
Abstract
Mitochondrial carbonic anhydrase V (CA V) in liver provides HCO3 − to pyruvate carboxylase for the first step in gluconeogenesis and HCO3 − to carbamyl phosphate synthetase I for the first step in ureagenesis. Because carbamyl phosphate synthetase I and ornithine transcarbamylase are also expressed in enterocytes, we tested the hypothesis that CA V is expressed in the gastrointestinal tract in addition to liver. Polyclonal rabbit antisera were raised against a polypeptide of 17 C-terminal amino acids of human CA V and against purified recombinant mouse isozyme and were used in Western blotting and immunoperoxidase staining of human and rat tissues. Immunohistochemistry showed that CA V is expressed cell-specifically in the alimentary canal mucosa from stomach to rectum. Immunoreactions for CA V were detected in the parietal cells and gastrin-producing G-cells of the stomach and in intestinal enterocytes. Western blotting of human and rat gastrointestinal tissues with isozyme-specific antibodies showed positive signals for CA V with the expected molecular mass. The findings in human tissues paralleled those in rat. The cell-specific pattern of CA V expression suggests a role for CA V in alimentary canal physiology. We propose that mitochondrial CA V participates in the detoxification of ammonia produced in the gastrointestinal tract by providing bicarbonate to carbamyl phosphate synthetase I.
Keywords
C
Because carbamyl phosphate synthetase I and ornithine transcarbamylase have both been reported to be expressed in epithelial cells of the alimentary tract, we tested the hypothesis that CA V is also expressed there. Here we demonstrate that CA V is indeed expressed in specific cells in human and rat gastrointestinal canal. The cell-specific expression of CA V in the epithelia and endocrine cells of the alimentary tract suggests a functional role in ureagenesis and/or gluconeogenesis in extrahepatic organs.
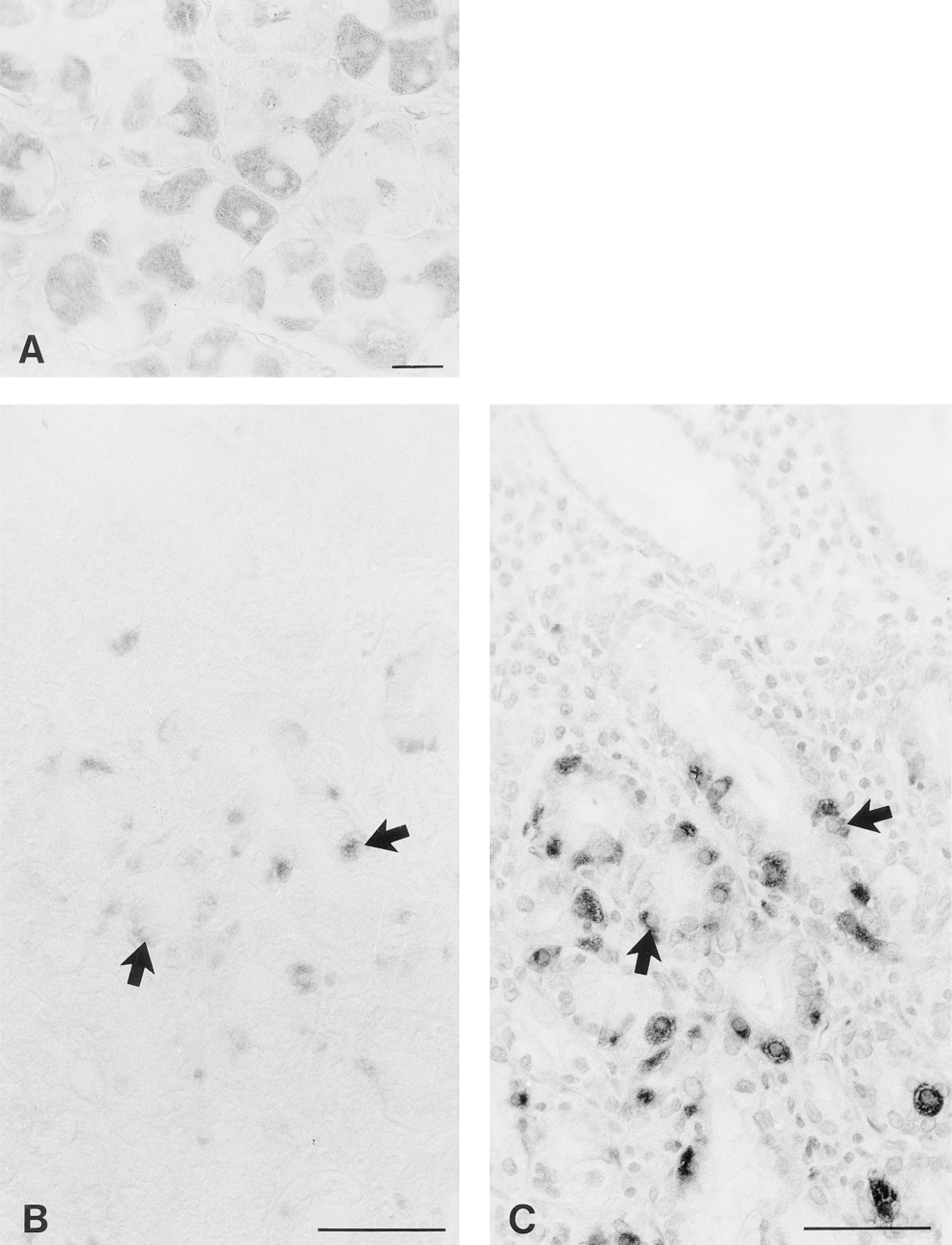
Immunohistochemical demonstration of CA V in the human gastrointestinal tract. In the body of the stomach, the parietal cells show fine granular immunostaining (A). Bar = 20 μm. In the pyloric antrum, positive signal for CA V is seen in cells with the morphological appearance of endocrine cells (B). These cells were identified as gastrin-producing G-cells by subjecting the parallel section of the pyloric antrum to immunostaining with an anti-gastrin antibody (C). The arrowheads indicate the same cells in the parallel sections. In the ileum (D), cecum (E), and ascending colon (F), the immunoreaction for CA V is confined to the superficial enterocytes. Bars = 50 μm.
Materials and Methods
Antibodies
Polyclonal rabbit antiserum against the synthetic C-terminal 17-amino acid peptide of human CA V was produced and characterized by Nagao et al. (1993). The anti-CA V serum was preabsorbed with purified human CA I, CA II, and CA IV before use for immunohistochemistry to avoid crossreactions with these isozymes, even though no crossreaction could be demonstrated by Western blots with CA I, II, III, or IV. Purified CA I, CA II, and CA IV (300 μg each) were adsorbed onto a nitrocellulose filter using a dot-blot apparatus (Bio-Rad Laboratories; Richmond, CA). Pieces of filter containing adsorbed CA I, II, and IV were incubated with 30 μl of the antiserum diluted 1:100 in 1% bovine serum albumin (BSA) in 0.1 M PBS for 12 hr at 4C, and the preabsorbed antiserum was used for the experiments at 1:100 dilution.
Polyclonal anti-mouse CA V serum was raised in a rabbit using recombinant mouse CA V (Heck et al. 1994) (a generous gift from Dr. David Silverman) as described (Zhu and Sly 1990). Anti-CA V IgG was affinity-purified using recombinant mouse CA V coupled to Affi-Gel 10 matrix. The anti-mouse CA V IgG recognized mouse and rat CA V but did not recognize any other CA isozymes tested (CA I, II, III, IV). The anti-mouse CA V antibody reacted equally well with rat and mouse CA V on Western blot and on immunohistochemistry of COS cells expressing rat or mouse CA V cDNA (data not shown).
Rabbit anti-human gastrin antibody was purchased from Dakopatts (Glostrup, Denmark).
Preparation of Tissues
Human tissue samples were obtained together with routine histopathological specimens taken during surgical operations carried out on four patients at Oulu University Hospital. The rat tissue samples were obtained from four adult rats, which were sacrificed by decapitation. Each tissue sample was divided into several small pieces about 5 mm thick. The specimens were fixed in 4% paraformaldehyde in 0.1 M PBS for 18 hr. All fixations were carried out at 4C. The samples were then dehydrated, embedded in paraffin in a vacuum oven at 58C, and 5-μm sections were placed on gelatin-coated microscope slides. The animal and human studies were approved by the ethics committee of the Oulu University Hospital.
Immunohistochemistry
CA isozymes were immunostained using the biotin-streptavidin complex method. The steps in the stainings were as follows: (a) pre-treatment of the sections with undiluted cow colostral whey for 40 min and rinsing in PBS; (b) incubation for 12 hr with the anti-human CA V serum diluted 1:100 in 1% BSA-PBS or with affinity-purified anti-mouse CA V antibody (10 μg/microscope slide) diluted in 1% BSA-PBS; (c) treatment with cow colostral whey for 40 min and rinsing in PBS; (d) incubation for 1 hr with biotinylated swine anti-rabbit IgG (Dakopatts) diluted 1:300 in 1% BSA-PBS; (e) incubation for 30 min with peroxidase-conjugated streptavidin (Dakopatts) diluted 1:600 in PBS; and (f) incubation for 2 min in DAB solution containing 9 mg 3,3,'-diaminobenzi dine tetrahydrochloride (Fluka; Buchs, Switzerland) in 15 ml PBS plus 10 μl 30% H2O2. The sections were washed in PBS after incubation steps b, d, and e. All the incubations and washings were carried out at room temperature, and the sections were finally mounted in Permount (Fisher Scientific; Fair Lawn, NJ). The stained sections were examined and photographed with a Leitz Aristoplan (Wetzlar, Germany) microscope.
SDS-polyacrylamide Gel Electrophoresis (SDS-PAGE) and Western Blotting
All the reagents for SDS-PAGE were from Bio-Rad Laboratories or Sigma (St Louis, MO). The electrophoreses were performed under reducing conditions according to Laemmli (1970). Protein concentrations were determined by a Lowry (Peterson 1979) or Bio-Rad protein assay, and 5 μg (human tissue homogenates) and 150 μg samples (rat tissue homogenates) were applied to each lane. The proteins were transferred electrophoretically from the gel to a polyvinylidene fluoride (PVDF) membrane as described (Waheed et al. 1992). After the transblotting the PVDF membranes were subjected to immunostaining with anti-human or anti-mouse CA V antibodies. The polypeptide bands of human and rat CA V were visualized by chemiluminescence reaction (Kivelä et al. 1997) or 4-chloronaphthol (Waheed et al. 1992), respectively.
Results
Immunohistochemistry
The distribution of mitochondrial CA V was studied in tissue samples obtained from normal human and rat gastrointestinal tract. Both human and rat tissues expressed the enzyme in a similar tissue- and cell-specific manner. In the stomach, the parietal cells of the gastric glands showed quite strong positive immunoreaction (Figures 1A and 2A). The surface epithelial cells of the gastric pit region also exhibited positive immunostaining (not shown). In addition, the endocrine cells of the pyloric antrum expressed CA V (Figure 1B). The immunohistochemical staining of serial sections with anti-gastrin (Figure 1C) and anti-CA V (Figure 1B) antibodies suggested that the CA V-positive cells in the pyloric antrum represented gastrin-producing G-cells.
The non-goblet surface epithelial cells of the entire gut from the duodenum to the rectum expressed CA V (Figures 1D-1F and 2B-2G). However, the staining was more intense in the large intestine than in the small intestine, the latter showing only weak immunoreactivity in the villous enterocytes (Figures 1D and 2B-2D). In the large intestine, the superficial enterocytes of the surface epithelial cuff region showed quite strong immunoreaction (Figures 1E, 1F, and 2E-2G). The high-magnification view (Figure 2G) demonstrates that the signal for CA V was fine and granular, suggesting that a predominant fraction of the enzyme is localized in the mitochondria. The control immunostainings using nonimmune rabbit serum in place of the first antibody showed no positive immunoreaction (compare Figures 2H and 2G).
Western Blotting of CA V in the Human and Rat Gastrointestinal Tract
The presence of CA V in the alimentary tract tissues was confirmed by Western blotting of some selected tissues from human and rat alimentary tract. The anti-human CA V antibody identified two polypeptide bands of 34 and 30 kD in the tissue homogenates of the human ileum, ascending colon, and rectum, representing the precursor (34-kD) and mature (30-kD) forms of CA V (Figure 3). The strongest signal of these three was identified in the ascending colon.
All rat tissue homogenates showed precursor (34-kD), mature (30-kD), and proteolytically degraded (28-kD) forms of CA V polypeptides previously identified in rat liver, isolated rat liver mitochondria, and COS cells (Nagao et al. 1994). The signals for CA V were slightly stronger in the stomach, cecum, and colon than in the small intestine. The intensity of CA V signal in the duodenum and jejunum was higher than in the ileum. Cecum, proximal, middle, and distal segments of the colon (Figure 4) showed the presence of CA V with an apparent molecular weight of 28–34 kD, and no distinct differences in the intensity of signals were detected among these segments (not shown). Interestingly, the signals for the precursor (34-kD) and proteolytically degraded (28-kD) forms of CA V were much stronger in the stomach and large intestine than in the small intestine.
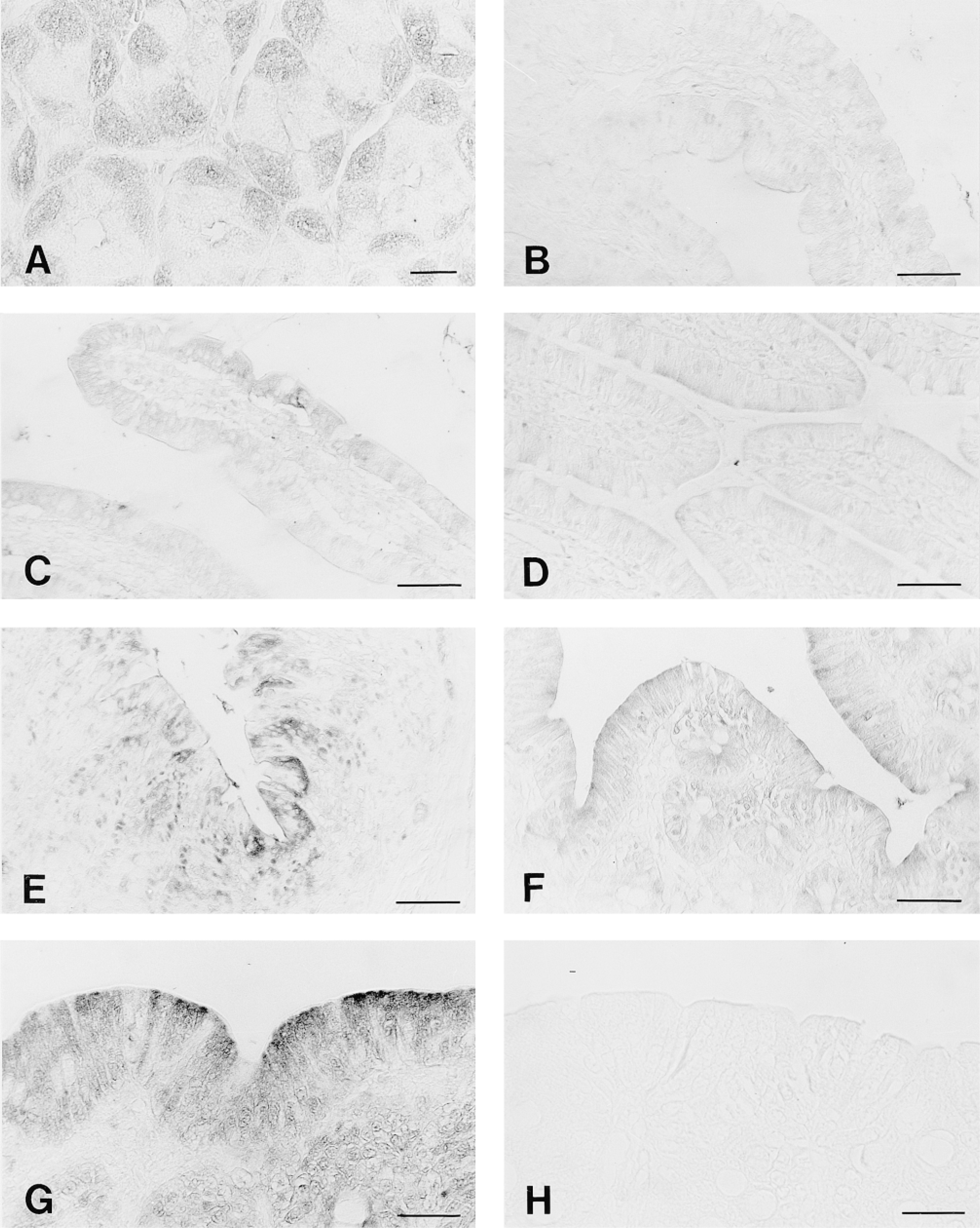
Immunohistochemical demonstration of CA V in the rat gastrointestinal tract. In the body of the stomach, positive immunoreaction is seen in the parietal cells (A). Immunoreactions for CA V are quite weak in the duodenum (B), jejunum (C), and ileum (D). In the cecum (E) and ascending colon (F,G), the positive reactions are distinctly stronger than those in the small intestine and are confined to the superficial enterocytes of the surface epithelial cuff region. A high-magnification view (G) shows that the immunoreaction was mainly fine and granular in each positive enterocyte. Control immunostaining of the ascending colon using nonimmune rabbit serum instead of the anti-CA V antibody showed no positive immunoreaction (H). Bars: A,G-H = 20 μm; B-F = 50 mm.
Discussion
The mitochondrial carbonic anhydrase (CA V) is a unique CA isozyme with respect to its ability to participate in metabolic processes such as gluconeogenesis and urea synthesis in liver (Dodgson et al. 1983; Metcalfe et al. 1985; Dodgson 1991a,b; Sly and Hu 1995; Hazen et al. 1996,1997; Parkkila and Parkkila 1996). Although CA V may function in mitochondria of other tissues as well, its distribution has not been extensively studied. There is only one previous immunohistochemical report demonstrating that CA V is expressed in various tissues of the rat alimentary tract (Väänänen et al. 1991). Nagao et al. (1994) questioned these results because another mitochondrial enzyme, malate dehydrogenase, was shown to co-purify with CA V under conditions used to isolate rat CA V to raise antibodies in the initial study (Ohlinger et al. 1993), and the molecular weight of the protein identified by the antibody was that of malate dehydrogenase (34 kD). In this study, the availability of isozyme-specific antibodies against a C-terminal polypeptide of human CA V and against recombinant mouse CA V produced in bacteria enabled us to compare the immunohistochemical localization of CA V between human and rat tissues and among different segments of the mammalian gastrointestinal tract.
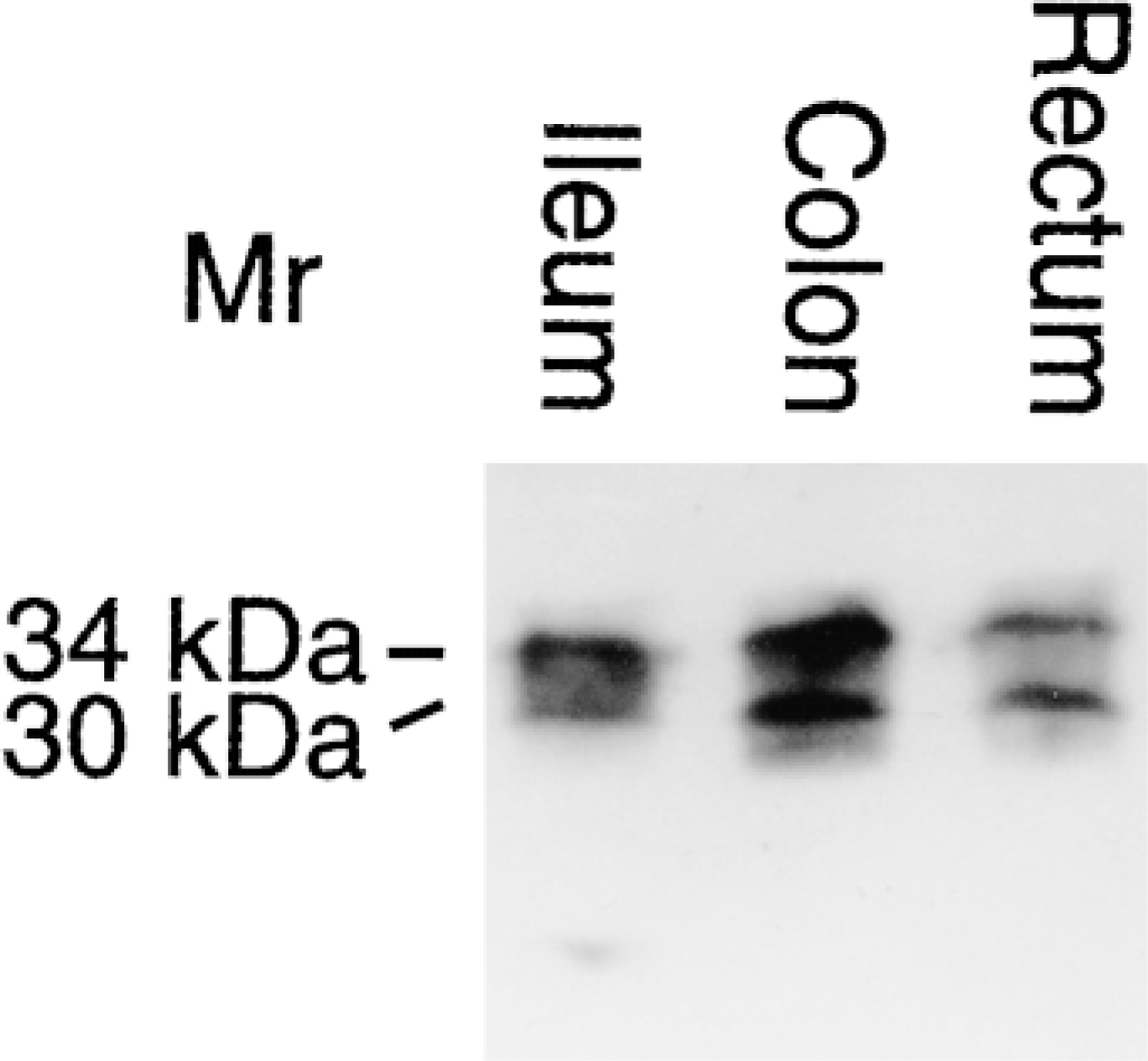
Expression of CA V in the human alimentary tract. Proteins from tissue homogenates of ileum, ascending colon, and rectum were subjected to SDS-PAGE followed by Western blotting. The apparent molecular masses of the precursor (34-kD) and mature (30-kD) CA V polypeptides are indicated.
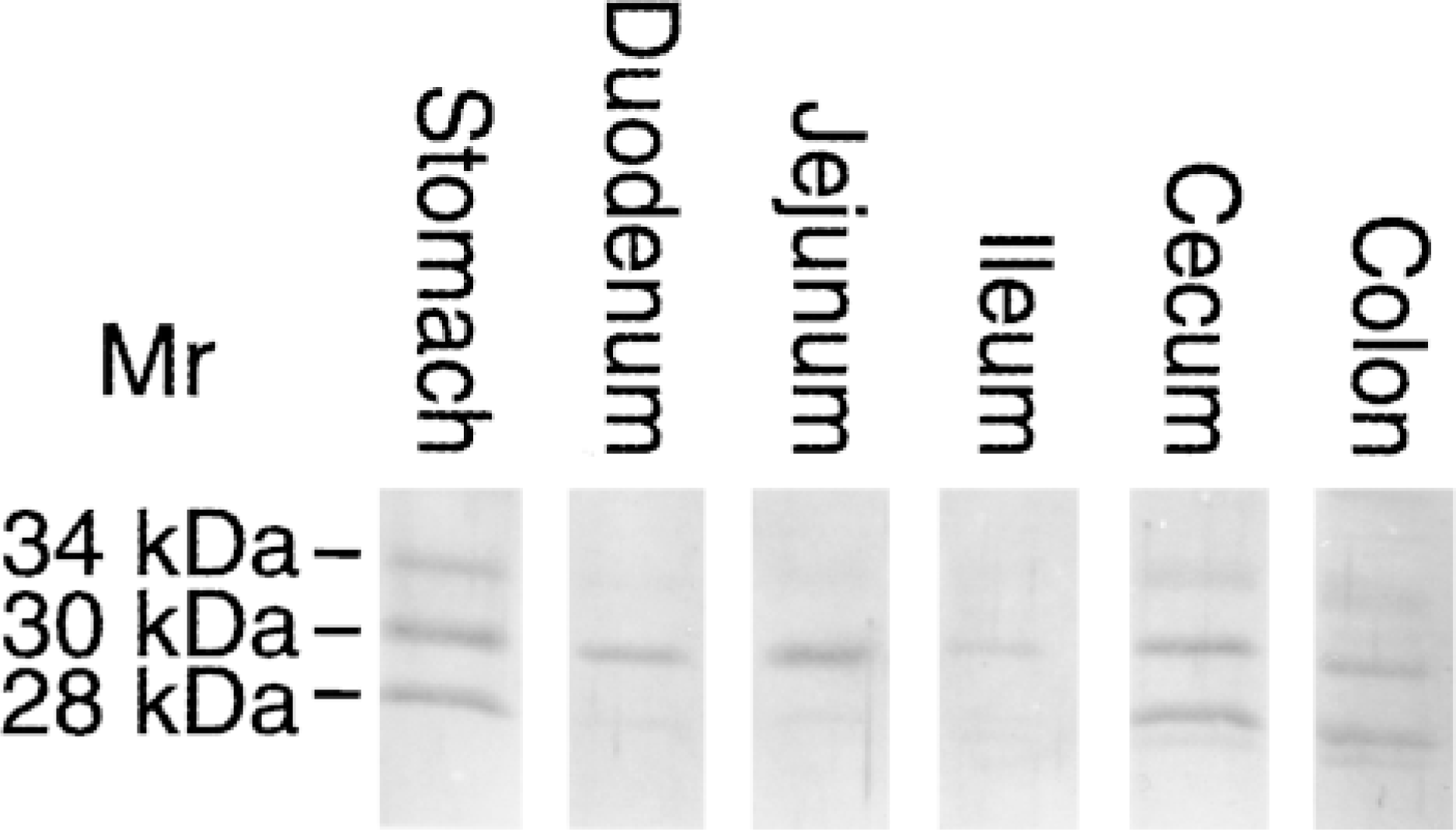
Expression of CA V in different segments of the rat gastrointestinal canal. Tissue homogenates from stomach, duodenum, jejunum, ileum, cecum, and middle colon were analyzed by SDS-PAGE followed by Western blotting. The apparent molecular weights of the precursor (34-kD), mature (30-kD), and proteolytically nicked (28-kD) CA V are marked.
Three major conclusions can be drawn from the present results. First, mitochondrial CA V is widely expressed in all segments of the alimentary canal, from stomach to rectum. Second, the expression patterns of human and rat CA V in the gastrointestinal tract are quite similar. Third, parietal cells and endocrine G-cells of the stomach and enterocytes of the large intestine appear to be the richest sources of CA V in the tissues of the alimentary canal.
The present immunostainings revealed several differences from those in the previous report on the immunolocalization of CA V in rat tissues (Väänänen et al. 1991). Our results showed much less signal in the duodenum than was reported. On the other hand, we found positive immunoreaction in the surface epithelium of the gastric pit region and the gastric G-cells, where Väänänen et al. (1991) did not demonstrate any immunoreactivity. In addition, we have found high expression of CA V in the pancreatic β-cells (Parkkila et al. 1998), although the previous study showed no immunoreaction in the rat pancreas. These discrepancies, together with the report that malate dehydrogenase co-purifies with CA V by the method used in the prior study to prepare antigen to raise the antibody, suggest that the antibody used earlier identified malate dehydrogenase instead of CA V. According to the published cDNA sequence data, the precursor form of CA V is 34 kD; after transit to mitochondria the precursor polypeptide matures to the 30-kD mature mitochondrial CA V (Nagao et al. 1994). Our antibodies recognized both forms on Western blots from most tissues, as well as the 28-kD proteolytic product in rat tissues (Nagao et al. 1994).
The cell-specific expression of CA V suggests its physiological importance in gluconeogenesis and/or ureagenesis in mucosa of the gastrointestinal canal. Prior studies have indeed shown the presence of mitochondrial urea cycle enzyme carbamyl phosphate synthetase I (a companion enzyme of CA V in liver) in the gut epithelium (Ryall et al. 1985,1986; Moorman et al. 1988; Rings et al. 1992; Van Beers et al. 1998). Although Ryall et al. (1985) demonstrated by immunological measurements that rat intestinal mucosa contains 10–20 times less carbamyl phosphate synthetase I than liver, they proposed that the actual concentration of the enzyme in the mitochondrial compartment does not differ greatly between liver and intestine. The lower levels observed in the intestinal mucosa were suggested to be attributable to a smaller proportion of total cell volume occupied by the mitochondria in enterocytes than in hepatocytes and to the greater number of enzyme-expressing cells in liver. They suggested that the concentration of carbamyl phosphate synthetase I in the enterocytes is still sufficient within mitochondria to perform its physiological function. On the basis of earlier studies showing that enterocytes do not contain all the enzymes required for urea synthesis, Ryall et al. (1985) proposed that a complete functional urea cycle may not exist in the intestinal mucosa. However, because the enzymes catalyzing the first two steps of the urea cycle, carbamyl phosphate synthetase I and ornithine transcarbamylase, have been reported to be expressed in the intestinal mucosa, ammonia could be metabolized to citrulline in the gut. The citrulline produced from ornithine and carbamyl phosphate could then be released into the circulation for delivery to other tissues, where it can be converted to arginine (Windmueller and Spaeth 1981). We suggest that the presence of CA V in the enterocytes places it in a strategic site to provide bicarbonate ions to carbamyl phosphate synthetase I and facilitate the conversion of ammonia to citrulline. On this basis, we propose that the physiological role of CA V in the epithelia of stomach and gut is the detoxification and disposal of ammonia produced in the gut.
Footnotes
Acknowledgements
Supported by grants DK40163 and GM34182 from the US Public Health Service to WSS, from the Sigrid Juselius Foundation to SP, and from the Maud Kuistila Foundation and Academy of Finland to A-KP.
We thank Elizabeth Torno for editorial assistance.