration of SRL, but the reaction products were still visible at a concentration of 1000 μg/ml. In contrast, the decrease in the density of the reaction products was not discernible after addition of PGN. Figure 2B shows the relationship between the concentration of tripeptides and percent inhibition obtained from measuring the result of the dot-blot analysis. The percent inhibition was plotted against the log concentration of tripeptides. The slopes of the approximate lines, indicating the effect of the inhibition, were 31, 26, 25, 18, 9.9, and 1.8 for SKL, AKL, SRL, SHL, ANL, and PGN, respectively. The larger the value, the stronger was the inhibition of the reaction by a peptide.
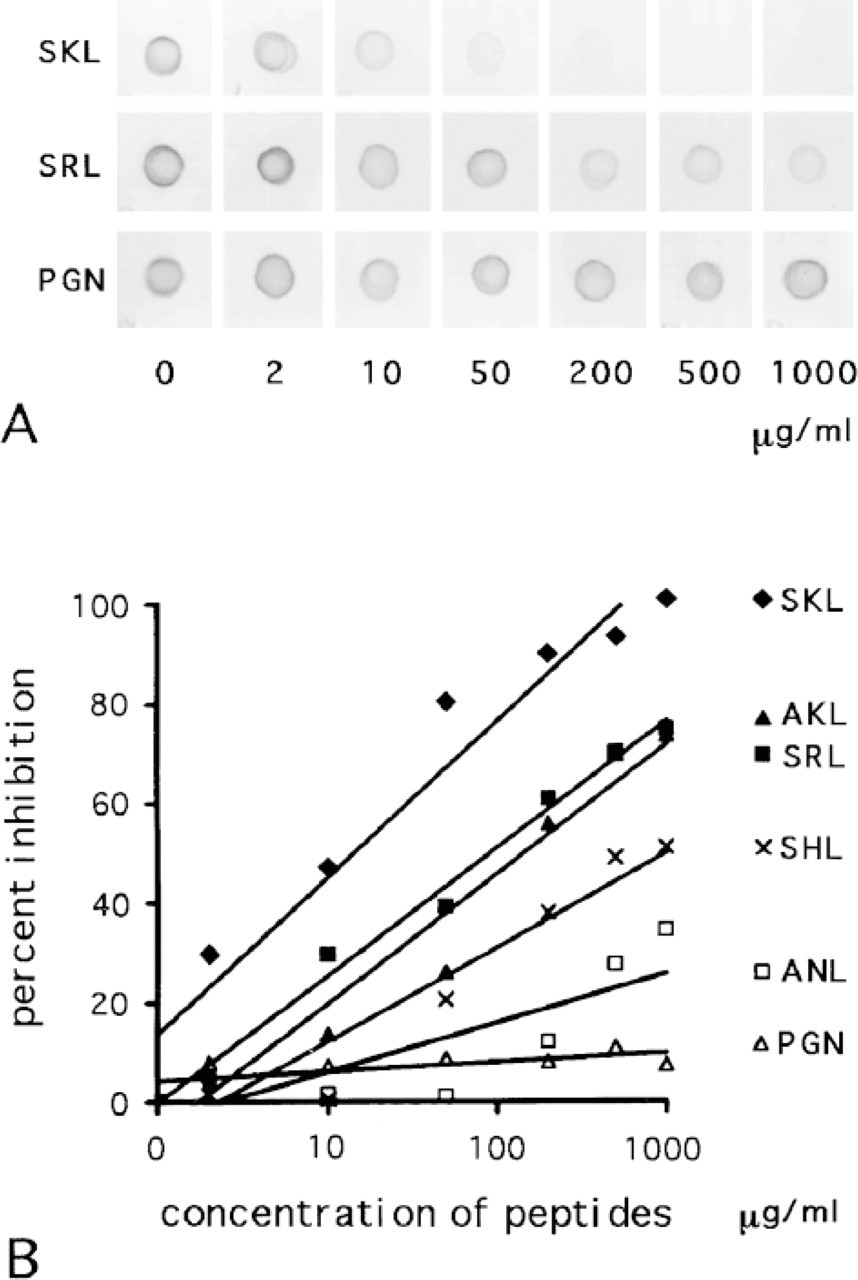
(
Immunoelectron Microscopy
Figures 3–5 illustrate the reactivity of SKL-Ab to various ultrastructures. Electron micrographs of peroxisomes of normal and DOP-treated rat liver tissues stained by the protein A-gold technique with SKL-Ab show that gold particles are located over peroxisomes and that other cell structures are negative for the reaction (Figure 3). The core and the matrix of the peroxisomes are reactive in normal hepatocytes (Figure 3A) and in a DOP-treated hepatocyte (Figure 3B). The labeling density increased with the administration of DOP. In proximal tubule epithelial cells of DOP-treated rat kidney tissues stained by the protein A-gold technique, gold particles were located solely over peroxisomes, with other ultrastructures, such as the cell membrane, microvilli, absorption vesicles, lysosomes, mitochondria, and nucleus, being consistently negative for the staining (Figure 4A). High-power magnification showed that most gold particles were located over the peripheral area, but not over the central area, in the matrix of peroxisomes (Figure 4B). In the bovine renal peroxisome, the core and the marginal plates were distinctively labeled (Figure 4C). Figure 5 summarizes the reactivity of peroxisomes of various vertebrate hepatocytes, the insect fat body, and the plant cotyledon. Electron micrographs showed spherical cell organelles, i.e. peroxisomes, to be immunoreactive to SKL-Ab in chicken liver (Figure 5A), tortoise liver (Figure 5B), frog liver (Figure 5C), fish liver (Figure 5D), insect fat body (Figure 5E), and soybean cotyledon (Figure 5F). Other structures were immunonegative for SKL-Ab in the liver, the fat body, and the cotyledon as far as was observed.
Discussion
Data are accumulating on the cDNA sequences of peroxisomal matrix enzymes and their deduced amino acid sequences. Most are now known to possess the SKL sequence and its variants (Gould et al. 1989; Miura et al. 1992), as exemplified for rat enzymes used in the present study, e.g., SKL:AOX, PH; SRL:UOX; SHL:DAOX; ANL:CAT (Miyazawa et al. 1987; Reddy et al. 1988; Rachubinski and Subramani 1995; Purdue and Lazarow 1996; Konno et al. 1998), all of which are believed to function as transport signals (Miura et al. 1992; Purdue and Lazarow 1994), although this is somewhat controversial in the case of CAT (Purdue and Lazarow 1996). The third enzyme of the peroxisomal fatty acid β-oxidation system, PT, does not have this tripeptide but has a transport signal near the N-terminus, which is cleaved after transport (Tsukamoto et al. 1994). The former is called the PTS1 signal and the latter PTS2 (Purdue and Lazarow 1994).
Immunoblot analyses of these purified enzymes showed that SKL-Ab detects peroxisomal enzymes but with different detection limits. Firefly luciferase, AOX, and PH were detected clearly by SKL-Ab when small amounts of the samples were applied in SDS-PAGE. UOX and CAT were detected when large amounts were applied, but PT and DAOX were not. Judging from the slopes of the approximate lines obtained in the inhibition test by various peptides, SKL is the strongest inhibitor, SRL and AKL are moderate, and SHL and ANL are weaker. PGN has almost no effect on the reaction between SKL peptide and SKL-Ab. Among the three subunits of AOX, Subunits A and C were reactive to the antibody but B was not. Luciferase, Subunits A and C of AOX, and PH have an SKL sequence at the C-termini. UOX has SRL, CAT has ANL, and DAOX has SHL at the C-terminus. The B subunit of AOX and PT do not have SKL or its variants at their C-termini. From these findings in the inhibition test and the immunoblot analysis, the SKL-Ab was shown to react with the SKL motif and its variants, most strongly with SKL itself but more weakly also with the variants SRL, AKL, SHL, and ANL. SKL-Ab could not detect DAOX with SHL but it did detect CAT with ANL, although SKL-Ab has a higher affinity SHL than ANL. This discrepancy may indicate that the structure of enzymes apart from their C-termini may have some effects on the reaction. These immunochemical data show that SKL-Ab specifically reacts with SKL tripeptide and its variants with different affinities. SKL-Ab could also detect signals in peroxisome fractions. These data indicated that SKL-Ab can detect these enzymes with SKL sequence and its variants in rat liver peroxisomes.
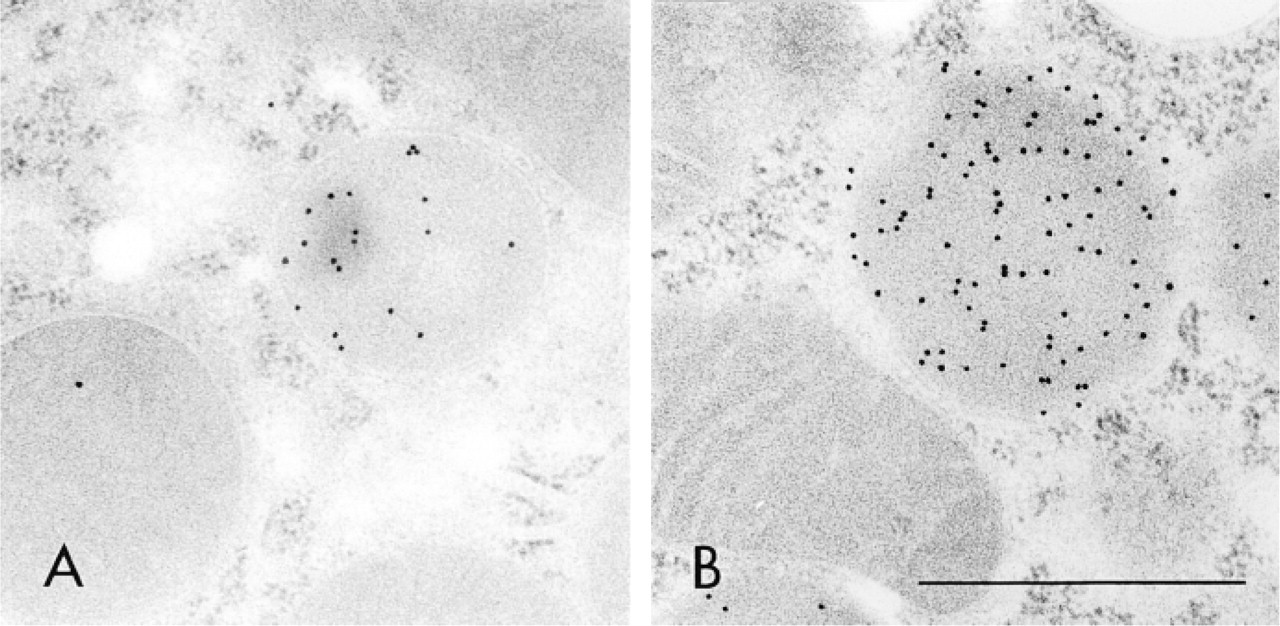
Electron micrographs of peroxisomes of normal and DOP-treated rat liver tissues stained by the protein A-gold technique with SKL-Ab. Hepatocytes of normal (
Other examples of the detection of various peroxisomal proteins employing differently designed antibodies for PTS1 and biotin-labeled PTS1 receptor have been reported. Antibodies detecting the SKL sequence, designated anti-PTS antibody, have been reported previously against synthetic peptides of CRYHLKPLQSKL and CRYLAGPHGSKL, which contain nine amino acids of the C-termini of AOX and PH, respectively (Gould et al. 1989, 1990). Immunoblot analyses employing these antibodies specifically detected 16 proteins containing the SKL sequence but not those with variants in the identification of PTS1-containing proteins containing SRL, AKL, and ANL in 50 μg of peroxisomal fractions, or these peptides. Immunoelectron microscopy on peroxisomes of various species employing ultrathin cryosections was made possible with these antibodies. Another approach to identifying proteins possessing PTS1 is the use of a biotinylated form of the human PTS1 receptor. This detected 20-25 proteins with a detection limit of 100-200 ng in 100 μg of peroxisome fractions by Western blotting (Fransen et al. 1996), but without obvious difference in the affinity with variants of the SKL sequence. These three procedures appear to differ in their sensitivity and specificity, perhaps in part as a result of differences in the procedures for blotting and visualization. A total of 50-100 μg of peroxisome fractions was needed for anti-PTS antibody and biotinylated PTS1-receptor, but SKL-Ab needed only 5 μg of peroxisome fractions to visualize peroxisomal proteins by immunoblot analysis. The detection procedure employing SKL-Ab in the present study may have a higher sensitivity than the other two. Anti-PTS antibody detected proteins only with C-terminus SKL, but the other two detected proteins with the SKL sequence and with its variants. The former has a higher specificity, but the other two, with broader specificity, may be more useful for detecting proteins with various kinds of PTS1, including SKL and its variants. All three procedures are useful for detecting PTS1-containing proteins, but with different characteristics in the detection.
SKL-Ab was reactive specifically with peroxisomes among various ultrastructures in rat hepatocytes and renal tubule epithelial cells by electron microscopy. The core and the matrix were reactive in normal and DOP-treated hepatocytes. The main constituent of the core is UOX (Usuda et al. 1988a) and that of the matrix of DOP-treated hepatocytes is PH (Bendayan and Reddy 1982). The peripheral matrix was more strongly reactive in the peroxisomes of the renal proximal epithelial cell than in the clear central matrix. The main constituent of the latter is D-amino acid oxidase and those of the former are other kinds of matrix proteins including CAT, fatty acid β-oxidation enzymes, and L-α-hydroxyacid oxidase (Bendayan and Reddy 1982; Yokota et al. 1985; Usuda et al. 1986; Nemali et al. 1988). In bovine renal peroxisomes, the antibody reacts most strongly with the core and the marginal plate. The main constituent of the former is UOX (Usuda et al. 1988b) and that of the latter is L-α-hydroxyacid oxidase (Zaar et al. 1989). The cores of both rat liver hepatocytes and bovine kidney are strongly immunoreactive, their main constituent being UOX, which has an SRL C-terminus. The immunoreactivity of UOX and the tripeptide of SRL is relatively lower than that of enzymes with the SKL C-terminus and the SKL peptide. There appears to be no conflict between the main constituents of each substructure and the immunochemical information on the affinity of SKL-Ab with enzymes and their C-termini. The C-terminus of rat D-amino acid oxidase is SHL, and the weak immunoreaction of the central area of rat renal peroxisomes may reflect the weak reaction of the antibody to this enzyme. Because the C-terminus of bovine L-α-hydroxyacid oxidase is not known and we do not have purified protein, we cannot say anything about the strong immunoreaction of the marginal plate. Considering the strong immunoreaction with the crystalline structure of the marginal plate, the magnitude of the immunoreactivity of this antibody may depend not only on the sequence but also on whether or not the crystalline structure is formed in peroxisomes.
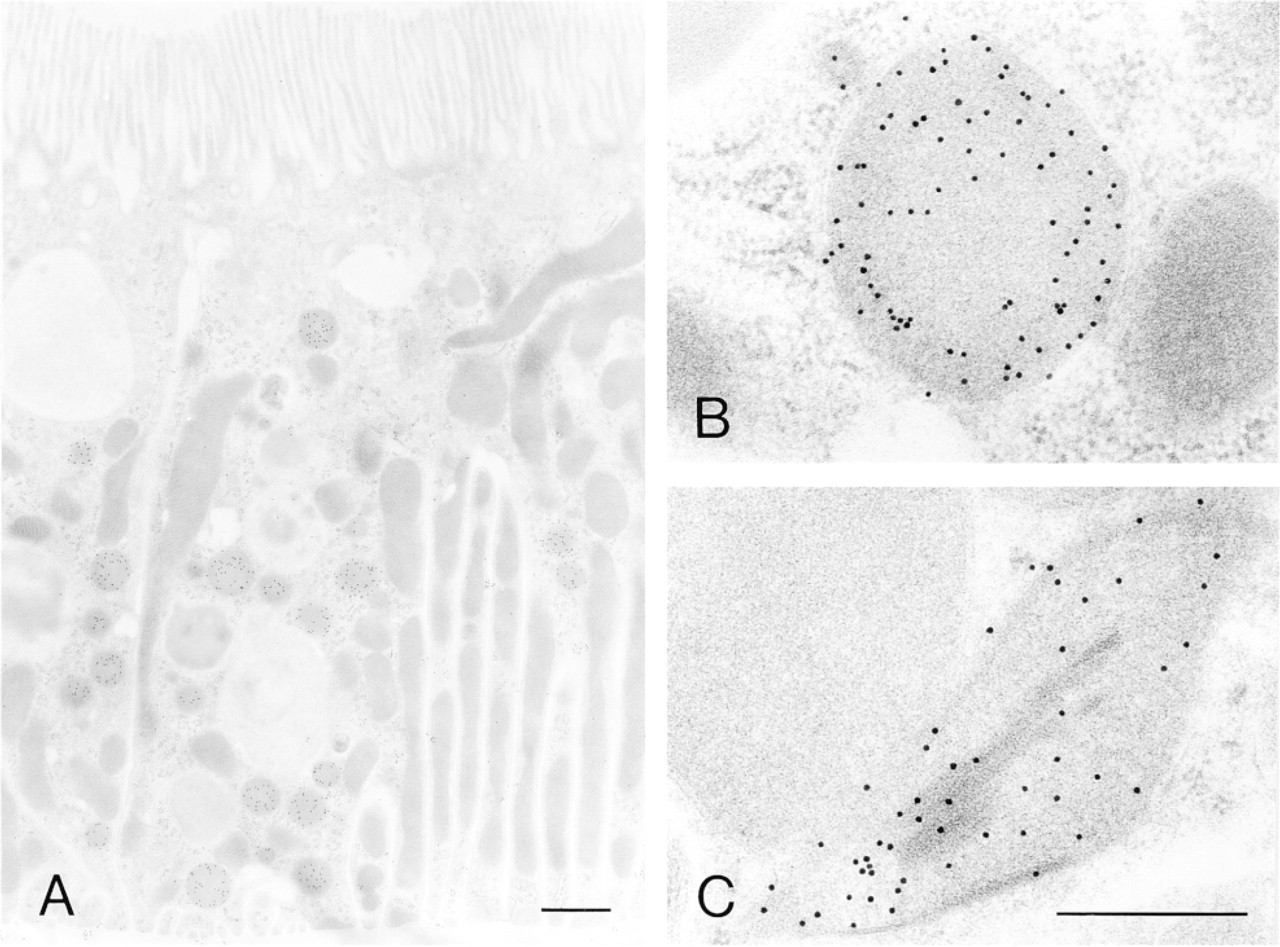
Electron micrographs of peroxisomes of rat and bovine kidney tissues stained by the protein A-gold technique with SKL-Ab. (
As examined by immunoblot analysis on purified proteins of rat peroxisomes and peroxisome fractions of rat liver tissues, by the inhibition assay, and by immunoelectron microscopy, this new antibody reacts with various enzymes with PTS1 targeting signal in peroxisomes, and SKL-Ab can be said to be a peroxisome-specific antibody, at least for rat tissues. Many enzymes have been purified and their cDNAs have been cloned from mammalian tissues, especially from rat tissues. However, no example exists for other vertebrates. The reaction of SKL-Ab to peroxisomal enzymes could not be described for birds, reptiles, amphibians, and fish based on the sequence of C-termini of the enzymes. Peroxisomal proteins with PTS1 signal might also be present in other species, because peroxisomal proteins of various species have the PTS1 signal (Gould et al. 1990; Subramani 1993; Purdue and Lazarow 1994). Moreover, spherical cell organelles with a single limiting membrane were specifically labeled with SKL-Ab in the hepatocytes of four species of vertebrate, in the fat body of an insect, and in the cotyledon of a plant. Because SKL-Ab could detect peroxisomes of various species including vertebrates, and an invertebrate, and a plant, it could be employed to detect peroxisomes across individual species to study the ontogenic and phylogenic development of peroxisomes.
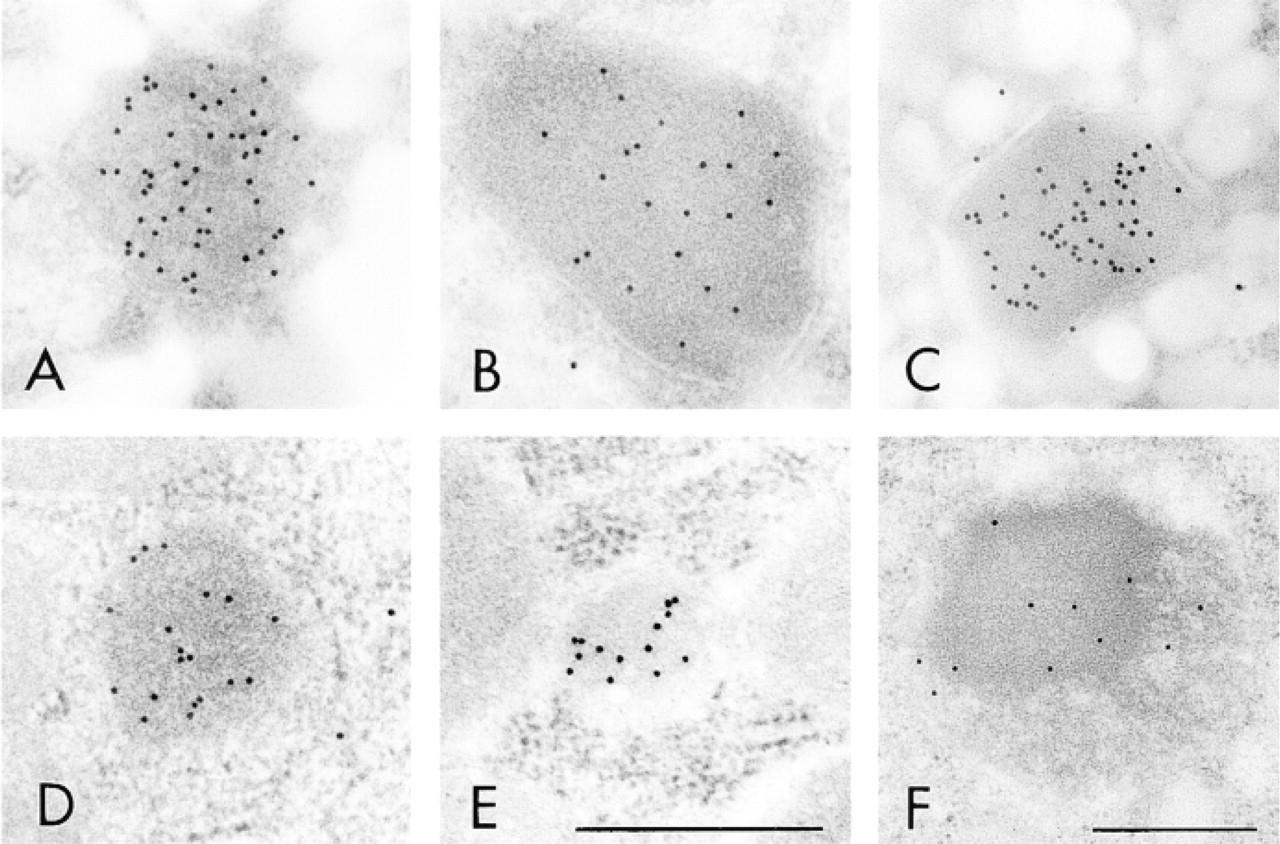
Electron micrographs of peroxisomes of various species stained by the protein A-gold technique with SKL-Ab. (
Footnotes
Acknowledgements
Supported in part by grants-in-aid for scientific research from the Ministry of Education of Japan.
We gratefully thank Dr T. Hashimoto (Department of Pathology, Northwestern University Medical School) for kind guidance in purifying enzymes and raising antibodies, and Mr Jun Usuda for obtaining tissue materials.