
Abstract
Carbonic anhydrase IX (CA IX) is a unique member of the CA gene family. In contrast to the other isozymes, it has been implicated in regulation of cell proliferation, adhesion, and malignant cell invasion. In a recently described knockout mouse model for CA IX deficiency, the only phenotypic abnormalities were limited to the gastric mucosa, while no changes were observed in the other tissues known to express CA IX in rats and humans. Here we investigated the expression of CA IX mRNA and protein in mouse tissues. Immunohistochemical (IHC) analysis showed strong staining in the gastric mucosa. Moderate reactions were seen in the colon enterocytes and pancreatic acini. The expression pattern of CA IX was similar in certain human and rodent tissues, although some differences existed, especially in the gut epithelium. Reverse transcriptase PCR analyses surprisingly revealed strong signals for CA IX mRNA in the kidney and skeletal muscle, while the IHC and Western blotting showed no or weak signals for the corresponding protein. This result suggests a tight tissue-specific post-transcriptional control for CA IX expression, possibly related to the physiological demands.
C
CA IX is composed of four domains: it has an N-terminal proteoglycan domain, a CA catalytic domain, a transmembrane region, and a short cytoplasmic tail (Opavský et al. 1996). CA IX is a sulfonamide-sensitive isozyme with high enzymatic activity, and it is also considered a cell adhesion molecule (Závada et al. 2000; Wingo et al. 2001; Svastová et al. 2003; Vullo et al. 2003). The distribution of CA IX has been studied in human and rat alimentary tracts (Pastoreková et al. 1997). In the human alimentary tract, strong expression of CA IX was detected in stomach and gallbladder. In addition, ileum, colon, liver, and pancreas showed positive expression. Rat alimentary tract showed a slightly different distribution: stomach, colon and duodenum appeared to be positive and liver gave weak positive staining. One of the interesting features of CA IX is its overexpression in human epithelial tumors derived from tissues that normally do not express this isozyme, including carcinomas of uterine cervix, lung, kidney, and breast (Liao et al. 1994; McKiernan et al. 1997; Vermylen et al. 1999; Bartosová et al. 2002). In contrast, tumors originating from CA IX-positive tissues, such as stomach, tend to have lowered expression of CA IX (Leppilampi et al. 2003). It is overexpressed in von Hippel-Lindau (VHL)-defective tumors and under hypoxic conditions (Wykoff et al. 2000; Loncaster et al. 2001; Mandriota et al. 2002). In tumors, CA IX has been proposed to maintain extracellular acidic pH, which is conducive to tumor growth and spread (Ivanov et al. 2001).
The aim of this study was to investigate the expression of CA IX in mouse tissues by reverse transcriptase polymerase chain reaction (RT-PCR), Western blotting, and immunohistochemistry (IHC). This has become an important area of investigation because a knockout mouse model with CA IX deficiency has been recently described (Ortova Gut et al. 2002). The study showed that CA IX deficiency leads to gastric hyperplasia of the glandular epithelium with many cysts. The results suggested that CA IX participates in signal transduction and has an important role in control of cell proliferation, differentiation, and protection of integrity of the stomach mucosa. On the other hand, the lack of CA IX did not have any major physiological consequences on gastric pH regulation or acid secretion, suggesting that other isozymes may compensate for a single enzyme deficiency. However, except for basic data provided therein, no thorough information on the CA IX distribution in mouse tissues is available thus far.
Materials and Methods
Polymerase Chain Reaction (PCR) Method
The expression of CA IX mRNA in mouse tissues was examined using a cDNA kit (MTC panel I) purchased from BD Biosciences (Palo Alto, CA). The mouse MTC panel I contained first-strand cDNA preparations produced from poly A RNAs isolated from 12 different mouse tissues. The first-strand cDNA was the template in the PCR reaction and specific primers for CA IX were used to reveal the tissues containing CA IX mRNA. In addition, primers for mouse β-actin were used to monitor the possible differences in cDNA concentrations.
mRNA was isolated using TRIZOL reagent (Invitrogen; Carlsbad, CA) from the tissues absent from the panel (stomach and gut) as well as from muscle and kidney. Reverse transcription was performed with Mo-MuLV reverse transcriptase (Finnzymes; Espoo, Finland) using random primers (500 μg/ml).
The primers for the PCR reaction were designed by using the published information on CA IX mRNA in GenBank (accession number NM_139305). To produce an amplification product of 873 bp, the forward primer (F1) was 5′-GCTCCAAGATTGAGATC-3′ (nucleotides 894–911) and the reverse primer (R1) 5′-TCTGCCTGCATAGTAAGA-3′ (nucleotides 1749–1766). In the second set of experiments, the forward primer (F2) was 5′-GGAGGCCTGGCAGTTTTGGCT-3′ (nucleotides 794–814) and the reverse primer (R2) 5′-CTCCAGTTTCTGTCATCTCTGCC-3′ (nucleotides 1336–1358). With these primers the PCR amplification product was predicted to be 565 bp. To confirm the results, we also used a third set of primers: (F3) 5′-CTGGGCGTGGATGTTCTGCCC-3′ (nucleotides 212–232) and (R3) 5′-CGAGATTTCTTCCAAATGGGACAG-3′ (nucleotides 863–886), with the resulting PCR product of 675 bp. The primers were produced by Sigma Genosys (Cambridge, UK) and all the other reagents for the PCR reaction were from BD Biosciences except for the dNTP mix, which was from Finnzymes. Five ng of cDNA was used as template. The PCR reaction was carried out on a thermal cycler (Gene Amp PCR system 9700; Applied Biosystems, Foster City, CA) and the protocol consisted of a 94C denaturation step for 1 min followed by 30 cycles of denaturation at 94C for 30 sec, annealing at 55C for 30 sec, and extension at 72C for 1 min 30 sec, followed by a final extension at 72C for 3 min. The control PCR reaction was performed with the following primers for mouse β-actin: the forward primer was 5′-GTTGGCATAGAGGTCTTTACG-3′ and the reverse primer was 5′-GCCGCATCCTCTTCCTCCCT-3′. The control reaction was carried out according to the following protocol: 94C denaturation step for 1 min followed by 30 cycles of denaturation at 94C for 30 sec, annealing at 60C for 30 sec, and extension at 72C for 40 sec, followed by final extension at 72C for 3 min. The results of the PCR reaction were analyzed using a 1.2% agarose gel containing 0.1 μg/ml ethidium bromide with DNA standard (100-bp DNA Ladder; New England Biolabs, Beverly, MA).
Sequencing of the PCR Products
The PCR products from mouse muscle and kidney were sequenced to confirm the presence of the correct amplification product. The PCR products were purified with a GFX PCR DNA and Gel Band Purification Kit (Amersham Biosciences; Poole, UK) following the protocol of the manufacturer. The sequencing was performed using ABI PRISM Big Dye Terminator Cycle Sequencing Ready Reactions Kit, version 2.0 (Applied Biosystems). Five μl of DNA template was mixed with 4 μl of Terminator Ready Reaction Mix (Applied Biosystems) and 1.6 pmol of primer was added to the solution. The sequencing was performed with both forward and reverse primers. The reactions were amplified by cycle sequencing on a thermal cycler (Gene Amp PCR system 9700; Applied Biosystems) according to the manufacturer's instructions. The products were purified by ethanol precipitation, resuspended in Template Suppression Reagent (Applied Biosystems), and denatured according to the manufacturer's protocol. The sequencing was performed with an ABI PRISM Genetic Analyser instrument (Applied Biosystems).
Western Blotting
Samples of stomach, colon, duodenum, jejunum, ileum, brain, cerebellum, heart, liver, kidney, pancreas, lung, spleen, thymus, psoas muscle, testis, and epididymis were obtained from adult mice (Balb/c). The procedures were approved by the institutional animal care committee (University of Tampere). The tissue samples were homogenized in PBS and 50 μg of protein from each sample was analyzed by SDS-PAGE (NuPAGE 10% Bis-Tris; Invitrogen) under reducing conditions.
The separated proteins were transferred electrophoretically from the gel to a polyvinylidene fluoride (PVDF) membrane (Macherey-Nagel; Düren, Germany) in a Novex Xcell II blot module (Invitrogen). The sample lanes were detected by an ECL detection system (Amersham Biosciences). The membranes were blocked with cow colostral whey (Biotop Oy; Oulu, Finland) diluted 1:10 in TBST buffer (20 mM Tris, 500 mM NaCl, 0.3% Tween-20, pH 7.5) for 25 min. The membranes were then incubated with primary antibody diluted 1:5000 in TBST buffer for 1 hr and washed five times for 5 min in TBST buffer. The production of polyclonal rabbit antibody raised against the recombinant mouse CA IX has been described by Ortova Gut et al. (2002). Normal rabbit serum was used for control purposes. The second antibody was horseradish peroxidase-labeled and the rest of the procedure was performed according to the manufacturer's instructions (Amersham Biosciences).
Immunohistochemistry
Immunoperoxidase staining for mouse tissues was performed using the peroxidase-antiperoxidase (PAP) complex method. A polyclonal rabbit antibody against the recombinant mouse CA IX was used to detect CA IX in tissue sections. Normal rabbit serum was used in the control stainings.
Tissue specimens from brain, stomach, duodenum, jejunum, ileum, colon, liver, psoas muscle, kidney, heart, lung, pancreas, spleen, thymus, testis, and epididymis were obtained from two adult mice (Balb/c). The specimens were fixed in Carnoy's fluid (absolute ethanol + chloroform + glacial acetic acid in 6:3:1 ratio) for 20 hr at 4C. The samples were then dehydrated, embedded in paraffin, and 4-μm sections were cut and placed on microscope slides.
Immunostaining was performed according to the following protocol: (a) 3% H2O2 in methanol for 5 min; (b) treatment with undiluted cow colostral whey (Biotop) for 30 min and rinsing in PBS; (c) incubation for 1 hr with primary antibody diluted 1:100 in 1% bovine serum albumin (BSA) in PBS; (d) treatment with undiluted cow colostral whey for 30 min and rinsing in PBS; (e) incubation for 1 hr with second antibody (swine anti-rabbit IgG; DAKO, Glostrup, Denmark) diluted 1:100 in 1% BSA in PBS; (f) incubation with peroxidase-antiperoxidase complex (PAP Rabbit; DAKO) diluted 1:100 in PBS for 30 min; (g) incubation for 2 min 30 sec in DAB solution (6 mg 3,3′-diaminobenzidine tetrahydrochloride; Sigma, St Louis, MO) in 10 ml PBS plus 3,3 μl 30% H2O2; and (h) counterstaining with Mayer's hematoxylin solution (Sigma). The sections were washed in PBS for 5 min after the step a, 3 times for 10 min after steps c and e, and finally four times for 5 min after step f. All incubations and washings were carried out at room temperature. In addition, each washing was performed on an orbital shaker. Finally, the sections were mounted in Entellan Neu (Merck; Darmstadt, Germany) and then examined and photographed with a Zeiss Axioskop 40 microscope (Carl Zeiss; Göttingen, Germany).
Results
PCR and Sequencing
The expression of CA IX mRNA in mouse tissues was studied using a commercially available cDNA kit as well as the cDNAs reverse-transcribed from mRNA that we isolated from selected mouse tissues. The mRNA was found in the kidney and muscle and weak bands were also seen in the brain, spleen, lung, and liver (Figure 1; Table 1). Heart and testis, as well as 7-day-old, 11-day-old, and 15-day-old embryos were negative. Embryos of 17 days appeared to express CA IX mRNA, which is in accordance with a recent study showing low levels of CA IX protein expression during the first postnatal day (Ortova Gut et al. 2002). In the gastrointestinal tract, mRNA was expressed in the stomach, duodenum, jejunum, ileum, and colon (Figures 1 and 3).
Because only low amounts of CA IX mRNA have been reported in kidney and muscle (Ivanov et al. 2001; Ortova Gut et al. 2002), the PCR products from these organs were further sequenced to confirm their specificity. The automated sequencing was performed using primers designed for both 5′ and 3′ ends and the sequencing clearly confirmed that the obtained 873-bp band represented the correct amplification product.
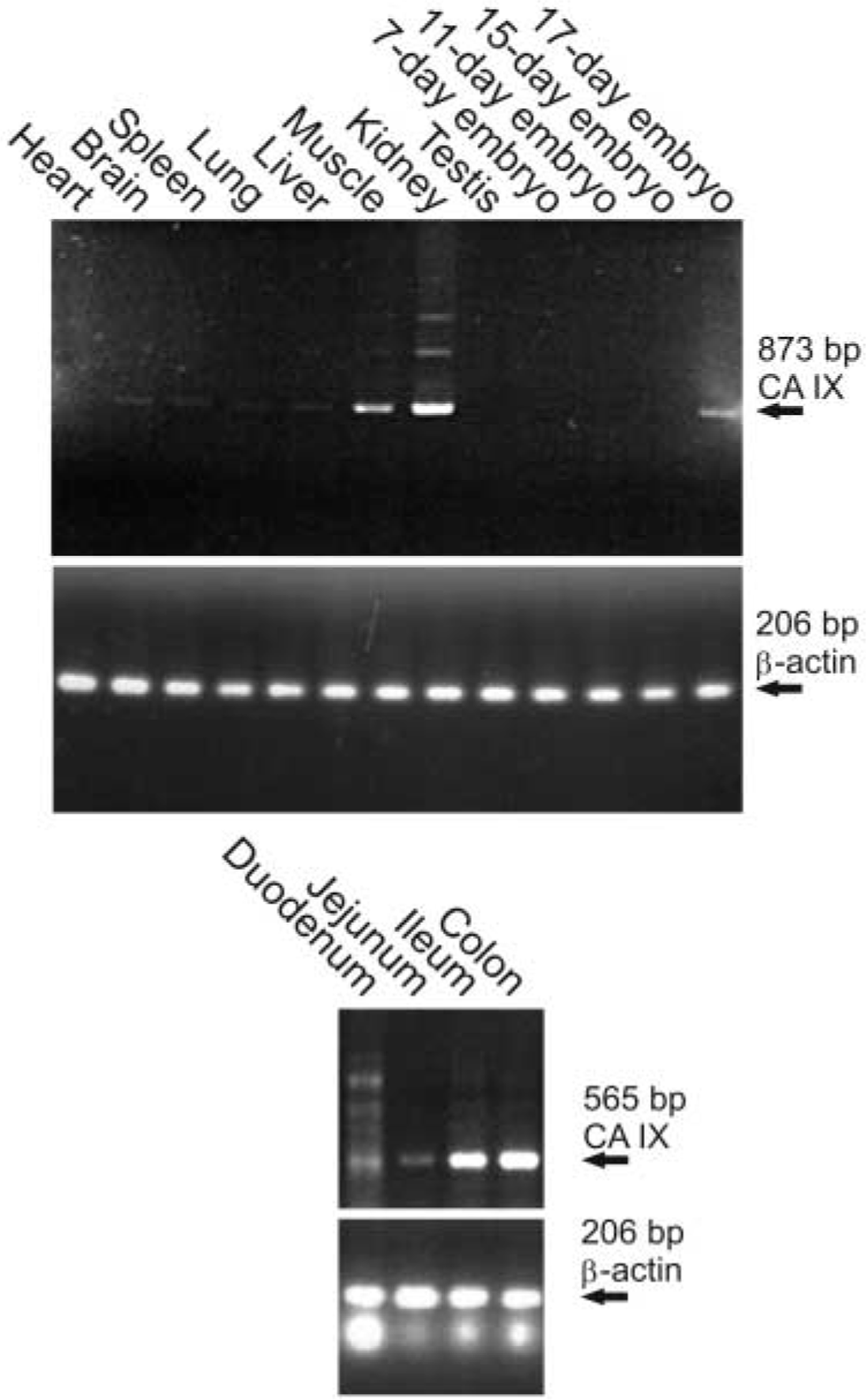
The PCR product of 873 bp is amplified in the kidney and muscle, followed by brain, spleen, lung, and liver. Testis and heart are negative. The expression of CA IX mRNA becomes positive in the 17-day-old embryo. The results in the lower panel show that CA IX mRNA is expressed in the colon and ileum, and weaker bands are also seen in the duodenum and jejunum. The amount of cDNAs produced from each tissue was monitored using primers for β-actin.
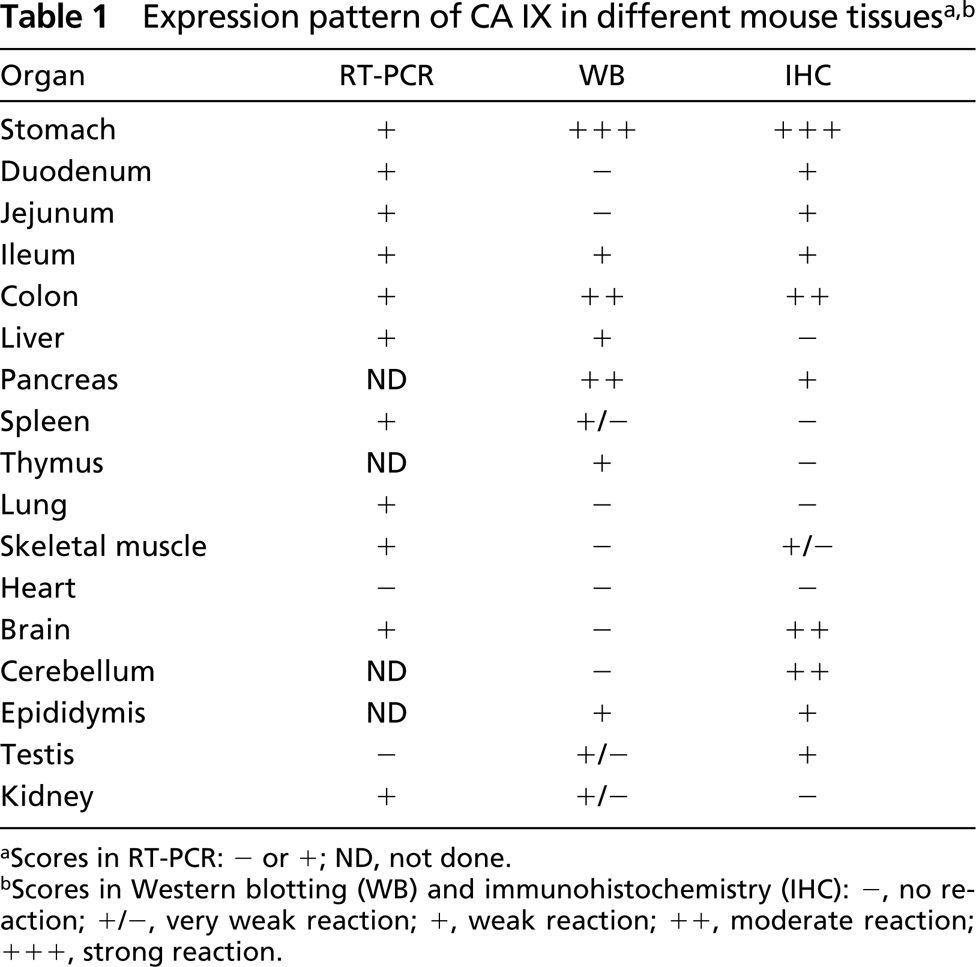
aScores in RT-PCR: − or +; ND, not done.
bScores in Western blotting (WB) and immunohistochemistry (IHC): −, no reaction; +/−, very weak reaction; +, weak reaction; + +, moderate reaction; + + +, strong reaction.
Western Blotting
Western blotting was performed to screen CA IX protein expression in 17 different mouse tissues. As expected, the highest signal for this protein was detected in the stomach (Figure 2; Table 1). Colon and pancreas showed moderate positive reactions. Very faint bands were detected in the kidney, liver, thymus, testis, epididymis, ileum, and spleen. Jejunum, duodenum, psoas muscle, heart, lung, brain, and cerebellum showed no CA IX-specific band. Smaller polypeptide bands were observed in the ileum, jejunum, and colon (~37 kD) and in stomach and pancreas (~30 kD). These polypeptides may represent CA IX fragments produced by degradation.
To confirm the differential expression of CA IX mRNA vs protein in the kidney and muscle, the same tissues (excised from two mice) were processed in parallel for RT-PCR and Western blotting (Figure 3). Results of RT-PCR performed with a different pair of primers corresponded well to those obtained in previous analyses and shown in Figure 1. Similarly, a profile of the proteins detected by Western blotting in the same tissues as used for RT-PCR largely agreed with the data in Figure 2. The CA IX-specific protein band was evident in the stomach and colon and was very weak also in the small intestine, but was completely absent from the kidney and muscle.
Immunohistochemistry
Immunoperoxidase staining was performed to localize the expression of CA IX in different tissues and cell types. A polyclonal rabbit antibody for mouse CA IX was used to detect this enzyme and pre-immune serum was used for control purposes. Because CA IX has been reported to be present in the gastric mucosa (Ortova Gut et al. 2002), stomach specimens served as a positive control tissue for the immunostaining. Strong reaction was seen in the basolateral plasma membranes of the mucus-producing surface epithelial cells, chief cells, and parietal cells (Figure 4; Table 1). In the colon, the signal was present in the plasma membrane of the enterocytes and the strongest immunoreaction was localized to the surface epithelial cuff region. In the small intestine, the staining was very weak and present only in sporadic enterocytes. The pancreas showed moderate positive reaction in the acinar cells, in which the staining was generally diffuse, most probably due to rapid protein degradation in the tissue samples (Figure 5). No staining for CA IX was detected in the liver. In the skeletal muscle, only very weak immunoreactions were seen in occasional fibers, while the heart muscle was negative. Faint positive signal was also observed in the epithelial cells of the epididymis and in mature spermatozoa located in the seminiferous tubules. No staining for CA IX was found in the kidney, lung, thymus, and spleen. The antibody also labeled some neuronal axons in the brain and Purkinje cells of the cerebellum (Figure 6).
Discussion
CA IX is an interesting and unique member of the CA gene family with respect to its molecular structure, distribution, and suggested functions. First, it contains two structurally distinct domains, a CA domain and a proteoglycan-like domain, which might contribute to different functions (Opavský et al. 1996). Second, human CA IX has been reported to be present in only a few normal tissues and is overexpressed in some carcinomas (Závada et al. 1993). Third, its expression is induced by hypoxia and is downregulated by the product of the wild-type von Hippel-Lindau gene (Ivanov et al. 1998; Wykoff et al. 2000). Fourth, recent studies in CA IX knockout mice have further shown that CA IX is functionally involved in normal gastric morphogenesis, and this feature in particular makes it a very distinctive isoform among the other members of the CA gene family (Ortova Gut et al. 2002).
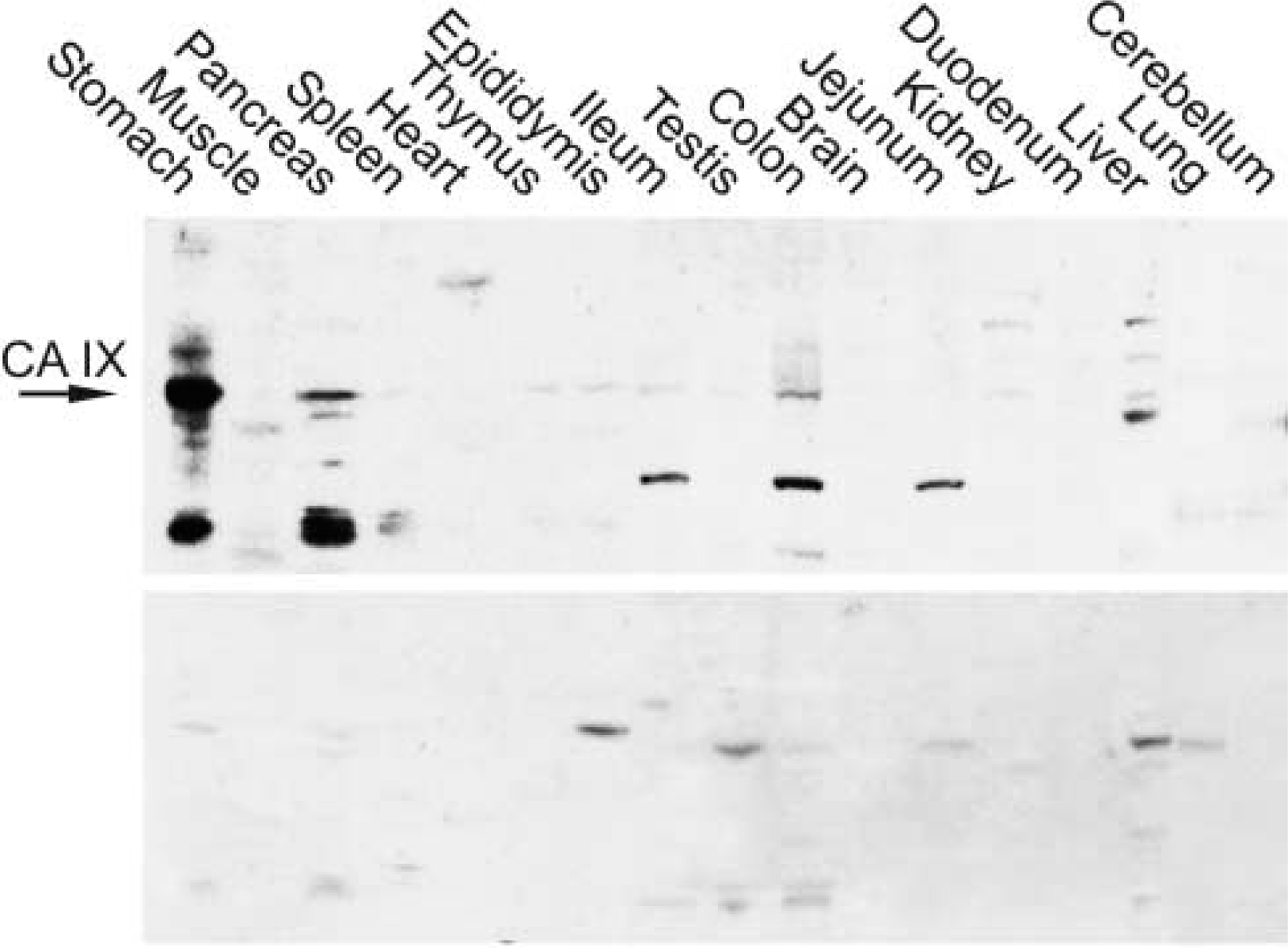
Western blotting of mouse tissues. The strongest 47-kD polypeptide of CA IX is detected in the stomach, followed by pancreas and colon. Very faint bands are seen in the kidney, liver, thymus, testis, epididymis, ileum, and spleen. In addition, strong polypeptides of ~37 kD are present in the ileum, jejunum, and colon and of ~30 kD in the stomach and pancreas. Control staining with normal rabbit serum also shows some nonspecific bands but they all are of different molecular weight compared with CA IX.
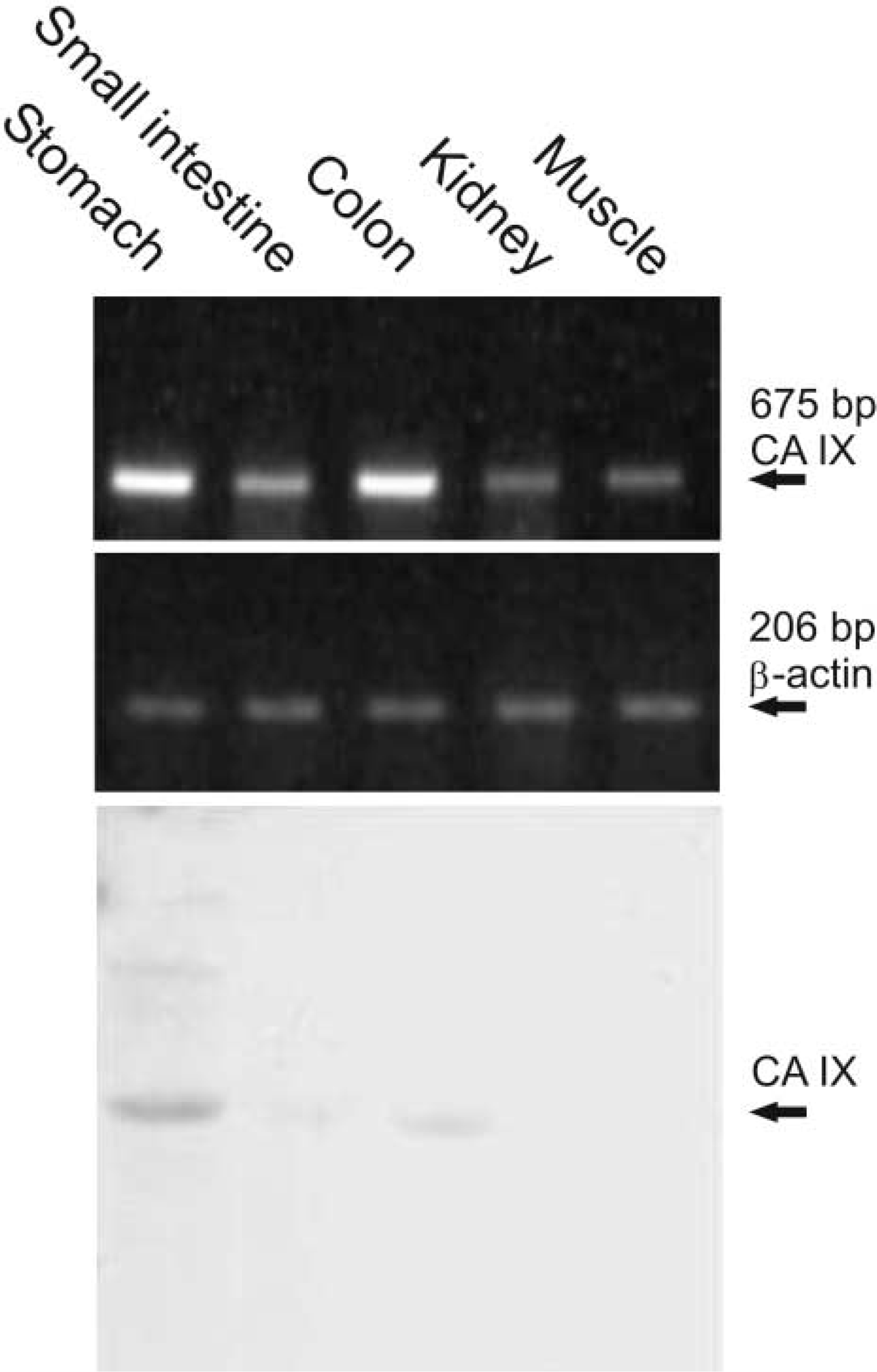
Comparison of the RT-PCR (first panel) and Western blotting (third panel) performed for CA IX in parallel from the same tissues. Note the discrepancy in the signals obtained by Western blotting and RT-PCR in the kidney and muscle.
Even though the expression of CA IX has been quite well established in human and rat tissues (Pastoreková et al. 1997), only little has been known about its expression in mouse. The results obtained in this study (summarized in Table 1) provide a basic overview of CA IX distribution in mouse tissues. Our findings indicate that, among a number of different tissues, gastric mucosa shows the highest expression of this enzyme. The same phenomenon has been described earlier in human and rat tissues. On the basis of both RT-PCR and Western blotting results, the colon and pancreas clearly expressed CA IX. IHC confirmed that colon enterocytes and pancreatic acini were indeed positive for CA IX immunostaining. Some other tissues, including kidney, liver, thymus, testis, epididymis, ileum, and spleen, also showed weak signals for CA IX in the Western blot, but the intensity was very weak, suggesting a low expression of the enzyme in those tissues. The PCR analysis included mouse tissues that were either available in the commercial cDNA kit or obtained from mice in our laboratory. Unexpectedly, the kidney and muscle showed strong positive signal for CA IX mRNA, while the control amplification using β-actin primers produced steady signal in all tissues. Furthermore, the specificity of the positive amplification products was confirmed by DNA sequencing. IHC and Western blotting, however, showed no or very weak reactions for CA IX in the kidney and skeletal muscle fibers. To confirm a differential expression of CA IX mRNA vs protein in the kidney and muscle, we performed RT-PCR and Western blotting analyses from identical tissue specimens. We again found the presence of CA IX mRNA and absence of the corresponding protein, confirming that the discrepancy was not due to inter-animal differences.
These results raise two basic questions: first, why CA IX transcription is activated particularly in the muscle and kidney tissues and, second, why there is a contradiction between expression of mRNA and protein. One possible answer for the first question may be related to the presence of a functional hypoxia-response element in the promoter of the mouse Car9 gene (unpublished results), which may allow transcription of CA IX under conditions of lowered oxygen and/or hypoglycemia, similarly as in human tissues. It is quite conceivable that such conditions may be locally present, at least in the muscle. Although this idea is indirectly supported by our finding of considerably increased levels of PCR product specific for the hypoxia-regulated glyceraldehyde-3-phosphate dehydrogenase (GAPDH) gene in both muscle and kidney (data not shown), definitive evidence requires further investigation. The response to the second question is also unclear but appears to point at a tissue-specific post-transcriptional regulation. As proved by RT-PCR amplification using different pairs of primers designed to cover whole mRNA, CA IX transcripts in kidney and muscle are intact. Therefore, the reason for the absence of corresponding protein does not appear to reside in production of incomplete polypeptide committed to degradation or of a truncated secretory form. Our results suggest that the translation efficiency of CA IX mRNA can be very low in certain cell types based on the physiological demands of each tissue. It is noteworthy that normal human kidney and muscle show no or only very weak expression of both CA IX protein and mRNA (Liao et al. 1997; McKiernan et al. 1997; Ivanov et al. 2001). Therefore, an interesting area for future investigations would be exploration of the regulatory mechanisms of CA IX mRNA translation in the murine kidney and muscle.
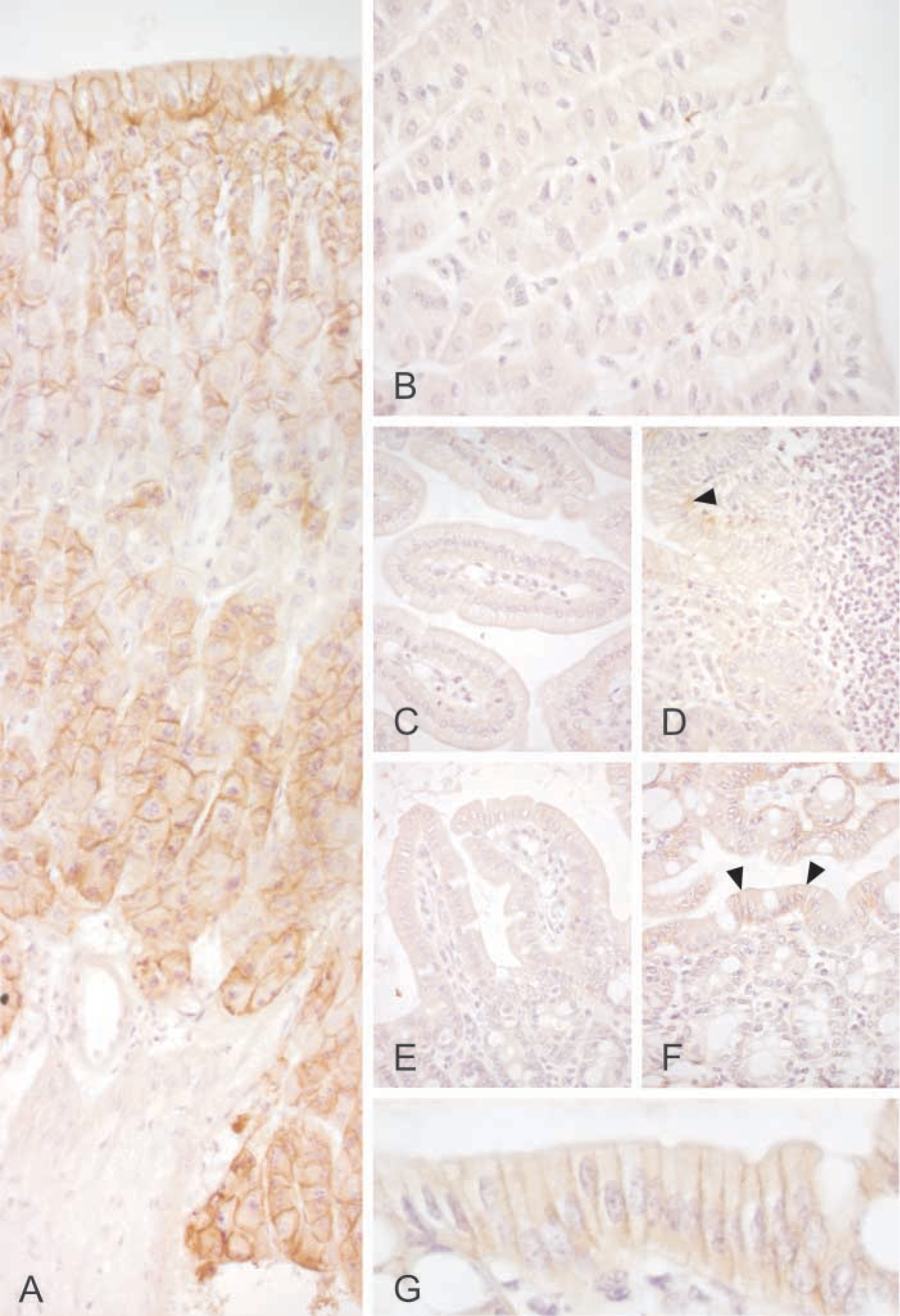
Immunohistochemical staining of CA IX in the mouse gastrointestinal tract. Stomach shows the most intense staining (
The present results provided a good opportunity to compare the CA IX distribution pattern among different species. We have previously shown that it is expressed in the rat stomach, duodenum, colon, and biliary tract as well as the human stomach, duodenum, jejunum, ileum, colon, biliary tract, and pancreas (Pastoreková et al. 1997; Saarnio et al. 1998). The distribution in the mouse gastrointestinal tract was quite similar to that of the rat and human tissues. In all species, gastric mucosa appeared to contain the highest levels of CA IX. Therefore, it is not surprising that CA IX knockout mice showed a distinct gastric phenotype with epithelial cell hyperplasia and cystic changes in the mucosa (Ortova Gut et al. 2002). Based on the previous and present studies, CA IX is expressed in the colon of all these species. Nevertheless, CA IX knockout mice exhibited no intestinal phenotype that might be partly explained by much lower level of expression in the gut compared with the gastric mucosa. It is also notable that CA IX immunostaining clearly shows different regional distribution in the rodent vs human colon (Pastoreková et al. 1997). In the rat and mouse colon, the enzyme is located in the most superficial part of the mucosa, called the epithelial cuff region. In contrast, the human colon shows positive signal only in the crypt enterocytes. This finding may reflect important differences in colon physiology among various species with respect to the function of CA IX. The present results also showed a moderate immunoreaction for CA IX in the pancreatic acini. This finding is in line with the previous observation in the human pancreas showing positive staining in the acinar cells (Kivelä et al. 2000). The major difference is that CA IX is restricted to occasional acini in the human pancreas, whereas the mouse pancreas shows a more diffuse staining.
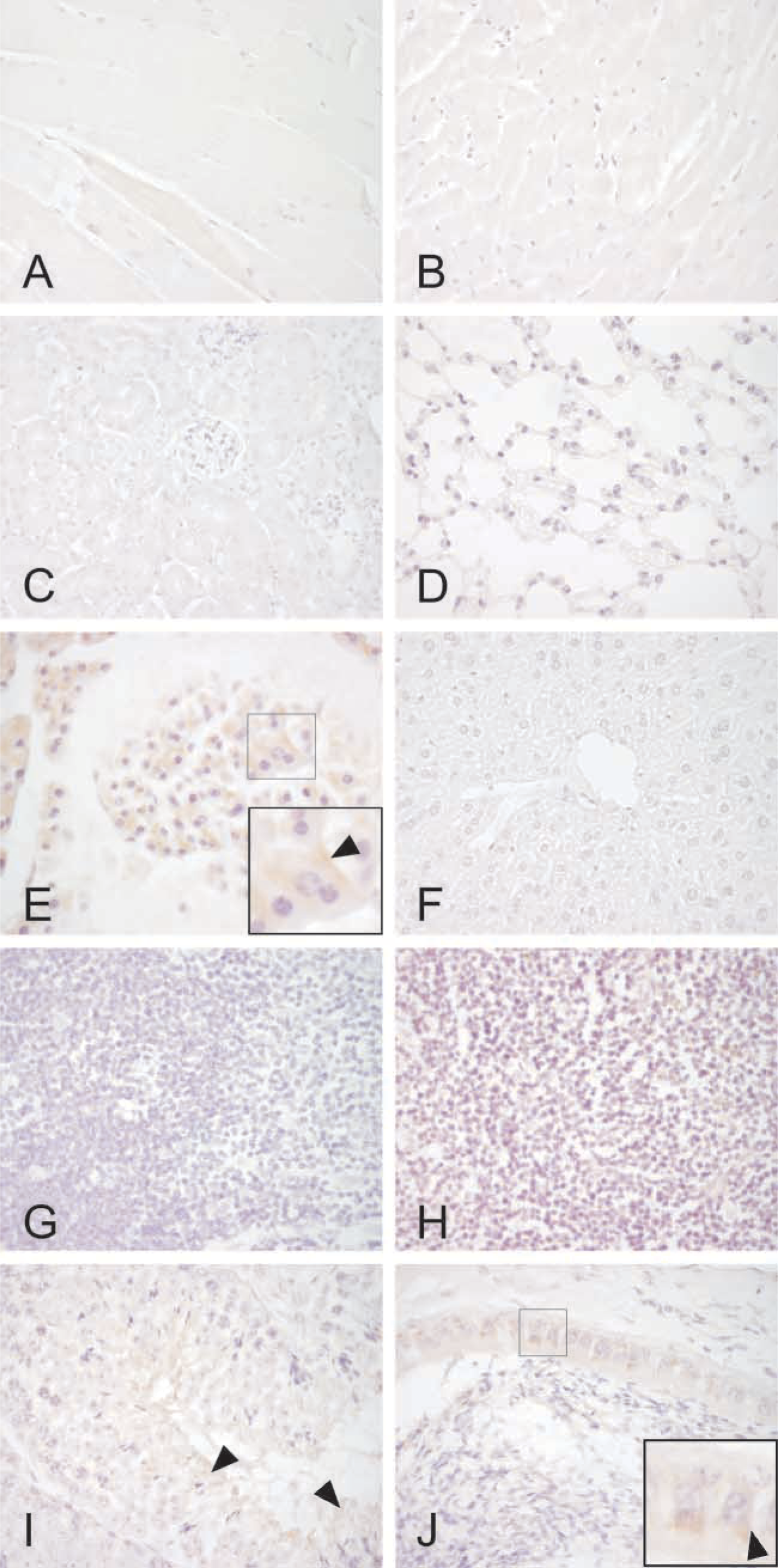
Immunohistochemical staining of CA IX in different mouse tissues. Psoas (
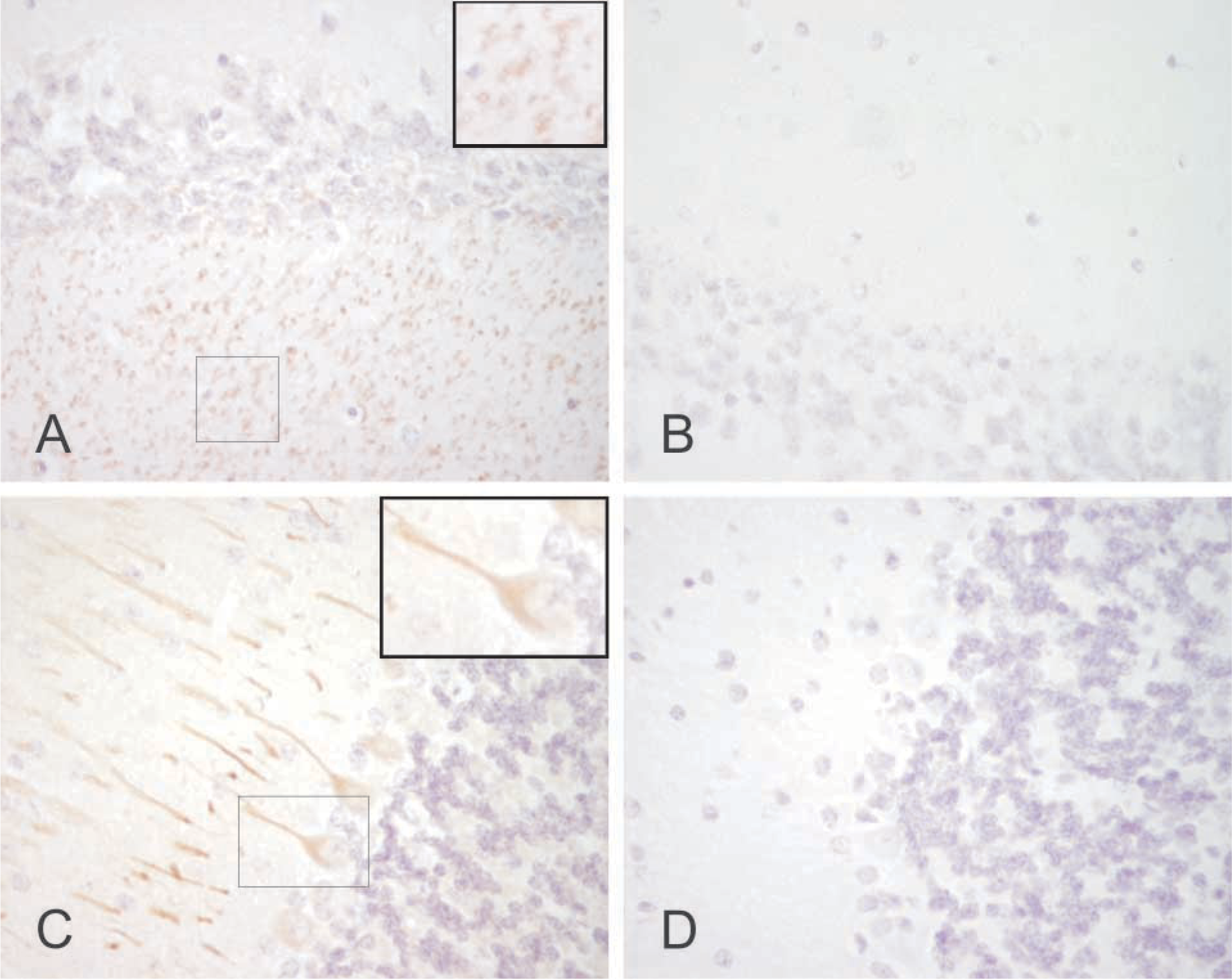
Immunostaining of CA IX in the mouse brain. Some neuronal axons are stained in the cerebrum (
The IHC also provided evidence that CA IX might be expressed in some neuronal axons and Purkinje cells. Weak signal was also observed for CA IX mRNA in the brain, while the Western blots of the brain and cerebellum remained negative, possibly due to lower detection sensitivity. We have previously demonstrated the presence of CA XIV in the human and mouse brain (Parkkila et al. 2001). The present findings suggest that CA IX might be a second membrane-associated isozyme specifically expressed in some neurons. Similar to our present findings, CA XIV showed the highest expression in the axon membrane. The membrane-bound CA activity has been proposed to participate in the production of an alkaline shift linked to neuronal signal transduction (Parkkila et al. 2001). It can be hoped that future studies will unravel the contribution of each CA isozyme to neuronal function.
Footnotes
Acknowledgements
Supported by grants from the Sigrid Juselius Foundation, the Academy of Finland, Bayer Corporation, and Slovak Grant Agencies VEGA (2/3055) and APVT (51–005802).