
Abstract
We outline here a protocol for high-resolution in situ localization of β-glucuronidase in murine tissues processed in glycol methacrylate (GMA). Murine tissues were first stained with 5-bromo-4-chloro-3-indolyl-β-D-glucuronic acid (x-gluc), followed by histological processing in GMA. Retention of the blue indigo reaction product after overnight incubations in x-gluc allowed high-resolution localization of β-glucuronidase activity by brightfield microscopy. When illuminated under darkfield, the x-gluc signal was enhanced, permitting detection even in cells with low-level enzyme activity. This technique offers for the first time a more sensitive enzyme histochemical method of detecting β-glucuronidase activity in animal tissues and also the opportunity to examine expression at high magnification.
β-GLUCURONIDASE is an acid hydrolase expressed at variable levels by virtually every cell in the vertebrate body (Paigen 1989). In its active form it participates in the degradation of glycaminoglycans (GAGs) by cleaving β-glucuronosyl residues at the nonreducing end of polysaccharides (Birkenmeier et al. 1989). Absence of this enzyme in humans results in the fatal lysosomal storage disease mucopolysaccharidosis type VII (MPS VII), or Sly syndrome (Sly et al. 1973, 1974). In 1989, a mutation in the murine β-glucuronidase gene complex (gusmps) was described that resulted in a phenotype similar to human MPS VII (Birkenmeier et al. 1989). Since then, the gusmps mouse has been used as a model system for studying this disease and various therapeutic modalities aimed at enzyme replacement (Sanes et al. 1986; Wolfe et al. 1990; Sands et al. 1995, 1997; Snyder et al. 1995; Levy et al. 1996; Naffakh et al. 1996; Vogler et al. 1996; Taylor and Wolfe 1997).
Biochemical, molecular, and cytochemical assays have been employed to comparatively examine β-glucuronidase activity in both wild-type and gusmps mice. Although many of these assays provide quantitative information, only the cytochemical assay provides information regarding the localization and differential expression of enzyme activity in situ. Currently available enzyme histochemistry (EHC) protocols for β-glucuronidase require tissues to be cryosectioned, resulting in poor to marginal microscopic resolution. Moreover, the cytochemical reaction product of the substrate naphthol-AS-BI-β-D-glucuronic acid, a red precipitate, is soluble in most organic reagents used for routine histological processing and, as a result, precludes the use of this substrate to pre-stain tissues before processing (personal observation). In this report we offer an alternative protocol which provides not only high-resolution localization of β-glucuronidase activity by brightfield microscopy but also enhanced detection by darkfield microscopy on the same tissue section. This technique is similar to that employed for the cytochemical demonstration of lacZ in transgenic mouse tissue (Sanes et al. 1986; Goring et al. 1987; Allen et al. 1988; Gossler et al. 1989; Lazik et al. 1997; Li et al. 1998) processed in 2-hydroxyethyl methacrylate or glycol methacrylate. Here we substitute the β-glucuronidase substrate 5-bromo-3-chloro-indolyl-β -D-glucuronic acid, or x-gluc, for x-gal, with modifications conducive to the pH optima of this enzyme. Therefore, any tissue can be stained by this method and subsequently processed in GMA with complete retention of the reaction product.
Materials and Methods
Frozen Section Enzyme Histochemistry
Adult H2bm1 mice (Jackson Laboratories; Bar Harbor, ME) were slowly perfused with 10 ml PBS, pH 7.4, followed by 10 ml 4% buffered paraformaldehyde, using a 27-gauge butterfly catheter. Spleen, liver, and brain tissues were then dissected into 5 × 5 × 3-mm portions and fixed for 2 hr at 4C. Specimens were then washed several times with PBS, cryopreserved in PBS/7% sucrose, and frozen in OCT compound (Sakura Finetek; Torrance, CA). All specimens were sectioned at 10 μm. Slides were then postfixed in a citrate-acetone-formaldehyde fixative for 30 sec at room temperature. EHC was carried out using a naphthol-AS-BI-β-D-glucuronic acid-based cytochemical kit (Sigma; St Louis, MO) per manufacturer's instructions and the slides were counterstained with methylene blue. The care and use of animals in this study followed the guidelines outlined by the university's Institutional Animal Care and Use Committee.
Glycol Methacrylate Enzyme Histochemistry
Mice were perfused and eviscerated, as described above, substituting 2% paraformaldehyde/0.2% glutaraldehyde and 0.02% IGEPAL (Sigma)/0.01% sodium deoxycholate for paraformaldehyde and postfixing tissues for only 1 hr. Tissues were washed in three changes of PBS at 4C and placed in x-gluc solution [5 mM potassium ferricyanide, 5 mM potassium ferrocyanide, 2 mM MgCl2, 0.02% NP40/0.01% sodium deoxycholate, and 1 mM x-gluc (Clontech; Palo Alto, CA) (from a 40 mg/ml stock prepared in DMSO) in PBS and pH adjusted to 5.2] and incubated at room temperature overnight on a low-speed shaker. The following day, the x-gluc-stained tissues were washed several times with PBS, then transferred to PBS/7% sucrose and placed on a rotator in a cold room for 6 hr. All subsequent steps were carried out on a rotator at 4C. Specimens were then removed and dehydrated in cold 70% ETOH for 4-5 hr, 95% ETOH overnight, and two changes of 100% ETOH each for 4 hr. Tissues were then infiltrated with GMA monomer per manufacturer's instructions overnight (Energy Beam Sciences; Agawam, MA). The following morning, GMA was replaced with two changes of 10:1 monomer to benzoyl peroxide-saturated 2-hydroxy ethanol, each for 4-5 hr. Tissues were then removed from the infiltration solution, oriented in 16 × 8-mm plastic embedding molds (Polysciences; Warrington, PA), and polymerized at approximately 0-1C overnight. The following day, plastic blocks were transferred to a vacuum dessicator to harden for several hours before sectioning. Blocks were cut at 2-4 μm on a Reichert-Jung JB-4 microtome with a Ralph knife. Sections were then placed on Superfrost/Plus microscope slides (Fisher Scientific; Pittsburgh, PA) and incubated on an 80C hotplate for 1-2 min to allow adequate attachment. Sections for brightfield microscopy were counterstained in 0.1% basic fuchsin for several seconds, washed in dH20, dehydrated, and mounted with Permount and Corning #1.3 glass coverslips (both from Fisher Scientific).
Microscopy
Photomicrographs were taken on a Nikon Optiphot-2 microscope. Brightfield photographs were taken with Sensia 100 slide film (Fujifilm; Tokyo, Japan). Darkfield photographs were taken with Ektachrome 64 tungsten slide film (Eastman Kodak; Rochester, NY). Slides were scanned into Adobe Photoshop 4.0 to generate a composite figure.
Results
Comparatively, in situ localization of β-glucuronidase activity in spleen was less ambiguous in the pre-stained plastic sections than in the morphologically compromised frozen sections (Figures 1A and 1B). High levels of expression in both reticular cells and vessel-associated macrophages of the red pulp (Figure 1C were clearly demonstrable at highpower magnification. In the liver, x-gluc staining was variable throughout the parenchyma (Figure 1D, suggesting that different levels of enzyme activity exist among hepatocytes. However, when illuminated by darkfield (Figure 1E, a more uniform pattern of staining became apparent, as evidenced by the magenta-colored reaction product. Thus, darkfield analysis makes detection of enzyme activity possible in those hepatocytes that appear negative in the bright-field photomicrograph. In the CNS, where β-glucuronidase expression is much lower than in tissues such as spleen and liver, enzymatic activity is difficult to demonstrate by conventional, naphthol-AS-BI-β-d-glucuronic acid-based protocols. In cells such as the hippocampal neurons, for example, staining is virtually undetectable by this method. In contrast, our method of plastic embedding pre-stained tissue and visualization of reaction product by darkfield microscopy makes low-level detection possible in such cell populations (Figure 1F.
Discussion
Enzyme histochemistry is an invaluable tool for demonstration of endogenous enzyme activity in virtually any cell type, as well as exogenous reporter genes expressed by ex vivo-manipulated somatic cells. Histochemical demonstration of enzyme activity typically requires tissues to be processed with mild fixatives and cryosectioned to prevent enzyme denaturation. The unfortunate consequence, however, is often poor preservation of cytological architecture. Here we have developed a method for detecting β-glucuronidase activity that provides high-resolution localization of this enzyme in situ by employing the substrate 5-bromo-3-chloro-indolyl-β-D-glucuronic acid and glycol methacrylate tissue processing. Hydrolysis of this substrate generates a halogenated indoxyl derivative which is subsequently oxidized to an insoluble blue precipitate. Because the reaction product is not solubilized by glycol methacrylate, it is possible to stain tissue with this substrate first, then process the tissue in this embedding medium. An additional benefit of this method lies in the refractility of the blue precipitate observed when sections are illuminated under darkfield. By this method, appreciable light scatter is generated wherever reaction product is deposited, resulting in a very intense magenta color, depending on the local concentration of x-gluc staining. Therefore, even cells possessing low-level enzyme activity, which may be otherwise undetectable by brightfield, are demonstrable by this method. We and others have previously reported a similar technique for detection of β-galactosidase in transgenic mice (Sanes et al. 1986; Goring et al. 1987; Allen et al. 1988; Gossler et al. 1989; Lazik et al. 1997; Li et al. 1998). However, the method we describe here is not limited to the detection of β-glucuronidase or β-galactosidase but can also be employed to detect other enzymes for which halogenated indolyl conjugates are commercially available.
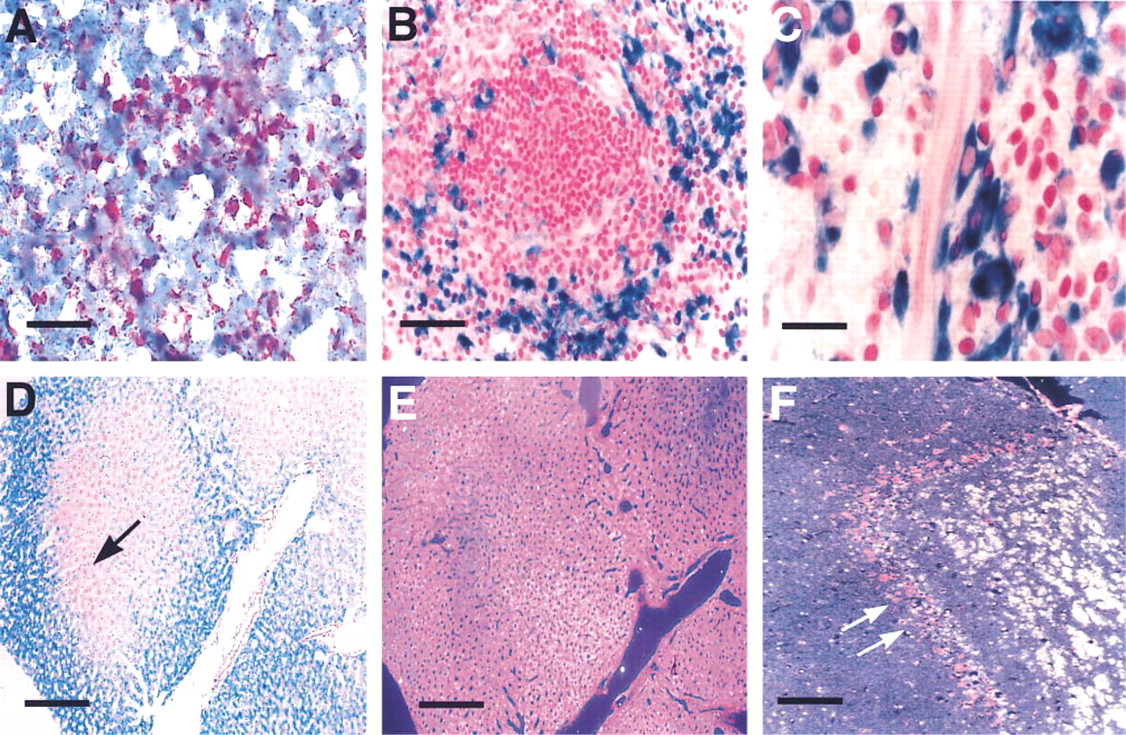
Murine tissues stained by enzyme histochemistry for β-glucuronidase. Brightfield of spleen cryosectioned at 10 μm and stained with naphthol-AS-BI-β-D-glucuronic acid (
β -Glucuronidase is expressed by virtually every cell in the vertebrate body and is the affected allele in the fatal human lysosomal storage disease mucopolysac-charidosis Type VII. Since the discovery of a mutation in the murine β-glucuronidase gene complex that gives rise to a phenotype similar to the human MPS VII, the gusmps mouse has been used as a model for treatment of this disease. Therefore, demonstration of endogenous levels of this enzyme in diseased mice of different zygosity, as well as enzyme replacement by transplanted donor cell populations, should be greatly improved by this histochemical technique.
Footnotes
Acknowledgements
Supported in part by NIH grant AR44507.
We would like to thank Robert Munger, HT (Allegheny University of the Health Sciences, Dept. of Pathology) for excellent technical expertise and helpful suggestions.