
Abstract
Carbonic anhydrase isoenzyme VI (CA VI), which is transported in high concentrations in saliva and milk into the alimentary tract, is an important element of mucosal protection in the upper alimentary tract. Like alimentary tract mucosa, the respiratory tract mucosa is also exposed to heavy microbial, physical, and chemical stress. The protective and renewal-promoting factors present in the surface mucus of the respiratory tract are mainly produced by the seromucous tracheobronchial glands. Here we studied the secretion of CA VI by these glands in adult and developing rats using immunohistochemical techniques. The serous acinar and duct cells of the tracheobronchial glands stained for CA VI. The presence of the enzyme also in the duct content indicates its active secretion into the surface mucus. CA VI was also visible in the secretory cells and at the base of the ciliated cells of the tracheobronchial surface epithelium. Moreover, the Clara cells of the bronchiolar surface epithelium stained for CA VI. These findings are consistent with the hypothesis that CA VI has a mucosa-protective role not only in the gastrointestinal tract but also in the respiratory tract, where CA VI may act as a pivotal pH neutralizer and growth factor.
C
Mucosa of the lower airways, similarly to the upper alimentary tract mucosa, is exposed to heavy microbial, physical, and chemical stress and is covered by protective mucus produced by the seromucous tracheobronchial glands and by the surface epithelium (Quinton 1979; Tam and Verdugo 1981; Gail and Lenfant 1983; Basbaum et al. 1990). Expression of CA VI in the lower airways suggests that it has a general mucosa-protective and renewing role not only in the alimentary tract but also in the respiratory tract. CA activity has been demonstrated earlier in the tracheobronchial glands, goblet cells, and some other unidentified epithelial cells of the guinea pig trachea, but the only CA isoenzyme identified there to date is CA II in the tracheobronchial glands (Spicer et al. 1982; Okamura et al. 1996). Therefore, the present study aimed to elucidate whether CA VI is expressed in the lower airway epithelia.
Materials and Methods
Antibodies and Chemicals
Rhodamine-conjugated swine anti-rabbit immunoglobulins were obtained from DAKO (Glostrup, Denmark). Polyclonal rabbit antiserum to human salivary α-amylase, peroxidase-conjugated goat anti-rabbit IgG, biotin-conjugated swine anti-rabbit IgG, and streptavidin-conjugated peroxidase were obtained from Sigma Chemicals (St Louis, MO). 3,3'-Diaminobenzidine tetrahydrochloride (DAB) and Tween-20 were obtained from Fluka (Buchs, Switzerland). Complete protein inhibitor tablets were purchased from Boehringer Mannheim (Mannheim, Germany). Production and characterization of polyclonal rabbit anti-rat CA II and CA VI sera have been described earlier (Kaunisto et al. 1995; Leinonen et al. 2001).
Tissue and Saliva Samples
Tracheal and lung samples were collected from three fetuses and 1-, 2-, 5-, 10-, 20-, 30-day-old and adult (60- to 90-day-old) male Sprague-Dawley rats, fixed in Carnoy's fluid or in 4% paraformaldehyde for 18 hr at 4C, and embedded in paraffin. Sections of 5 μm were subjected to immunohistochemical (IHC) stainings. To confirm that saliva contamination does not cause positive CA VI staining in the respiratory tract, samples from sialoadenectomized rats were also subjected to CA VI staining. The salivary glands were removed surgically from six adult male rats under fentanyl fluanisone–midazolam anesthesia. Three of them were sacrificed on the third postoperative day and another three on the seventh postoperative day. The tracheas and lungs were dissected out and subjected to IHC stainings as described above. Rat saliva was collected from anesthetized animals injected IP with 6 mg/kg of pilocarpine. All animal experiments described had the approval of the Animal Care and Use Committee of the University of Oulu.
Absorption of the Anti-rat CA VI Serum with CA VI
To further confirm the specificity of the CA VI staining, anti-rat CA VI serum was absorbed with purified rat CA VI. Rat saliva samples were centrifuged for 10 min at 13,000 × g at 4C. A volume of 500 μl of the supernatant was incubated in constant rotation for 1 hr at 20C with 100 μl of inhibitor affinity gel matrix supplemented to 1 ml with buffer containing 0.1 M Tris-SO4 and 0.2 M sodium sulfate and Complete protease inhibitors dissolved and diluted according to the manufacturer's instructions at pH 7.0. The affinity gel matrix was prepared by coupling the CA inhibitor p-ami-nomethyl benzenesulfonamide to CM Bio-Gel A according to Khalifah et al. (1977). After incubation the gel was washed three times with 1.5 ml of the above-mentioned buffer. The gel was then incubated for 1 hr with anti-rat serum diluted 1:200 in 1% (w/v) BSA-PBS to absorb the CA VI antibodies from the serum to the inhibitor-bound CA VI. The gel was then centrifuged and the supernatant containing the serum devoid of anti-rat CA VI serum was used for IHC stainings. To confirm that the absorbed anti-rat CA VI serum was devoid of antibodies to CA VI, the antibodies bound to CA VI were eluted from the inhibitor affinity gel matrix using 50 μl of specific elution buffer containing 0.4 M NaN3, 0.1 M Tris-SO4, 0.1 mM benzamidine, and 20% (v/v) glycerol, pH 7.0. Aliquots (1 μl) of the eluate were subjected to immunoblotting with either rabbit anti-rat CA VI serum, absorbed anti-rat CA VI serum, or normal serum using dilution 1:10,000 in 0.1% (v/v) PBS–Tween-20. Immunoblotting in conjunction with anti-rat CA VI serum revealed that the eluate from the inhibitor affinity matrix contained the expected CA VI polypeptide bands of 41 kD (glycosylated form) and 37 kD (partially glycolysated form) and a CA VI antibody band (IgG) (Figure 1). Only one band representing IgG was detected with the absorbed serum and normal serum, indicating that the absorbed and normal sera lacked CA VI antibodies (Figure 1).
Immunohistochemical and Immunofluorescence Stainings
The following polyclonal rabbit antisera were used: rabbit anti-rat CA II serum, rabbit anti-human salivary α-amylase, rabbit anti-rat CA VI serum, normal rabbit serum to anti-rat CA VI serum, and rabbit anti-rat CA VI serum absorbed with purified rat CA VI. The sections used for IHC staining were first deparaffinized and pretreated with 3% (v/v) H2O2 for 5 min, followed by incubation with 1% (w/v) BSA-PBS for 30 min, rinses with PBS, and incubation with 30 μl of antiserum diluted 1:200 in 1% (w/v) BSA-PBS. The sections were then washed three times for 5 min with PBS and incubated for 1 hr with biotin-conjugated swine anti-rabbit IgG diluted 1:300 in 1% (w/v) BSA-PBS. This was followed by three washes with PBS, incubation with streptavidin-conjugated peroxidase diluted 1:600 in PBS for 30 min, and three washes with PBS. Finally, the sections were incubated for 3 min with a solution containing 9 mg of DAB in 15 ml of PBS + 5 μl of 30% (v/v) H2O2 and rinsed in PBS. The immunofluorescence sections were deparaffinized and pretreated with 1% (w/v) BSA-PBS for 30 min. The sections were then rinsed with PBS and incubated with 30 μl of anti-serum diluted 1:200 in 1% (w/v) BSA-PBS. The sections were then washed three times for 10 min with PBS and incubated for 30 min with rhodamine-conjugated swine anti-rabbit immunoglobulins diluted 1:300 in PBS. Finally, the sections were washed four times for 5 min with PBS. All sections were examined with a Nikon Eclipse E600 microscope and photographed with a Nikon Coolpix 950 digital camera. The images were further processed with Corel Draw and Adobe Photoshop computer software.
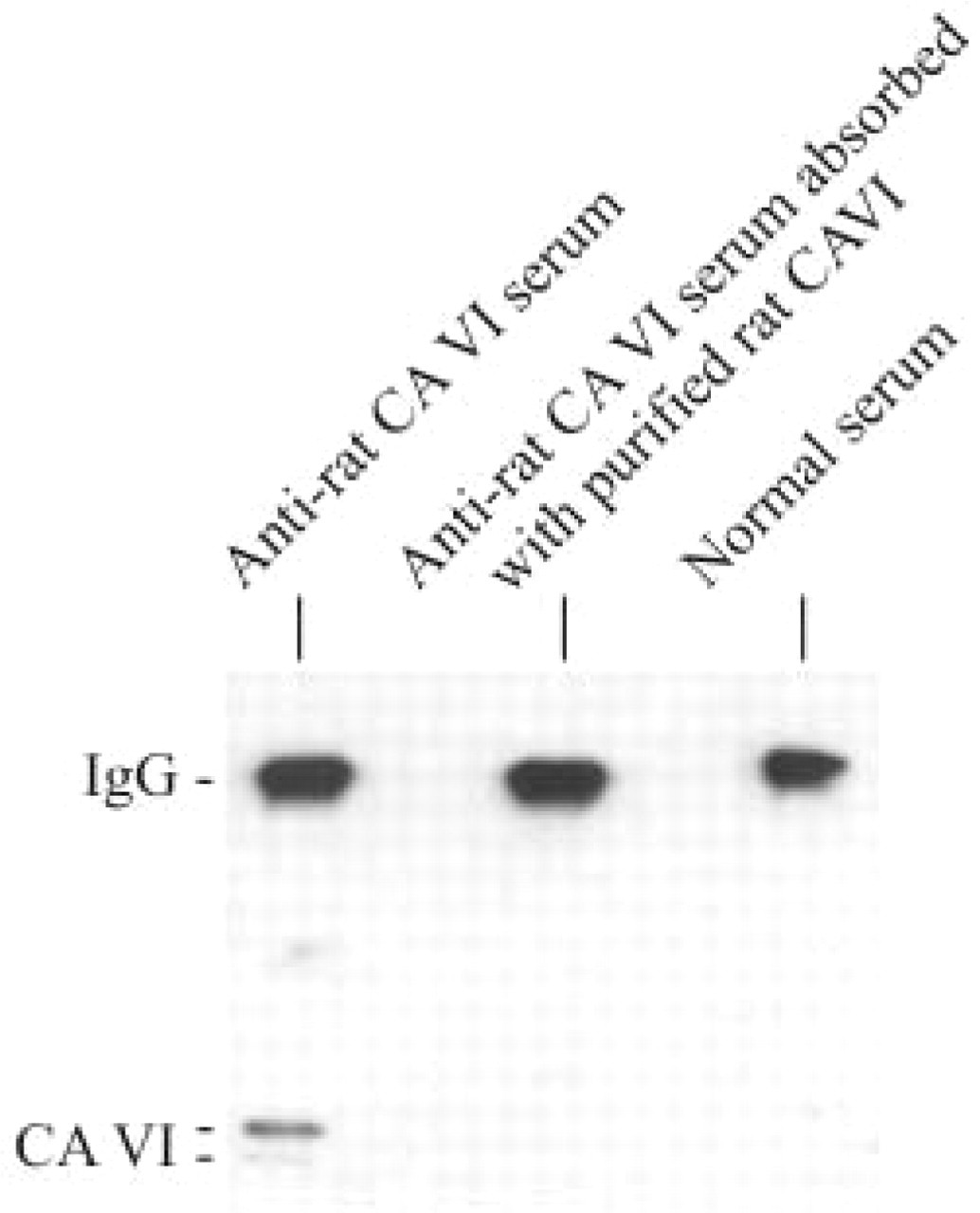
Samples (1 μl) of eluate from the inhibitor affinity matrix were immunoblotted with either rabbit anti-rat CA VI serum or anti-rat CA VI serum absorbed with purified rat CA VI or normal serum.
Results
Expression of CA VI in Trachea and Bronchi
The serous cells present in the tracheal and bronchial epithelium and in the submucosal tracheobronchial glands stained strongly for CA VI (Figure 2A). The staining was located in the apical secretory granules (Figures 2A and 2C). The staining of the serous acinar cells in the tracheobroncial glands appeared as early as the first signs of the glands were detected (on postnatal day 20), suggesting that the secretion of CA VI starts concomitantly with the development of the glands. The stained serous cells of the tracheal epithelium were not uniformly distributed in the epithelium but were most abundant in the vicinity of the orifice of the excretory ducts of the submucosal glands (Figures 2A and 2C). The duct epithelium and the glandular content also stained for CA VI (Figures 2A and 2A inset). Interestingly, the ciliated cells that were abundant in the bronchial epithelium showed distinct positive staining limited exclusively to the base of the cilia (Figure 2E).
Expression of CA VI in the Lung
The serous cells occurring in the surface epithelium and submucosal glands of the trachea and bronchi are not present in the bronchiolar epithelium, which consist of ciliated epithelial cells, goblet cells, and non-ciliated bronchiolar epithelial cells termed Clara cells (Gail and Lenfant 1983; Basbaum et al. 1990). In lung sections, the bronchiolar Clara cells stained strongly for CA VI from the first postnatal day onwards (Figure 2D). The staining transiently decreased in 5- and 10-day-old rats but was again visible in 20- and 30-day-old rats. Moreover, the most strongly stained cells were found in the mucosal folds of the peripheral bronchiolar branches (Figure 2F).
Controls
No difference was detected between the sialoadenectomized and the normal rats in the distribution of staining for CA VI in the trachea or lung (results not shown). Polyclonal rabbit antisera to human salivary α-amylase and rat CA II used as positive controls (Spicer et al. 1982) yielded distinct staining in the tracheobronchial glands (results not shown). CA II staining was diffusely distributed in the cytoplasm of all glandular cells, whereas salivary α-amylase was, similarly to CA VI, located exclusively in the apical secretory granules of the serous acinar cells (results not shown). Sections stained with anti-rat CA VI serum absorbed with purified rat CA VI and normal rabbit serum did not show positive staining, confirming that staining is CA VI-specific (Figures 2B and 2G).
Discussion
The present findings demonstrated that the carbonic anhydrase isoenzyme VI (CA VI) is secreted not only by the salivary, lacrimal, and mammary glands (Parkkila et al. 1990; Ogawa et al. 1995,1998,2002; Karhumaa et al. 2001; Leinonen et al. 2001) but also by the serous acinar cells present in the tracheobronchial epithelium and glands. Moreover, the Clara cells in the bronchiolar epithelium expressed CA VI. These results are in accordance with earlier findings demonstrating that the tracheobronchial glands and mucosal epithelial cells in guinea pig display CA activity (Okamura et al. 1996). The expression of CA VI starts very early during development, being already visible in bronchiolar Clara cells at birth and in tracheobronchiolar glands concomitantly with their emergence (at postnatal day 20). This early onset of CA VI secretion points to its essential physiological role in the maturation and functions of the respiratory tract. The respiratory tract mucosa is exposed from birth to various chemical, physical, and microbial stresses. Therefore, efficient defense mechanisms and strictly regulated renewal systems in the mucosa and particularly in the lining epithelium are required. The present results suggest that CA VI delivered to the mucosal surface forms an essential factor in this defense system and in the renewal of the lining epithelium. Secreted CA VI may form, together with CA II, a cytosolic high-activity isoenzyme, mostly considered a bicarbonate producer, a mutually complementary system for the removal of acidity from mucosal surfaces by accelerating the reaction of acid and bicarbonate into water and CO2. The recent findings by Nakayama et al. (2002) demonstrated that acidity reduces bactericidal activity in the surface liquid of human airway epithelia and increases the susceptibility of these epithelia to bacterial infection. Accordingly, CA VI together with CA II -generated bicarbonate may be implicated in increasing the bactericidal activity of the surface liquid.
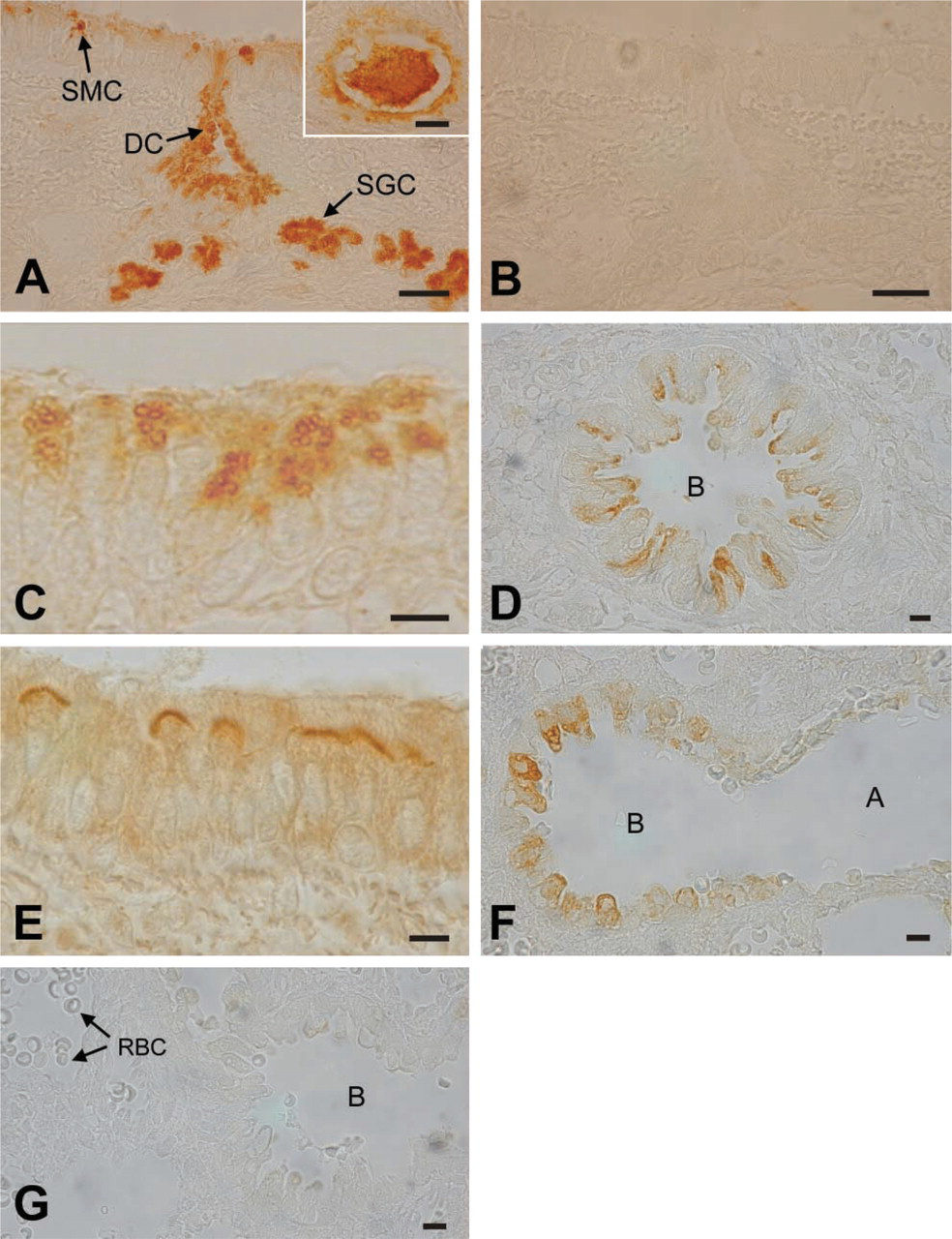
Immunohistochemical localization of CA VI in sections of rat trachea and lung. (
Recent studies have provided evidence that CA VI is a multifunctional protein possessing growth factor activity in addition to carbonic anhydrase activity (Henkin et al. 1999a,b; Karhumaa et al. 2001). Gustin, a salivary secretory protein identical to CA VI (Thatcher et al. 1998), has been demonstrated to protect the taste receptor cells from apoptosis (Henkin et al. 1999a,b). The exact mechanisms, however, remain unidentified. An attractive possibility is that, as in the taste buds, CA VI could also promote the maintenance of the lining epithelium in the respiratory tract. The mechanism may be related to pH neutralization or growth factor function via a specific receptor protein (Hooper et al. 1995). It is also possible that CA VI is implicated in the acidification of the cytoplasm which, in turn, triggers apoptosis, one of the responses of the respiratory tract epithelium to an injury, via a pH-dependent pro-apoptotic regulator called Bax (Basbaum et al. 1990; Antonsson et al. 1997; Sok et al. 1999). The association of CA VI with the brush border of the ciliated epithelial cells in the trachea and bronchi is another interesting finding. Acidic pH has been shown to lower the beat frequency of bronchial cilia (Clary-Meinesz et al. 1998). Our results suggest that CA VI binds from the airway surface liquid to cilia and may be implicated locally in the regulation of their motility via pH neutralization. This could be one mechanism by which CA VI is linked to the protection of the respiratory tract epithelium from microbial infections because cilia motility reduces susceptibility of the airway epithelia to bacterial infection (Afzelius 1976; Whitelaw et al. 1981).
Our results here demonstrate that CA VI is secreted by the seromucous tracheobronchial glands, the serous epithelial cells of the trachea and bronchi, and by the Clara cells of the bronchioli. The presence of CA VI in the airway surface liquid is consistent with the hypothesis that CA VI has a mucosa-protective role not only in the gastrointestinal tract but also in the respiratory tract, where CA VI may act as a pH neutralizer and/or growth factor.