
Abstract
Metabolic mapping of enzyme activities (enzyme histochemistry) is an important tool to understand (patho)physiological functions of enzymes. A new enzyme histochemical method has been developed to detect transketolase activity in situ in various rat tissues and its ultrastructural localization in individual cells. In situ detection of transketolase is important because this multifunctional enzyme has been related with diseases such as cancer, diabetes, Alzheimer's disease, and Wernicke-Korsakoff's syndrome. The proposed method is based on the tetrazolium salt method applied to unfixed cryostat sections in the presence of polyvinyl alcohol. The method appeared to be specific for transketolase activity when the proper control reaction is performed and showed a linear increase of the amount of final reaction product with incubation time. Transketolase activity was studied in liver, small intestine, trachea, tongue, kidney, adrenal gland, and eye. Activity was found in liver parenchyma, epithelium of small intestine, trachea, tongue, proximal tubules of kidney and cornea, and ganglion cells in medulla of adrenal gland. To demonstrate transketolase activity ultrastructurally in liver parenchymal cells, the cupper iron method was used. It was shown that transketolase activity was present in peroxisomes and at membranes of granular endoplasmic reticulum. This ultrastructural localization is similar to that of glucose-6-phosphate dehydrogenase activity, suggesting activity of the pentose phosphate pathway at these sites. It is concluded that the method developed for in situ localization of transketolase activity for light and electron microscopy is specific and allows further investigation of the role of transketolase in (proliferation of) cancer cells and other pathophysiological processes.
M
Changes in TK activity have been demonstrated in a large number of pathologies. In Wernicke-Korsakoff's syndrome, diminished TK activity with reduced affinity for TPP has been shown to correlate with the neurological disorders and thiamine deficiency, which is characteristic for this disease (McCool et al. 1993). In Alzheimer's disease patients, TK activity has been found to be decreased because of proteolytic cleavage (Paoletti et al. 1997). Recently, it has been pointed out that TK plays an important role in preventing hyperglycemia-induced vascular damage (Hammes et al. 2003). This disease is related to diabetes and has its origin in inhibition of glyceraldehyde-3-phosphate dehydrogenase (GAPD) by superoxide metabolites. These reactive oxygen species are produced in the mitochondrial electron transport chain, and inhibition of GAPD results in increased levels of glyceraldehyde-3-phosphate and fructose-6-phosphate that activate the major pathways of hyperglycemic damage. It has been demonstrated that benfotiamine prevents the hyperglycemic damage in diabetic rats by activating TK, which converts these metabolites into pentose-5-phosphates and erythrose-4-phosphates (Hammes et al. 2003).
TK has also been associated with cell proliferation and cancer by its role in the generation of ribose-5-phosphate, which is the backbone of nucleic acids. An important role of TK in cancer cell proliferation was based on the development of thiamine deficiency in patients with breast and bronchial carcinomas (Basu and Dickerson, 1976). It has been established, using 13C-labeled glucose and stable isotope-based metabolic profiling, that cancer cells generate ribose-5-phosphate mainly by nonoxidative PPP reactions (Boros et al. 1997). Moreover, inhibition of TK activity, the rate-limiting enzyme of the nonoxidative PPP, resulted in a higher decrease of cancer cell growth than that obtained after inhibition of glucose-6-phosphate dehydrogenase (G6PD), the rate-limiting enzyme of the oxidative PPP (Raïs et al. 1999). These results stress the value of TK as therapeutic target to reduce cancer development.
Pathological alterations in TK activity may have many causes: decreased amounts of protein, posttranslational inactivation, or changed subcellular compartmentation of the enzyme. Traditional biochemical assays do not provide precise information on the localization of an enzyme, whereas immunohistochemistry does not allow conclusions whether the enzyme is active or inactive. Therefore, it is important to detect the activity of enzymes in their natural environment in cells or tissues by enzyme histochemical methods. Enzyme localization can be of great importance to understanding its function and to establishing whether disease alters activity and its ultrastructural localization, because the microenvironment of enzymes may determine their activity.
In the present study, a new enzyme histochemical method has been developed to detect TK activity in situ in various tissues and cells and to determine its ultrastructural localization. It is demonstrated that TK activity is present in liver parenchymal cells, small intestine, trachea, tongue, cornea and proximal tubules in kidney, and nerve cells. Moreover, it is shown that TK is not only a cytosolic enzyme as assumed so far, but is also present in peroxisomes and at membranes of granular endoplasmic reticulum in liver parenchymal cells. The ultrastructural localization of TK activity has large similarities with that of G6PD activity, suggesting that the entire PPP is active in these subcellular compartments.
In situ localization of TK activity can be useful to study its role in cancer progression and in diseases such as diabetes, Alzheimer's disease, and Wernicke-Korsakoff's syndrome.
Materials and Methods
Tissue Collection
Six male Wistar rats, weighing 200–250 g, were used to obtain liver, small intestine, trachea, tongue, kidney, adrenal gland, and eye tissues. The animals were sacrificed with an overdose of barbiturates. Treatment of the animals was in accordance with the Animal Ethical Committee guidelines at the Academic Medical Center of Amsterdam. The various organs were immediately removed and small parts of ∼0.5 cm3 were cut and frozen in liquid nitrogen and stored at −80C before use. Cryostat sections, 8 μm thick, were cut at −25C on a motor-driven cryostat (Bright; Huntington, UK) fitted with a rotary retracting microtome. Sections were picked up on clean glass slides and stored at −25C until use.
Isolation of Rat Liver Parenchymal Cells
Hepatocytes were isolated from 24 hr–starved male Wistar rats by collagenase perfusion, as described by Groen et al. (1982) and were kindly supplied by Dr. A. J. Meijer (Department of Biochemistry, Academic Medical Center, Amsterdam, The Netherlands). The obtained hepatocytes (∼107) were centrifuged in 1 ml Krebs-Henseleit bicarbonate buffer, pH 7.4, at 250 X g for 5 min at 4C. After centrifugation, cells were permeabilized for 30 min at room temperature under continuous rotation by adding 5 ml of a solution containing 0.025% (v/v) glutaraldehyde (Merck; Darmstadt, Germany) in 100 mM phosphate buffer, pH 7.4 (Frederiks and Vreeling-Sindelárová 2001). Then, cells were washed three times for 1 min with 3 ml 100 mM phosphate buffer, pH 7.4, at 4C, followed by rapid centrifugation at 250 X g. The cells were resuspended in 0.5 ml of the same buffer. Fresh cells and permeabilized cells were brought onto glass slides to demonstrate the effect of glutaraldehyde treatment on TK activity in individual cells. Permeabilized cells were also used to demonstrate the ultrastructural localization of TK activity.
Light Microscopical Procedure for the Localization of TK Activity
TK activity was localized for light microscopical purposes by a modification of the dehydrogenase method with a tetrazolium salt and polyvinyl alcohol as described by Van Noorden and Frederiks (1992). The substrates for the histochemical reaction were the same as for the biochemical method to determine TK activity in homogenates (Smeets et al. 1971). The reaction was based on the transfer of the ketol moiety from xylulose-5-phosphate to ribose-5-phosphate, obtaining sedoheptulose-7-phosphate and glyceraldehyde-3-phosphate. Monitoring TK activity can be achieved by coupling the reaction to GAPD with the formation of NADH, which is the first product in the chain reaction of electron transfers that will lead to formazan formation precipitated at the exact place where the reaction has taken place. Therefore, GAPD is an auxiliary in the present assay to detect TK activity, as described previously for other enzymes that need an auxiliary enzyme such as hexokinase, creatine kinase, and phosphofructokinase (Frederiks et al. 1987). Incubation media were prepared using 50 mM Tris-HCl buffer, pH 7.6, containing 18% (w/v) polyvinyl alcohol (weight average
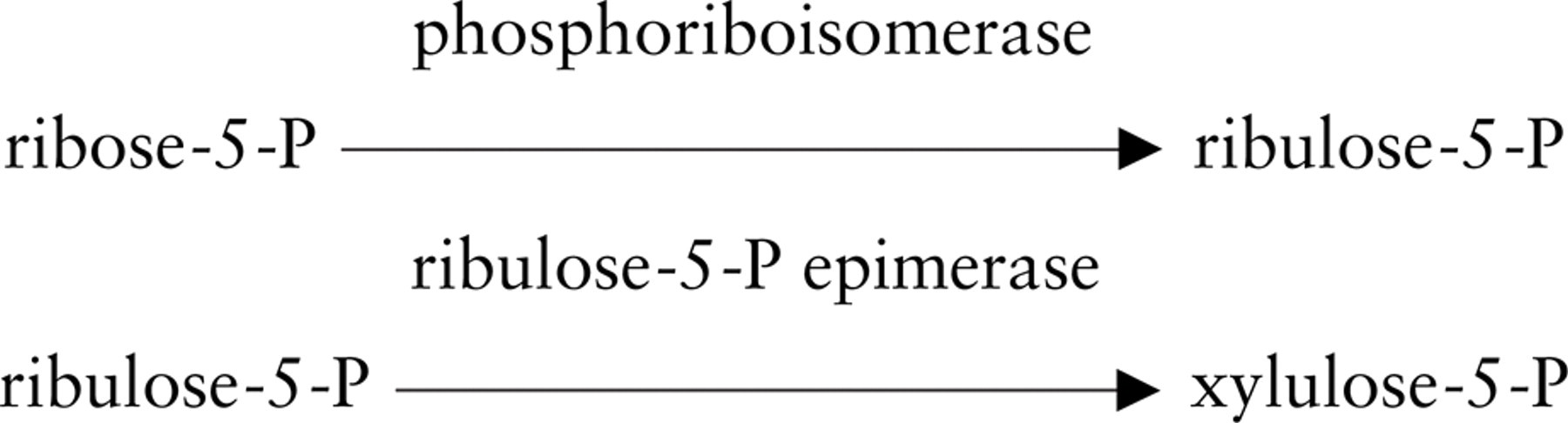
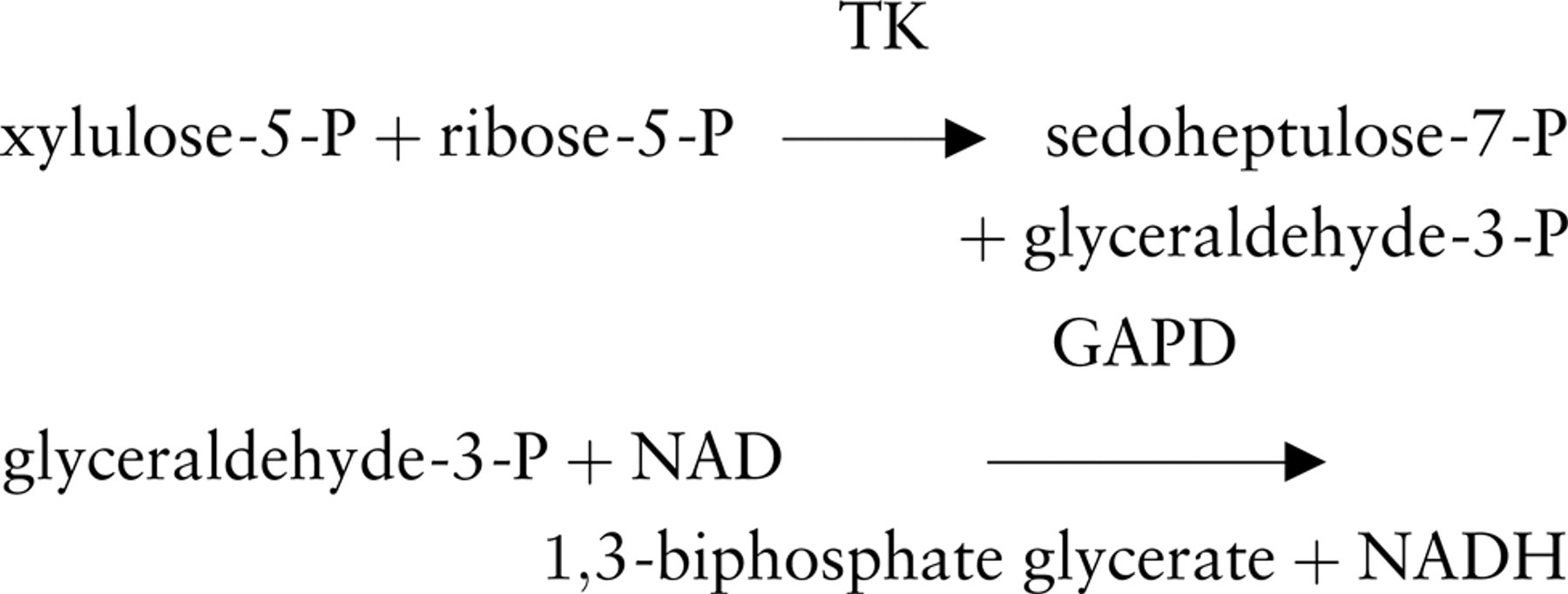
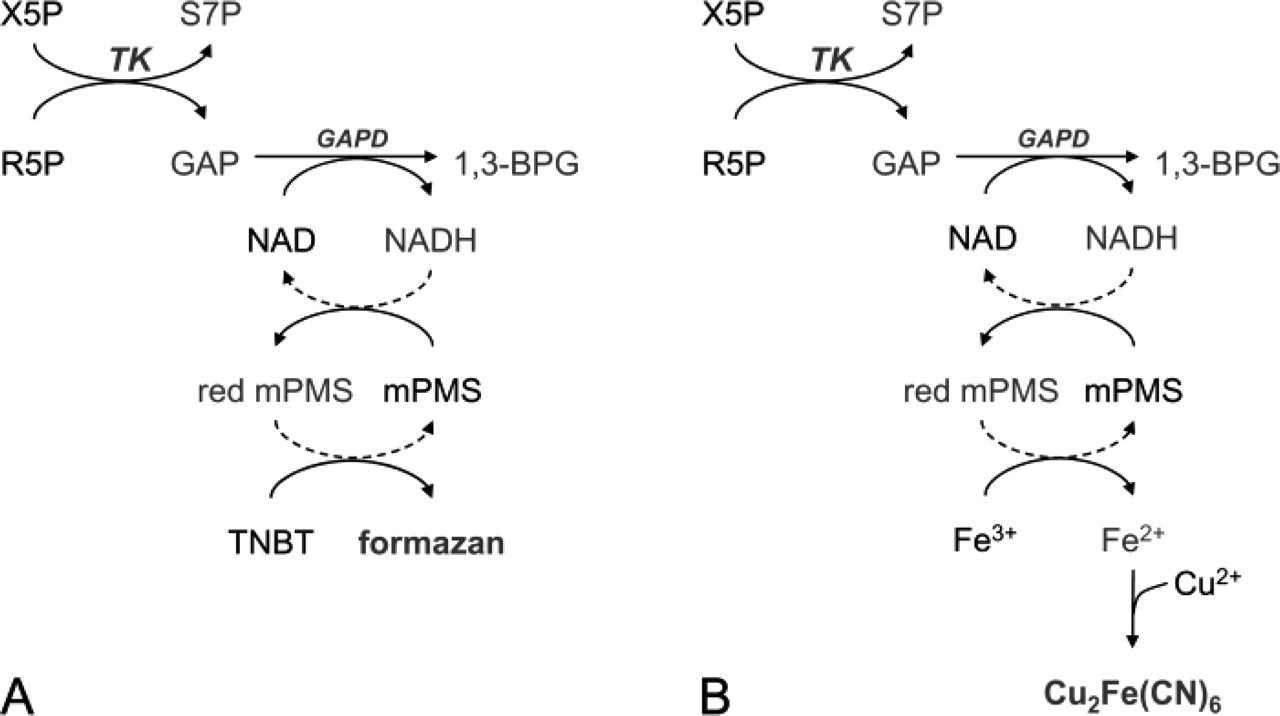
Schemes showing the reactions involved in the histochemical assay to detect transketolase activity in unfixed sections or cells for light microscopy (
The reactions involved in the histochemical assay are shown in Figure 1A. Control reactions were performed by using incubation media that lacked the substrate mixture. The specificity of the reaction was established by adding 10 mM ADP to the test and to the control incubation media (Hosomi et al. 1989). The importance of the auxiliary enzyme was tested by incubations with and without exogenous GAPD (Sigma) according to Van Noorden and Frederiks (1992). The effect of addition of 0.1 mM TPP to incubation media was also investigated. Incubations were performed at 37C. To stop the reaction, sections were rinsed with phosphate buffer, pH 5.0, at 65C and then mounted in gelatin-glycerin. The validity of the histochemical method was established by measuring the amount of formazan in relation to incubation time. The specific reaction was defined as the test minus control reaction.
GAPD activity was demonstrated using the incubation medium to demonstrate TK activity, in which the substrate mixture was replaced by 10 mM fructose-1,6 diphosphate (Henderson, 1976).
Quantification of TK Activity Using Image Analysis
To quantify TK activity in liver tissue, amounts of final reaction product were determined by image analysis using a Vanox-T photomicroscope (objective 2X) (Olympus; Tokyo, Japan) with a monochromatic filter of the isobestic wavelength of TNBT-formazan (557 nm). For image capturing, a Cohu 4913 CCD camera (Cohu; San Diego, CA) was used, and data analysis was performed using the public domain NIH Imaging software program available via the Internet from http://rsb.info.nih.gov/. To obtain specific TK activity, absorbances in images of control incubations were subtracted from absorbances in images of test reactions. Moreover, absorbances in images of incubations performed in the presence of 10 mM ADP were determined. Quantification was performed according to Jonker et al. (1995). In each section, four areas were measured and values were averaged.
Electron Microscopical Procedure for the Localization of TK Activity
The cupper iron method to detect TK activity was a modification of the method published by Frederiks and Vreeling-Sindelárová (2001) for the ultrastructural localization of G6PD activity in cells. Liver parenchymal cells were permeabilized with glutaraldehyde as described previously. The incubation medium was prepared in 50 mM Tris-HCl buffer, pH 7.6, containing 6% polyvinyl alcohol. Test incubation media contained 5 mM sodium azide, 5 mM MgCl2, 3.7 mM KH2PO4, 10 mM potassium ferricyanide, 30 mM sodium citrate, 30 mM cupric sulfate, 0.45 mM methoxyphenazine methosulphate, 7.5 mM NAD, and 100 μl substrate mixture per ml incubation medium (prepared as described previously). The reactions involved in the histochemical assay for electron microscopy are shown in Figure 1B. Control reactions were performed using media lacking the substrate mixture and in the presence of 10 mM ADP. Incubations were performed for 1 hr at 37C under continuous agitation. The incubation was stopped by addition of 5 ml cold phosphate buffer. Several washes with phosphate buffer were performed to remove incubation media.
After incubation, cells were fixed immediately with 4% paraformaldehyde and 1% (v/v) glutaraldehyde in 100 mM cacodylate buffer, pH 7.4, at 4C for 2–48 hr. After fixation, cells were rinsed with 100 mM cacodylate buffer, pH 7.4, for 40 min, postfixed with 1% OsO4 in 100 mM cacodylate buffer, pH 7.4, for 60 min at 4C or with 1% OsO4 and 1.5% potassium ferrocyanide in 100 mM phosphate buffer, pH 7.4, for 2 hr at 4C and thoroughly rinsed with bidistilled water at 4C. Afterwards, samples were dehydrated and embedded in epoxy resin LX-112 according to standard procedures. Semithin sections (1–2 μm thick) were cut on a LKB Pyramitone and stained with methylene blue to check the quality of the cell preparations. Ultrathin sections (30–70 nm thick) were cut on an ULTRACUT E ultramicrotome (Leica Microsystems; Wezlar, Germany) and studied with an EM-10C transmission electron microscope (Zeiss; Oberkochen, Germany).
The activity of GAPD was demonstrated using the incubation medium to demonstrate TK activity in which the substrate mixture was replaced by 10 mM fructose-1,6 diphosphate.
Results
Light Microscopical Method to Detect TK Activity In Situ
Unfixed cryostat sections of rat liver incubated for the detection of TK activity showed formazan formation in liver parenchymal cells after incubation in the presence of the substrate mixture (Figure 2). Addition of exogenous GAPD did not affect the amounts of formazan produced. This means that endogenous GAPD activity is high enough for driving formazan formation by TK activity. Addition of TPP to incubation media did not affect the amounts of final reaction product generated in liver parenchymal cells incubated with test and control incubation media, suggesting that the endogenous TPP concentration was not limiting for the detection of TK activity. The amounts of formazan produced in liver parenchyma increased linearly with incubation time (Figure 2). After incubation in the absence of the substrate mixture, high amounts of formazan were produced that also increased linearly with time (Figure 2). To test the specificity of the test minus control reaction, test and control incubations were performed in the presence of 10 mM ADP, an inhibitor of TK activity. Test and control reactions were strongly inhibited by ADP and both reactions did not increase in time (Figure 2). This means that the relatively high control reaction was due to the use of endogenous substrates by TK to produce final reaction product. Therefore, we conclude that the specific TK reaction is the test reaction in the presence of the substrate mixture of TK minus the proper control reaction, which is an incubation in the absence of the substrate mixture of TK and in the presence of 10 mM ADP. This test reaction minus the proper control reaction is again linear in time (Figure 2). ADP did not affect GAPD activity (data not shown), which means that the effect of ADP on the TK reaction is a consequence of inhibition of TK activity and not from inhibition of the auxiliary enzyme.
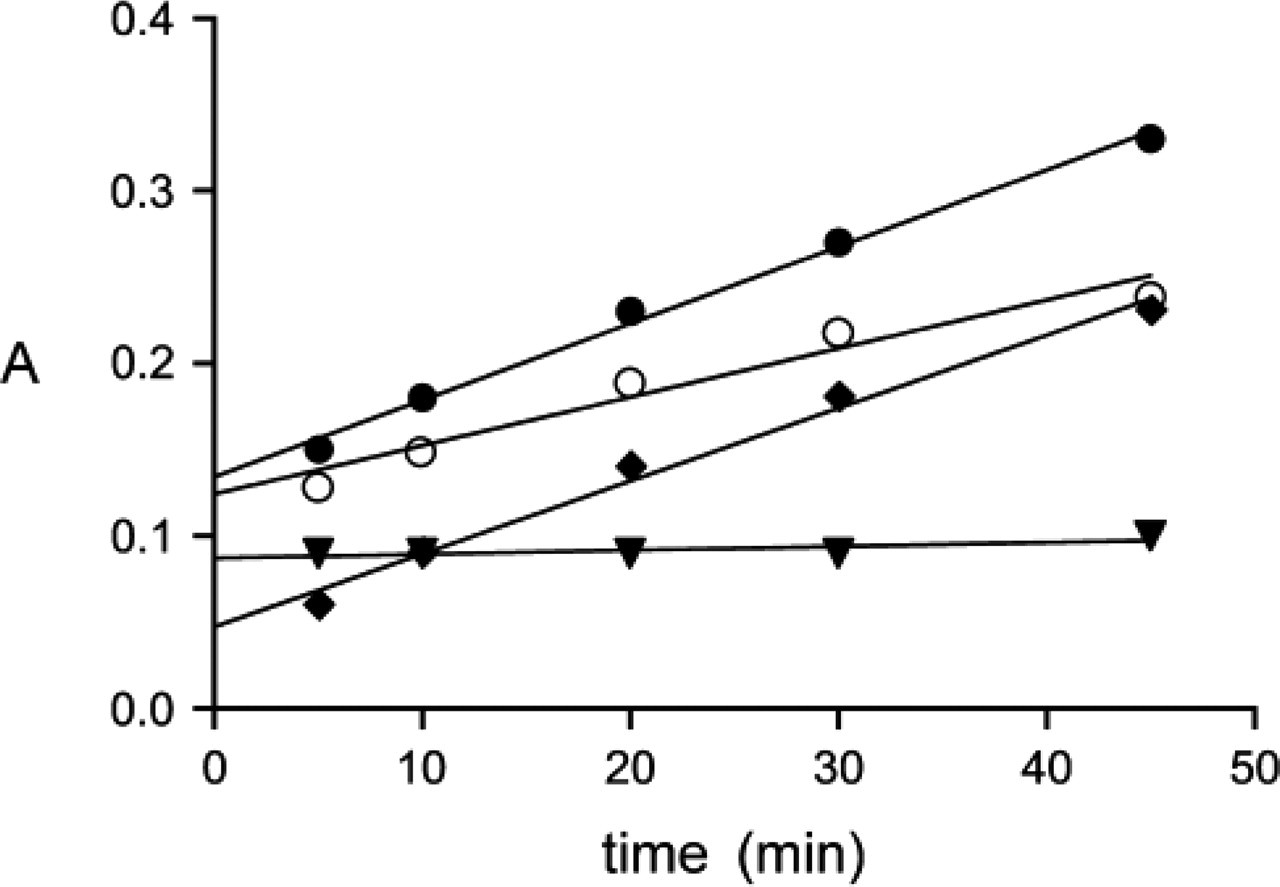
The relation between mean absorbance (A) and incubation time as measured in pericentral areas of rat liver incubated for the demonstration of transketolase activity in the presence of the substrate mixture (closed circles, test reaction), in the absence of the substrate mixture (open circles, control reaction), and in the absence of the substrate mixture and in the presence of 10 mM ADP (triangles, real control reaction); the specific reaction (diamonds) is the test reaction (in the presence of the substrate mixture) minus the real control reaction (in the absence of the substrate mixture and in the presence of 10 mM ADP).
Localization of TK Activity in Various Tissues
The histochemical procedure to detect TK activity was applied to various organs. In rat liver, TK activity was mainly localized in liver parenchymal cells and was higher in pericentral than in periportal areas (Figure 3). Enzyme activity in small intestine was present in epithelial cells (enterocytes and goblet cells). Activity was higher in epithelium in villi than in crypts (Figure 3). In trachea, activity was found in the epithelial cell layer of the wall (Figure 3). Activity was present in goblet cells, cilia-containing cells and basal cells. TK activity was present in the epithelial layer of the tongue with highest activity in the basal cells, whereas TK activity was absent in the superficial cornified layer (Figure 3D). In kidney, TK activity was observed in epithelial cells of proximal tubules but not in epithelial cells of distal tubules, collecting ducts, lis of Henle, and glomerular cells (Figure 3E). In adrenal gland, TK activity was found in particular cells in the medulla but not in the cortex (Figure 3F). In rat eye, TK activity was present in epithelial cells of the cornea (Figure 3G) and in nerve cells of the retina (Figure 3H). Control reactions performed in the absence of the substrate mixture and in the presence of ADP were similar to reactions performed in the presence of the substrate mixture and ADP, again indicating the specificity of the reaction. Hardly any formazan was produced under these conditions in all tissues tested.
TK activity was also demonstrated in isolated liver parenchymal cells. The permeabilization procedure with 0.025% glutaraldehyde, which is necessary for TK incubations for electron microscopy, did not affect the amount of formazan produced by liver parenchymal cells (Figure 4). This was confirmed by measurements with image analysis. The morphology of glutaraldehyde-treated cells is better than of untreated cells brought on glass slides. Moreover, untreated cells showed inhomogeneity with respect to TK activity (Figure 4).
In all tissues, activity of GAPD was higher than that of TK (data not shown).
Ultrastructural Localization of TK Activity
The histochemical procedure to localize TK activity in subcellular compartments was performed on permeabilized liver parenchymal cells using the cupper iron method. Electron-dense reaction product from TK activity was present in peroxisomes and at membranes of the granular endoplasmic reticulum (Figure 5). Reaction product observed in nuclei and at the plasma membrane was nonspecific, because cells incubated in the presence of control media showed reaction product at these sites as well. This is in agreement with previous findings obtained in control incubations for G6PD activity (Frederiks and Vreeling-Sindelárová 2001). Ultrastructural morphology of the cells is rather well-preserved after incubation. GAPD activity was found to be localized similarly as TK activity in liver parenchymal cells suggesting that the auxiliary enzyme was not rate limiting at any site for the demonstration of TK activity (data not shown).
Discussion
In the present study, an enzyme histochemical method to detect TK activity in situ is described. TK is a moonlighting protein, showing different functions depending on its localization, and has been implicated in different pathologies such as cancer, diabetes, and neurological disorders (McCool et al. 1993; Sax et al. 1996; Paoletti et al. 1997; Hammes et al. 2003). The method that we developed was shown to be specific for TK activity when the proper control reaction—an incubation in the absence of the substrate mixture and in the presence of 10 mM ADP—is subtracted from the test reaction, which is an incubation in the presence of the substrate mixture.
The procedure to demonstrate TK activity in unfixed cryostat sections or cells is as follows:
1. Dry, unfixed cryostat sections or unfixed cells brought on glass slides for 5 min at room temperature.
2. Incubate sections or cells for 15–120 min at 37C in test incubation media in the presence of the substrate mixture and in the proper control media; this is in the absence of the substrate mixture and in the presence of 10 mM ADP.
3. The incubation medium consists of
18% (w/v) polyvinyl alcohol in 50 mM Tris-HCl buffer, pH 7.6
5 mM sodium azide
7.5 mM NAD
3.7 mM KH2PO4
5 mM MgCl2
0.45 mM methoxyphenazine methosulphate
0.1 mM thiamine pyrophosphate
5.5 mM tetra nitroblue tetrazolium salt
100 μl substrate mixture per ml
The substrate mixture was prepared by dissolving 50 mM ribose-5-phosphate in 50 mM Tris-HCl buffer, pH 7.6, and adding 0.05 IU ribulose-5-phosphate epimerase and 0.25 IU phosphoriboisomerase. The substrate mixture was continuously stirred and heated at 37C for 1 hr and then kept at −20C until use.
4. Incubate sections or cells for the demonstration of GAPD activity; this is the incubation medium described previously, in which the substrate mixture is replaced by 10 mM fructose-1,6-diphosphate. The activity of the auxiliary enzyme GAPD should be higher than TK activity.
5. Rinse sections after the incubations with 100 mM phosphate buffer, pH 5.0, at 65C.
6. Mount in glycerin-gelatin.
7. Observe the localization and the amount of formazan with a light microscope.
The procedure was applied to sections of various organs of the rat. It was found that the level of enzyme activity was dependent on the organ and also on the region within the organ where it was found. This indicates the importance of this new method to localize TK activity in situ as classical biochemical methods use homogenates of organs where the different components are mixed, obtaining average activities only. We have found that TK activity was higher in pericentral than in periportal areas of rat liver. This is different from the localization of G6PD activity, the rate-limiting enzyme of the oxidative PPP that is extremely high in Kupffer cells and shows an equal distribution in liver lobules of male rats (Jonges et al. 1995; Jonges and Van Noorden 1989). However, the localization of TK activity is largely in agreement with that of phosphogluconate dehydrogenase activity, the second enzyme of the oxidative branch of the PPP (Jonges et al. 1995). It is suggested that the main function of TK in rat liver is recycling of carbons produced by G6PD back to glycolysis to provide NADPH for detoxification and lipid synthesis. On the other hand, TK activity was found to be high in epithelial cells of proximal tubules of rat kidney, which reabsorbs many compounds from primary urine, such as amino acids, small proteins, and glucose. Therefore, TK may be related with anabolic and catabolic processes in these epithelial cells using the absorbed compounds. High TK activity was also found in epithelial cells of other organs such as trachea, tongue, and small intestine in combination with G6PD indicating NADPH production for protection against reactive oxygen species (Biagiotti et al. 2000). On the other hand, the presence of TK activity especially in basal cells of squamous epithelium of the tongue is in contrast with the low activity of G6PD in these cells, suggesting a role of TK in cell proliferation (Biagiotti et al. 2000). In rat eye, highest TK activity was observed in cornea and in nerve cells of the retina. In cornea, TK probably has another function than in other epithelia because here it is a structural protein, being important for corneal transparency (Jester et al. 1999). These studies were based on TK protein and mRNA distribution and, therefore, it cannot be established which part of the protein is active (Sax et al. 1996). The high TK activity in nerve cells of rat eye and adrenal medulla is in agreement with the high G6PD activity in these cells, as demonstrated previously (Biagiotti et al. 2001). The cells with high TK activity and possibly G6PD activity are likely ganglion cells. Biagiotti et al. (2001) propose a number of functions for the high G6PD activity in these cells, which are related with the supply of NADPH for the NO synthase reaction and the production of neurosteroids (Biagiotti et al. 2003). The rather low TK activity in the cortex of adrenal gland is in contrast with the very high G6PD activity in this part (Berchtold 1979; Ishibashi et al. 1999), suggesting that NADPH is produced for biosynthesis of hormones rather than recycling of carbons.
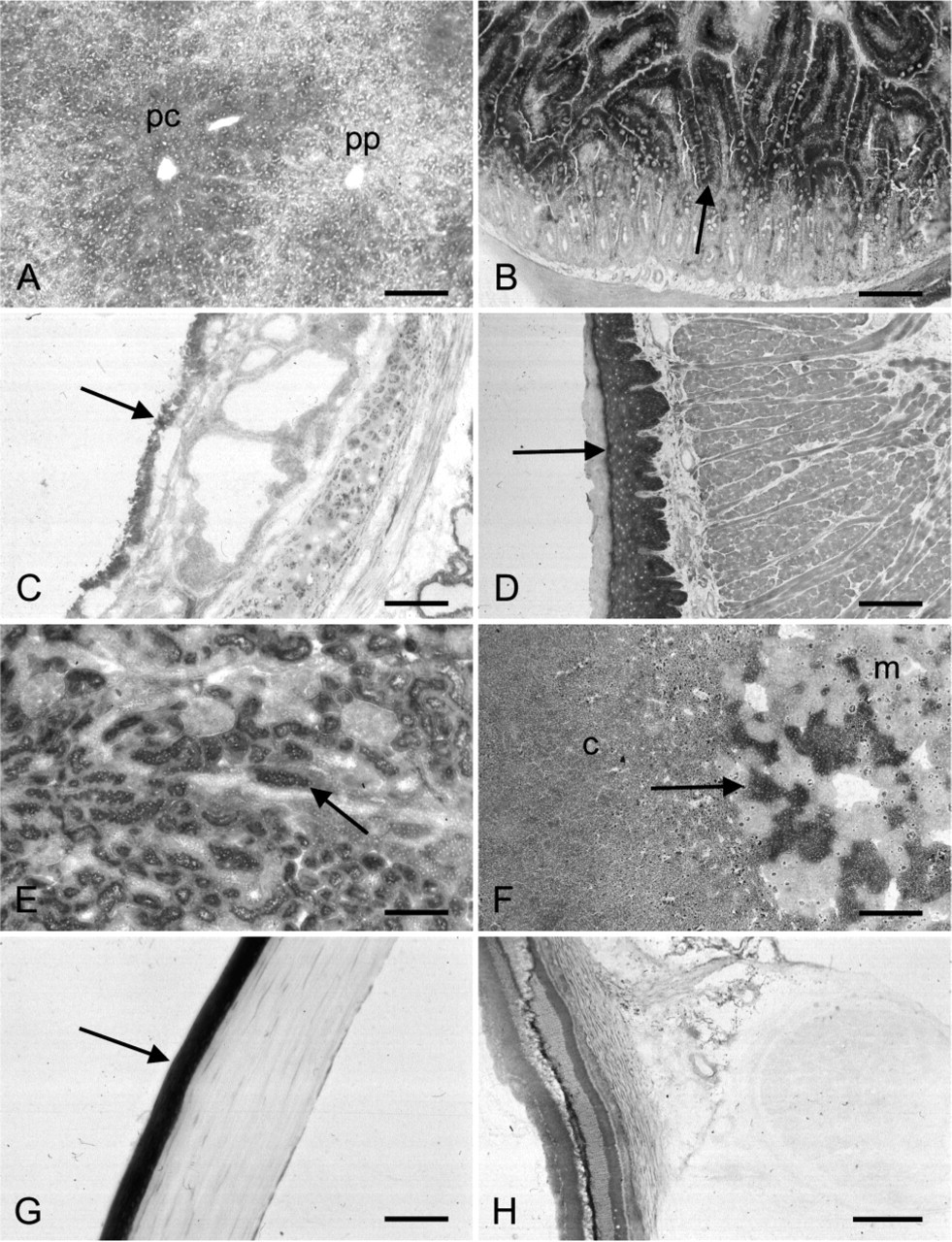
Localization of transketolase activity in various rat tissues. Enzyme activity is present in liver parenchymal cells with a higher activity in pericentral (pc) than in periportal areas (pp) (
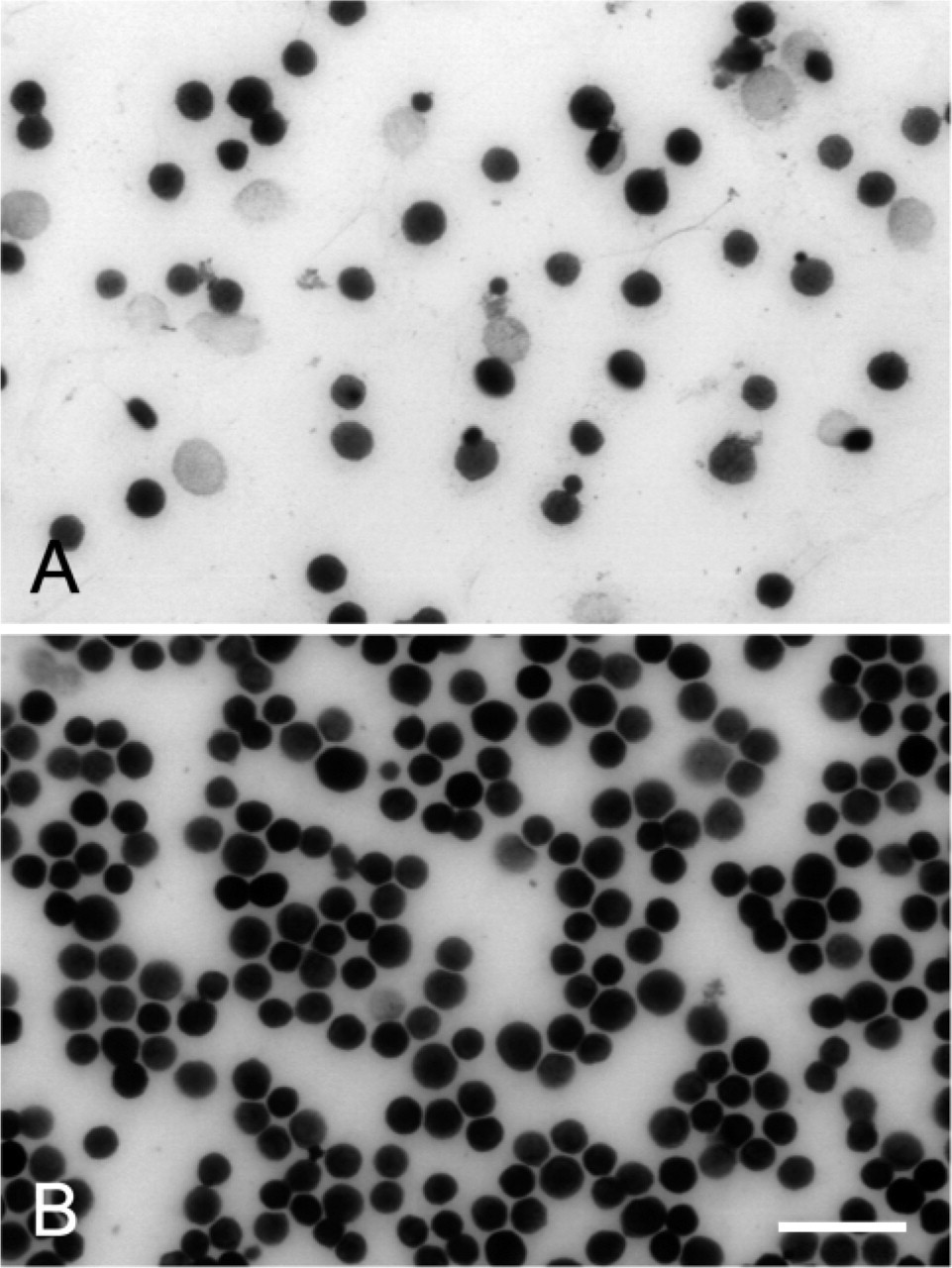
Effect of permeabilization with 0.025% glutaraldehyde on transketolase activity in isolated liver parenchymal cells. The amounts of final reaction product in fresh liver parenchymal cells (
Another important finding reported in the present study is the subcellular localization of TK activity. To demonstrate enzyme activity in cells at the ultrastructural level, cells should be permeabilized. In the present study, we applied 0.025% glutaraldehyde, which has been proven to be an excellent way for cell permeabilization (Van Noorden et al. 1982; Van Noorden and Vogels 1985). Moreover, this treatment did not affect TK activity (Figure 4), whereas it was shown before that G6PD activity was reduced by ∼30% (Frederiks and Vreeling-Sindelárová 2001). Previously, it has been demonstrated that some enzymes that were considered to be cytoplasmic were also present in other cellular compartments. For example, G6PD has been found at membranes of granular endoplasmic reticulum, in peroxisomes, and in the cytoplasm of rat liver parenchymal cells (Frederiks and Vreeling-Sindelárová 2001). In line with these findings, Veitch et al. (2004) have demonstrated recently that TK was associated with glycosomes, peroxisome-like organelles, in
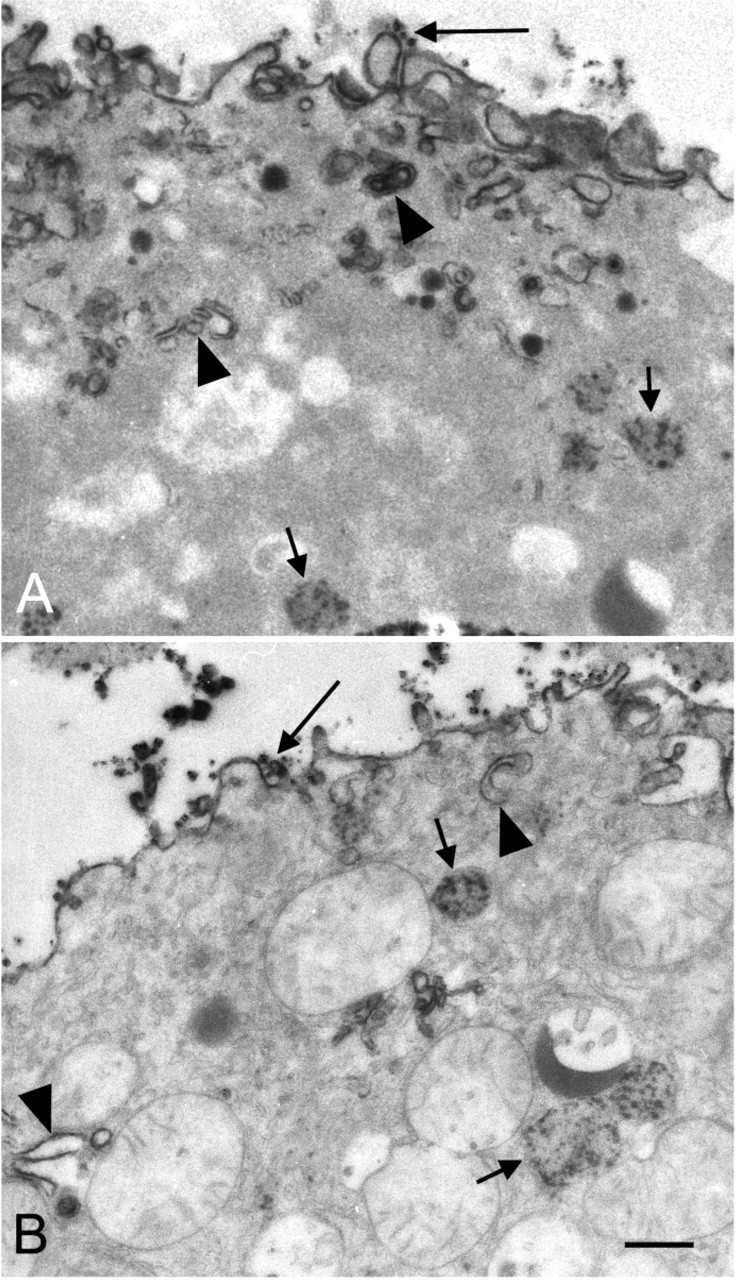
Electron micrographs of isolated liver parenchymal cells incubated for the demonstration of transketolase activity. Specific electron dense reaction product is present in peroxisomes (small arrows) and at membranes of granular endoplasmic reticulum (arrowheads). Aspecific reaction product (large arrows) is observed at the plasma membrane. Morphology of the cells is well-preserved as demonstrated in
In conclusion, the method developed for in situ localization of TK activity for light and electron microscopy is specific and allows further investigation of the role of TK in (proliferation of) cancer cells and other pathophysiological processes.
Footnotes
Acknowledgements
Support is given by “La Caixa” Oncology Grant Program (Fundacio “La Caixa,” ON03/070–00).
The authors are grateful to Prof. Dr. C.J.F. van Noorden for fruitful discussions and critical review of the manuscript, Mr. J.P.M. Peeterse for useful help with the illustrations, and Mrs. T.M.S. Pierik for careful preparation of the manuscript.