
Abstract
MN/CA IX is a recently discovered member of the carbonic anhydrase (CA) gene family that has been identified in the plasma membranes of certain tumor and epithelial cells and found to promote cell proliferation when transfected into NIH3T3 cells. This study presents localization of MN/CA IX in human gut and compares its distribution to those of CA I, II, and IV, which are known to be expressed in the intestinal epithelium. The specificity of the monoclonal antibody for MN/CA IX was confirmed by Western blots and immunostaining of COS-7 cells transfected with MN/CA IX cDNA. Immunohistochemical stainings of human gut revealed prominent polarized staining for MN/CA IX in the basolateral surfaces of the enterocytes of duodenum and jejunum, the reaction being most intense in the crypts. A moderate reaction was also seen in the crypts of ileal mucosa, whereas the staining became generally weaker in the large intestine. The results indicate isozyme-specific regulation of MN/CA IX expression along the cranial–caudal axis of the human gut and place the protein at the sites of rapid cell proliferation. The unique localization of MN/CA IX on the basolateral surfaces of proliferating crypt enterocytes suggests that it might serve as a ligand or a receptor for another protein that regulates intercellular communication or cell proliferation. Furthermore, MN/CA IX has a completely conserved active site domain of CAs suggesting that it could also participate in carbon dioxide/bicarbonate homeostasis.
T
MN/CA IX was first recognized as the novel tumor-associated antigen, MN, a transmembrane protein that was detected in several human carcinomas and in normal gastric mucosa (Liao et al. 1994; Pastorek et al. 1994). On Western blots, the protein appeared as two bands of 54 and 58 kD (Pastorek et al. 1994). Its expression was found to be regulated by cell density in HeLa cells and to correlate with tumorigenicity in HeLa cell/fibroblast cell hybrids (Závada et al. 1993). Moreover, expression of MN/CA IX in NIH3T3 cells promoted cell proliferation (Pastorek et al. 1994). When the full-length cDNA for MN protein was cloned, it was found to contain a central part with sequence homology to the CAs (Pastorek et al. 1994; Opavský et al. 1996), on which basis the MN protein was named CA IX (Hewett–Emmett and Tashian 1996). A recent study of rat and human tissues revealed that MN/CA IX is more widely expressed than was initially recognized (Pastoreková et al. 1997). Although the strongest signals were detected in the gastric and gallbladder mucosas, the stainings also demonstrated its presence in the intestine.
The major aim of this study was to immunolocalize MN/CA IX in different segments of the human intestine and to determine whether its membrane localization suggests a relationship to sites of cell proliferation. The availability of reagents for the other isozymes enabled us to contrast the localization of MN/CA IX with those of other CAs that have been reported to be present in gut but never completely evaluated in parallel.
Materials and Methods
Antibodies Against CA Isozymes
Murine monoclonal antibody (MAb) M75 recognizing the N-terminal domain of MN/CA IX protein has been previously described by Pastoreková et al. (1992). Polyclonal rabbit antisera to human CA I, II, IV, and VI have been produced and characterized previously (Parkkila et al. 1991, 1993a,1996). The anti-CA IV antibodies were raised against the secretory form of human recombinant CA IV (Okuyama et al. 1995). The crossreactivities of these antibodies have been tested in dot and Western blottings and in time-resolved immunofluorometric assays, in which they have shown high isozyme specificities (Parkkila et al.1991,1993a,b,c,1994a, 1996). Because salivary CA VI is not expressed in the gut epithelium (Parkkila et al. 1994b), the anti-CA VI serum was used for the control stainings.
Transfection of COS-7 Cells
The cDNA of human MN/CA IX (Opavský et al. 1996) was subcloned into the pCAGGS vector originally described by Miyazaki et al. (1989) and used by Yoshida et al. (1991). COS-7 cells in 60-mm dishes and in chamber slides for microscopy were transfected with 10 μg DNA/dish and 2 μg DNA/200 μl, respectively, using the DEAE-dextran procedure (Lopata et al. 1984) and chloroquine treatment 12 hr after transfection (Luthman and Magnusson 1983). The transfected cells were harvested by scraping into PBS 72 hr after transfection and were disrupted and lysed by sonication in 25 mM Tris-SO4 buffer, pH 7.2, containing 0.05% Triton X-100, 1 mM PMSF, 1 mM benzamidine, and 1 mM o-phenanthroline.
SDS-PAGE and Western Blotting
All the reagents for SDS-PAGE were from Bio-Rad Laboratories (Richmond, CA) or Sigma (St Louis, MO). The electrophoreses were performed in a Mini-Protean electrophoresis unit (Bio-Rad Laboratories) under reducing conditions according to Laemmli (1970), using a 9% acrylamide separating gel and a 4% acrylamide stacking gel. An aliquot of 10 or 20 μg of COS-7 cell homogenate or 0.5 μg of purified CA I, II, IV, or VI was applied to each lane. The proteins were transferred electrophoretically from the gel to a PVDF membrane (Millipore; Bedford, MA) in a Novex Blot Module (Novex; San Diego, CA). After the transblotting, the sample lanes were first incubated with TBST buffer (10 mM Tris-HCl, pH 7.5, 150 mM NaCl, 0.05% Tween-20) containing 10% cow colostral whey for 30 min at room temperature (RT) and then with M75 hybridoma medium (anti-MN/CA IX antibody) diluted 1:100 in TBST buffer for 15 hr at 4C. The sheets were washed five times for 5 min with TBST buffer and incubated for 2 hr at RT with peroxidase-conjugated anti-mouse IgG (Sigma) diluted 1:1000 in TBST buffer. After washing four times for 5 min in TBST buffer, the polypeptides were visualized by a chemiluminescence substrate.
Immunocytochemistry
The COS-7 cells transfected with MN/CA IX cDNA were fixed with 4% neutral-buffered formaldehyde for 15 min. Then they were rinsed with PBS and subjected to immunofluorescence staining that consisted of the following steps: (a) pretreatment of the cells with 1:10 diluted cow colostral whey in PBS for 40 min and rinsing in PBS; (b) incubation for 1 hr in the M75 hybridoma medium diluted 1:10 in 0.1% bovine serum albumin in PBS (BSA–PBS); and (c) incubation with 1:100 diluted fluorescein-conjugated goat anti-mouse IgG antibodies (Sigma) in 0.1% BSA-PBS. The cells were washed three times for 10 min after the incubation steps. All incubations and washings were done in the presence of 0.05% saponin. The cells were viewed with a conventional Zeiss Axioplan epifluorescence microscope (Oberkochen, Germany) and a confocal laser scanning microscope (Zeiss Axiovert 135 microscope combined with an LSM 410 CLSM system). The specimens were excited with a laser beam at a wavelength of 488 nm and the emission light was focused through a pinhole aperture. The full field was scanned in square image formats of 512 X 512 pixels and built-in software was used to reconstruct the images obtained from the confocal sections.
Samples of human duodenum (n = 2), jejunum (n = 2), ileum (n = 3), cecum (n = 3), ascending colon (n = 3), transverse colon (n = 3), descending colon (n = 3), sigmoid colon (n = 3), and rectum (n = 2) were obtained together with routine histopathological specimens taken during surgical operations for gastric carcinoma (n = 2), rectal carcinoma (n = 1), colon polyposis (n = 2), or colon inertia (n = 1). The procedures had the approval of the ethics committee of Oulu University Hospital and the research was carried out according to the provisions of the Declaration of Helsinki. Each tissue sample was divided into several small pieces about 5 mm thick. The specimens were fixed in Carnoy's fluid (absolute ethanol + chloroform + glacial acetic acid 6:3:1) for 6 hr or in 4% paraformaldehyde in PBS for 18 hr. All fixations were carried out at 4C. The samples were then dehydrated, embedded in paraffin in a vacuum oven at 58C, and sections of 5 μm were placed on gelatin-coated microscope slides. Carnoy's fluid preserved best the antigenicity of CA I, II, and MN/CA IX, whereas the staining reaction for CA IV was best preserved after paraformaldehyde fixation, which also was found previously to be the most suitable fixative for CA VI (Parkkila et al. 1990).
The CA isozymes were immunostained by the biotin-streptavidin complex method, employing the following steps: (a) pretreatment of the sections with undiluted cow colostral whey for 40 min and rinsing in PBS; (b) incubation for 1 hr with the primary antiserum diluted 1:100 (CA I, II, IV, and VI) or the M75 hybridoma medium diluted 1:10 in 1% BSA-PBS; (c) treatment with cow colostral whey for 40 min and rinsing in PBS; (d) incubation for 1 hr with biotinylated swine anti-rabbit IgG (Dakopatts; Glostrup, Denmark) or rabbit anti-mouse IgG (Dakopatts) diluted 1:300 in 1% BSA–PBS; (e) treatment with cow colostral whey for 5 min and rinsing in PBS; (f) incubation for 30 min with peroxidase-conjugated streptavidin (Dakopatts) diluted 1:500 in PBS; and (g) incubation for 2 min in DAB solution containing 9 mg 3,3'-diaminobenzidine tetrahydrochloride (Fluka; Buchs, Switzerland) in 15 ml PBS + 10 μl 30% H2O2. The sections were washed three times for 10 min in PBS after incubation Steps b, d, and f. All the incubations and washings were carried out at RT, and the sections were finally mounted in Permount (Fisher Scientific; Fair Lawn, NJ). The stained sections were examined and photographed with a Leitz Aristoplan microscope (Wetzlar, Germany).
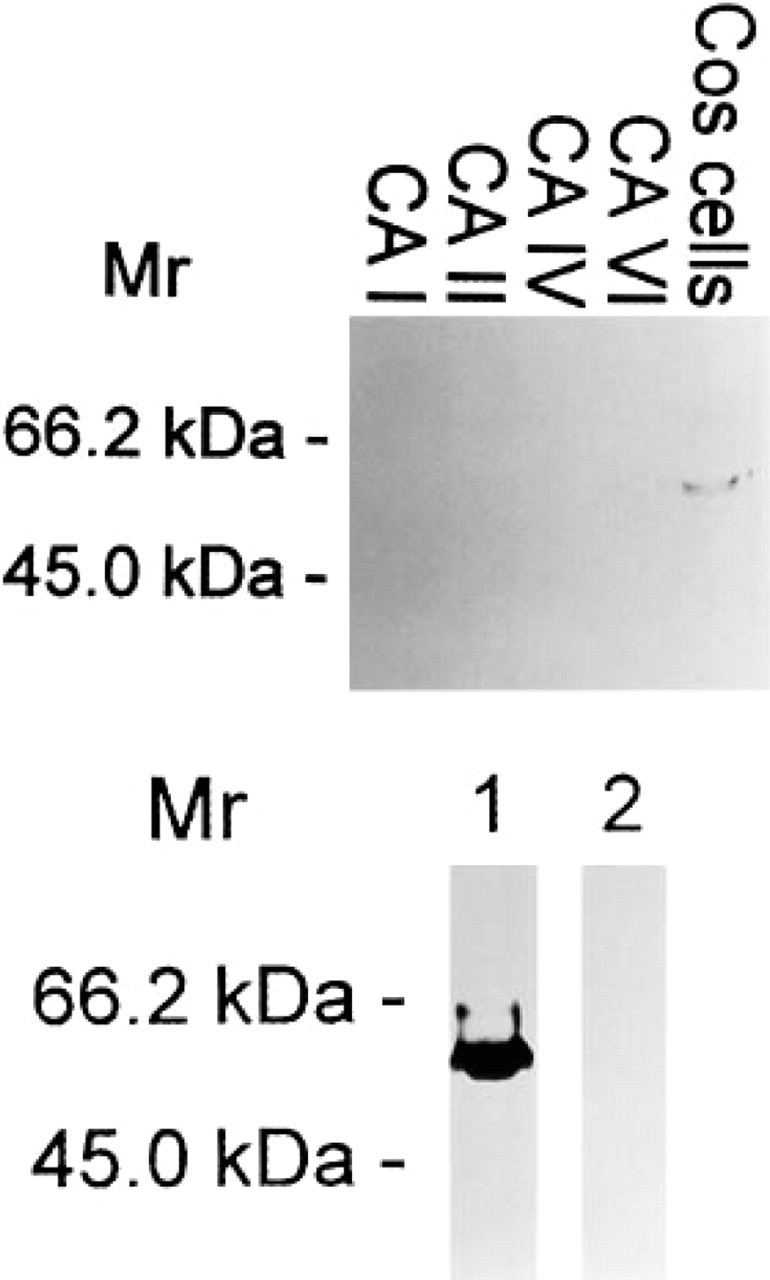
Specificity of the monoclonal anti-MN/CA IX antibody tested by Western blotting. Aliquots of 10 (upper panel) and 20 μg (lower panel) of COS-7 cell homogenate or 0.5 μg of purified CA I, II, IV, or VI was applied to each lane. In the Western blot of COS-7 cells transfected with MN/CA IX cDNA (upper panel, COS cells; lower panel, Lane 1), the antibody identified a major polypeptide band of 58 kD. No signal for MN/CA IX was seen in the proteins of non-transfected COS-7 cells (lower panel, Lane 2).
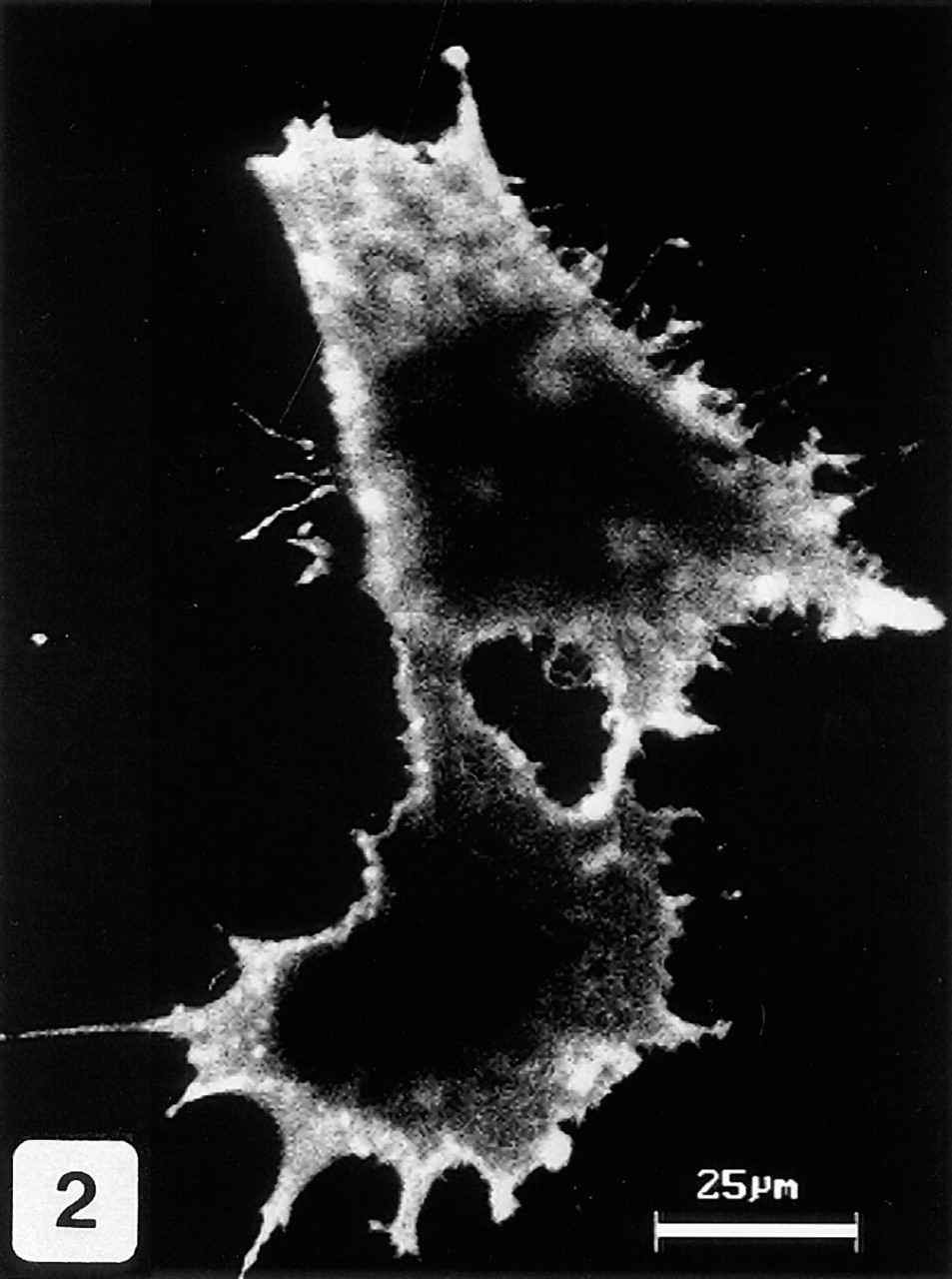
Immunocytochemical staining of permeabilized COS-7 cells transfected with MN/CA IX cDNA. A strong signal for MN/CA IX protein is mainly associated with the plasma membrane.
Results
Specificity of Anti-MN/CA IX Antibody
The monoclonal anti-MN/CA IX antibody, which identified a strong 58-kD and a weak 54-kD polypeptide band on Western blots of COS-7 cells transfected with MN/CA IX cDNA, showed no crossreactivity with other CAs (CA I, II, and IV) which are known to be expressed in the human gut (Parkkila and Parkkila 1996), nor with CA VI, which is not expressed in the gut but has the greatest homology to MN/CA IX (Opavský et al. 1996) (Figure 1). The molecular weights of MN/CA IX polypeptide bands were the same as reported earlier (Pastoreková et al. 1997). The lower panel of Figure 1 presents a Western blot, showing a strong immunoreaction at 58 kD in transfected COS-7 cells that is completely absent in non-transfected cells. Figure 2 shows a confocal laser scanning microscopic image of immunofluorescence staining for MN/CA IX in COS-7 cells transfected with MN/CA IX cDNA. The permeabilized cells showed a strong plasma membrane-associated signal and a weaker intracellular signal. Immunocytochemistry did not show any immunoreactivity for MN/CA IX in the non-transfected COS-7 cells (data not shown).
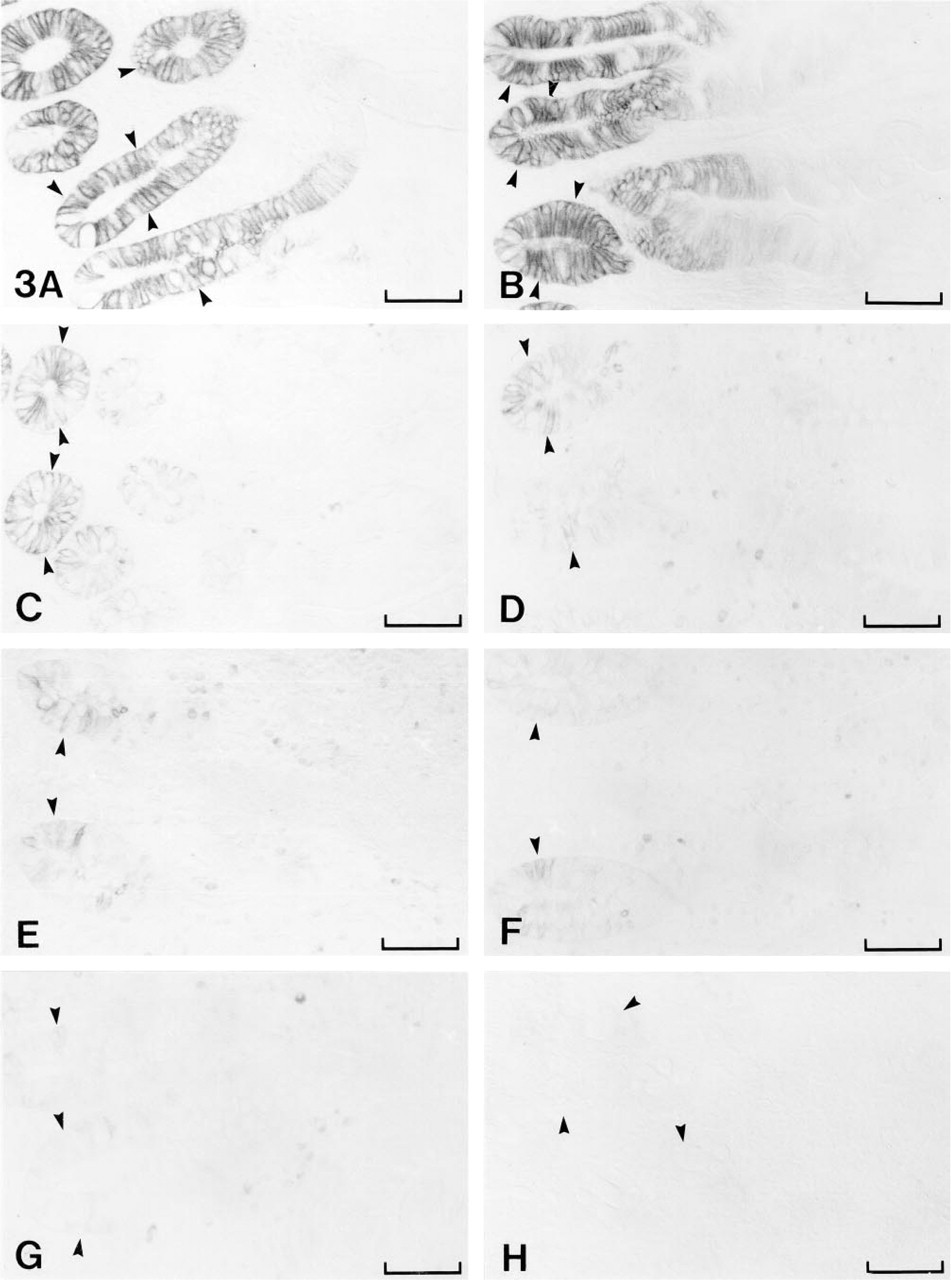
Immunohistochemical staining of MN/CA IX in different segments of the human gut. The positive immunoreaction is most intense in the crypt enterocytes of duodenum (
Distribution of MN/CA IX, CA I, CA II, and CA IV in human guta
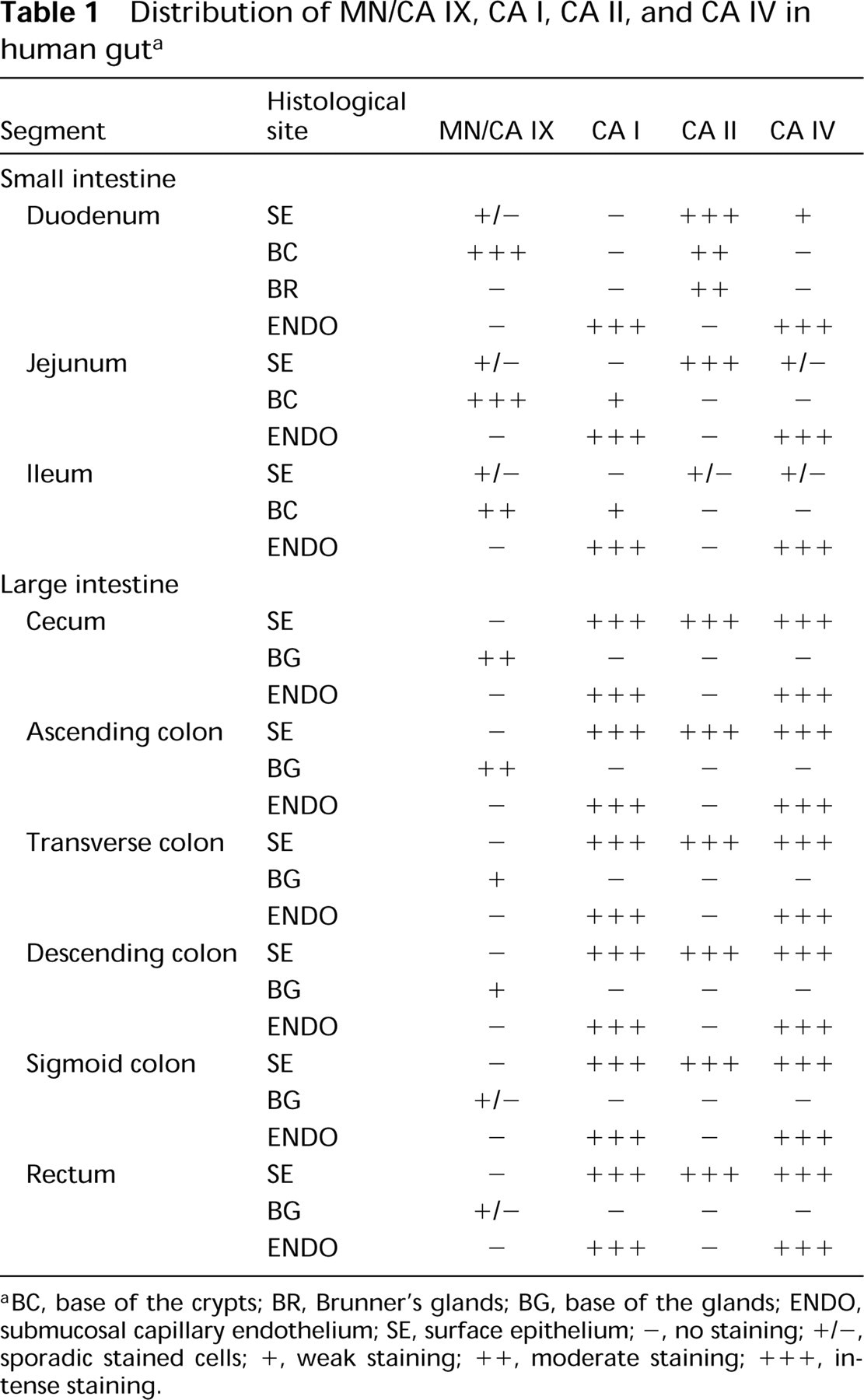
BC, base of the crypts; BR, Brunner's glands; BG, base of the glands; ENDO, submucosal capillary endothelium; SE, surface epithelium; -, no staining; +/-, sporadic stained cells; +, weak staining; + +, moderate staining; + + +, intense staining.
Immunohistochemistry of MN/CA IX in the Gut
MN/CA IX showed a unique staining pattern in human gut. First, it was highly expressed in duodenum and jejunum (as in stomach) and was distinctly less expressed in more distal segments of the gut. Second, the intestinal staining for MN/CA IX was most intense in the crypts. Third, the positive signal for MN/CA IX was confined to the basolateral plasma membranes in all stained enterocytes.
Table 1 and Figure 3 show the immunohistochemical localization of MN/CA IX in different segments of the gut. Figures 3A (duodenum) and 3B (jejunum) show the intense staining in the crypt enterocytes, whereas the epithelial cells in upper portions and tips of the villi showed only a weak immunoreaction. MN/ CA IX was also present in enterocytes of human ileum (Figure 3C), although the positive reaction was much weaker than in duodenum and jejunum. In the large intestine, the basolateral surfaces of the crypt enterocytes showed positive staining for MN/CA IX, with a gradual change in staining intensity in a distal direction. The reaction was still moderate in cecum (Figure 3D) and ascending colon (Figure 3E), whereas the transverse (Figure 3F) and descending parts (Figure 3G) showed considerably weaker signals. Only sporadic enterocytes in sigmoid colon and rectum exhibited weak basolateral staining (Figure 3H).
Immunohistochemistry of CA I, II, and IV in the Gut
CA I showed three distinct sites of expression in the gut. In jejunum and ileum, weak but definite cytoplasmic staining for CA I was localized to the epithelial cells of crypts of Lieberkühn, whereas no staining was seen in the villi (Figure 4A). In the large intestine, prominent reactions for CA I were seen in the non-goblet epithelial cells of the surface epithelial cuff, but the staining gradually decreased towards the base of the gland epithelium (Figure 4D). The endothelial cells of the subepithelial capillaries were the third major site of CA I expression (Figures 4A, and 4D). This is consistent with previous reports showing that CA I is expressed in the capillary endothelium of gallbladder (Juvonen et al. 1994), brain (Parkkila et al. 1995), and placenta (Mühlhauser et al. 1994).
In the small intestine, CA II was localized in the villi of duodenum and jejunum and the staining decreased towards the base of the crypts (Figure 4B). In the ileum, only sporadic non-goblet epithelial cells showed positive signal for CA II (data not shown), whereas the staining again became intense in the non-goblet surface epithelial cells of the large intestine (Figure 4E).
The membrane-associated CA IV showed intense immunoreaction in two different cell types: the endothelial cells of the subepithelial capillaries (Figure 4C) and the epithelial cells of the large intestine (Figure 4F). The epithelial brush border membrane showed intense immunoreaction in all segments of the large intestine. The reaction was strongest in the luminal part of the mucosa.
Discussion
MN/CA IX is a novel member of the α-CA gene family and is the only known integral membrane protein with CA activity (unpublished observations). Another membrane-associated CA isozyme, CA IV, is not considered an integral membrane protein because the mature protein is anchored to the plasma membrane through a glycosyl phosphatidylinositol linkage (Sly and Hu 1995). Like other active CAs, MN/CA IX is also inhibitable with sulfonamides, which further supports its role as an active CA (unpublished observations). According to the amino acid sequence deduced from the cDNA of the MN/CA IX, four distinct regions can be distinguished: an N-terminal signal peptide, an extracellular part containing the carbonic anhydrase- and proteoglycan-related regions, a transmembrane region, and a cytoplasmic tail at the C-terminal part (Opavský et al. 1996). The major aim of the present study was to investigate the regional, cellular, and subcellular localization of MN/CA IX in the human gut. The availability of other purified CAs expressed in the gut and antibody reagents to each of them enabled us to verify the specificity of the anti-MN/CA IX antibody and to compare its sites of expression to those of other CAs in parallel tissue sections. Another opportunity to verify the specificity of the MN/CA IX immunostainings was provided by the expression of MN/CA IX in COS-7 cells, which produced immunoreactive protein that was localized to the plasma membrane. As reported for Western blots of human stomach (Pastoreková et al. 1997), the two polypeptides identified by the monoclonal MN/CA IX antibody on Western blots of transfected COS-7 cells were the 58-kD form and the 54-kD form, which was here barely apparent. The anti-MN/CA IX antibody did not crossreact with purified CA I, II, IV, and VI, all of which are expressed in various parts of the alimentary tract (Parkkila and Parkkila 1996), providing further evidence that the immunostaining for MN/CA IX in the gut is isozyme-specific.
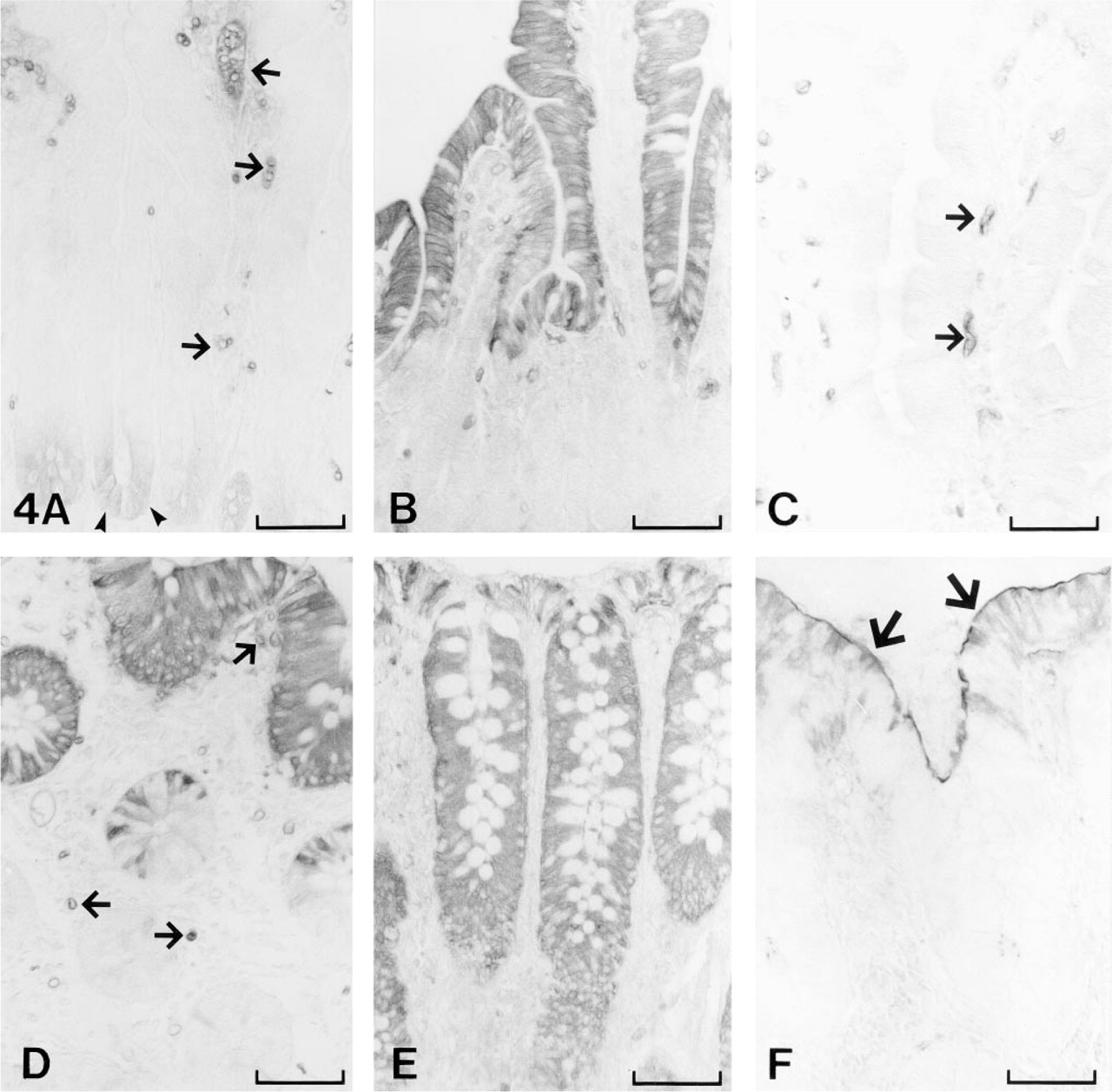
Immunohistochemical demonstration of CA I (
The present studies demonstrated that the distribution of MN/CA IX in the gut has unique features. First, its subcellular localization is restricted to the basolateral surfaces of epithelial cells. Second, its cellular distribution is restricted to the crypt enterocytes, which is not seen with any other CA. Third, its regional expression is distinctive compared to other CAs, being most intense in duodenum and jejunum, and decreasing distally from moderate expression in crypts of ileum, cecum, and ascending colon to only weak and sporadic expression in distal large intestine.
Restriction of MN/CA IX to the epithelial cells with the greatest proliferative capacity is consistent with its proposed role in cell proliferation (Pastorek et al. 1994). Stem cells located in the crypts are a source for continuous and rapid renewal of the epithelium (Gordon 1989). Earlier studies in mice have shown that each small intestinal crypt produces an average of 13–16 new cells/hr (Gordon et al. 1992). Cell differentiation and cell migration are coupled events in the gut. The epithelial cells differentiate and mature during migration from the crypt to the apex of the villus or surface epithelial cuff (Lipkin 1985; Gordon 1989; Gordon et al. 1992). This process is completed in a few days, which makes the mammalian intestinal epithelium a unique model system for studying cell differentiation. Because MN/CA IX is much more abundant in the proliferating crypt epithelium than in the upper part of the mucosa, it may serve as a useful marker to study the proliferation and differentiation of the intestinal epithelium. Further support for its role as a marker for cell proliferation is obtained from studies showing that the rate of cell proliferation is highest in the proximal segments of the intestine (Lipkin 1985; Gordon 1989), which was also the site of the highest MN/CA IX expression.
As a CA-homologous transmembrane protein, MN/ CA IX has some structural similarity with recently described receptor-type protein tyrosine phosphatases (RPTPs) β and γ, which are members of a distinct group of phosphatases that have CA-homologous regions in their extracellular domains (Krueger and Saito 1992; Barnea et al. 1993; Levy et al. 1993). Both transmembrane and secretory forms of RPTPβ have been found to be identical with a chondroitin sulfate proteoglycan called phosphacan (Barnea et al. 1994a,b; Maurel et al. 1994; Shitara et al. 1994). The CA-like domains of the RPTPβ and -γ showed about 30–50% amino acid identity with CAs (Krueger and Saito 1992; Barnea et al. 1993; Levy et al. 1993). Unlike MN/CA IX, RPTPβ and -γ have only one of the three conserved histidine residues required to bind to zinc ion in catalytically active CAs. Therefore, it is unlikely that they have any CA activity. Recent studies have indicated that the CA-like domain of RPTPβ binds to contactin, a neuronal cell recognition molecule (Peles et al. 1995), and to tenascin, an extracellular matrix protein (Milev et al. 1995). The CA-like domain induced cell adhesion and neurite growth of primary tectal neurons and differentiation of neuroblastoma cells (Peles et al. 1995). Because the expression of MN/CA IX is regulated by cell density (Závada et al. 1993) and is greatest in basolateral surfaces of proliferating cryptal enterocytes, MN/CA IX might also serve as a ligand or a receptor for another protein that regulates intercellular communication or cell proliferation. However, unlike the RPTPβ and -γ, MN/CA IX has a completely conserved active site domain that suggests that it could also participate in carbon dioxide/ bicarbonate homeostasis. It will be of interest to search for ligands and/or receptors for MN/CA IX and to explore its functional role in the regulation of carbon dioxide homeostasis and in the growth and development of intestinal cells.
Footnotes
Acknowledgements
Supported by grants DK40163 and GM34182 from the U.S. Public Health Service to WSS, from the Sigrid Juselius Foundation to SP, from the Maud Kuistila Foundation and Academy of Finland to AKP, and from Chiron Diagnostics Corporation and the Slovak Scientific Grant Agency to SP and JP.