
Abstract
Here we examined the changes in NADPH diaphorase (NADPHd) and inducible nitric oxide synthetase (iNOS) positivity in the medulla of the mouse thymus in response to treatment with the superantigen, Staphylococcal enterotoxin B (SEB). A few NADPHd+ and iNOS+ cells scattered in the medulla were detected in the thymi of naive mice. SEB induced the appearance of a large number of NADPHd+- and iNOS-immunoreactive cells in the thymic medulla. In the thymus of iNOS-deficient mice, a total absence of these NADPHd+ and iNOS+ medullary cells was found both under basal conditions and after SEB stimulation. With the NADPHd reaction, only endothelial staining was detected in the thymi of iNOS-deficient mice. Our data indicate that NADPHd+ cells in the thymic medulla express iNOS and that SEB induces iNOS expression in the mouse thymus.
N
Nitric oxide synthases have been shown to have NADPH diaphorase (NADPHd) activity, as evidenced by co-localization and co-precipitation of NADPHd and NOS activity (Snyder 1992). Furthermore, cells transfected with NOS cDNA also assume NADPH diaphorase activity (Dawson et al. 1991; Snyder 1992). The NADPHd method has been widely used to detect bNOS in the nervous system (Dawson et al. 1991; Murphy 1994). However, it has only sporadically been applied to reveal iNOS expression in peripheral tissues (Kobzik et al. 1993; Kikuchi et al. 1996; Bedard et al. 1997). Although NADPHd+ cells have been found in the chicken and rat thymic medulla (Gulati et al. 1993; Downing 1994), their NOS positivity has not yet been confirmed. Furthermore, immunohistochemistry revealed iNOS expression in the thymus of marmosets, whereas NADPHd did not (Grozdanovic et al. 1995). Here we investigated whether NADPHd histochemistry detects NOS+ cells in the thymis of naive and Staphylococcal enterotoxin B (SEB)-treated mice. SEB is a superantigen that causes the MHC-dependent activation of T-cells bearing the Vβ 8 motif on the T-cell receptor (Marrack and Kappler 1990). This T-cell activation is accompanied by the release of a variety of pro-inflammatory cytokines (IFN, TNF) and NO (Florquin et al. 1994). The regulatory role of NO in SEB-induced clonal proliferation and cell death has been proposed (Bras et al. 1997; Kuschnaroff et al. 1997). The SEB-induced changes in intrathymic iNOS expression have not yet been investigated. To affirm specificity of the NADPHd reaction, we compared the responses in wild-type mice with the responses in iNOS-deficient mice.
Materials and Methods
Materials
Anti-mouse iNOS antibody was purchased from Upstate Biotechnology (Lake Placid, NY). The Vector Elite kit was obtained from Vector Laboratories (Burlingame, CA). All other reagents were from Sigma (St Louis, MO).
Treatment of Animals
Animal experiments conform with the “Guide for the Care and Use of Laboratory Animals” published by the US National Institute of Health and the treatment protocol was approved by the Institutional Animal Care and Use Committee. Male C57BL10 mice and Wistar rats were purchased from Jackson (Bar Harbor, ME) and Charles River Laboratories (Wilmington, MA), respectively. Male iNOS-deficient mice [on C57BL/10 background (Laubach et al. 1995), a generous gift of Dr. V. Laubach] were bred at the animal facility of our hospital. Mice were either left untreated or treated with 100 μg SEB
NADPH Diaphorase Histochemistry
The method was carried out essentially as described previously (Downing 1994). Sections were defrosted and washed in assay buffer (0.1 M Tris, pH 7.4, 1 mM manganese chloride, 0.5 % Triton X-100) for 30 min at room temperature. Sections were then incubated in a solution containing 30 mM malic acid, 1 mM NADP, 0.2 mM nitroblue tetrazolium chloride in assay buffer for 60 min at 37C. Incubations without NADP were carried out as controls. Slides were then briefly rinsed with PBS, dehydrated, and mounted in Permount medium. Photomicrographs were taken with an Olympus BX-40 microscope.
Immunohistochemistry
Cryostat sections (10 μm) were treated with 0.3% hydrogen peroxide for 15 min to block endogenous peroxidase activity and then rinsed briefly in PBS. Nonspecific binding was blocked by incubating the slides for 1 hr in 2% goat serum (in PBS). To detect iNOS, rabbit polyclonal anti-iNOS antibody was applied at a dilution of 1:500 at 4C overnight. (Control sections were incubated in PBS without the primary antibody.) After extensive washing (five times for 5 min) with PBS, immunoreactivity was detected with a biotinylated goat anti-rabbit secondary antibody and the avidin-biotin-peroxidase complex (ABC), both supplied in the Vector Elite kit. Color was developed using Ni-DAB substrate (95 mg diaminobenzidine, 1.6 g NaCl, 2 g nickel sulfate in 200 ml 0.1 M acetate buffer). Sections were counterstained with nuclear fast red for 2 min and mounted in Permount medium.
Results
NADPHd Reveals iNOS-expressing Cells in the Medulla of Naive Mouse Thymus
The inducible isoform of nitric oxide synthetase (iNOS) is usually expressed in response to cytokine and/or bacterial stimuli. The thymus appears to be a unique organ in this respect, because a number of NADPHd+ medullary cells have been described in the resting chicken and rat thymus (Gulati et al. 1993; Downing 1994). Their being iNOS-positive, however, has not been confirmed. Here we report the existence of a distinct population of NADPHd+ cells in the medulla of naive mouse thymi (Figure 1A). Their morphology is somewhat different from what has previously been found in the rat thymus (Downing 1994). Rat thymic NADPHd+ cells are also located in the medulla, with the highest density at the corticomedullary junction, and they have long branching processes. On the basis of their location and morphology, they were assumed to be dendritic cells (Downing 1994). Mouse thymic NADPHd+ cells are similarly distributed, but they appear to be less abundant than their rat counterparts and their processes are shorter, giving them a star-like morphology. We have observed iNOS+ immunoperoxidase staining in the thymic medulla of naive mice, with a pattern similar to the pattern of cells expressing NADPHd (Figure 1B)
SEB Induces iNOS Expression in Thymic Medullary Cells
NADPHd reaction after SEB administration revealed a massive increase in the number of positive cells in the mouse thymus (Figures 1C and 1E) compared to the naive thymus. SEB induced a marked increase in the amount of immunohistochemical staining for iNOS, with a pattern identical to that of NADPHd (Figures 1D and 1F). In the thymi of iNOS-deficient animals, neither NADPHd (Figure 1G) nor iNOS staining (Figure 1H) could be detected after SEB challenge (Figures 1G and 1H). The lack of these NADPHd+ medullary cells in the thymocytes of iNOS knock-out mice (Figure 1G) strongly indicates that the NADPHd+ staining was indeed related to iNOS. We have observed some endothelial NADPHd staining in the iNOS knock-out thymi, which probably corresponds to eNOS expressed constitutively by endothelial cells (Figure 1G).
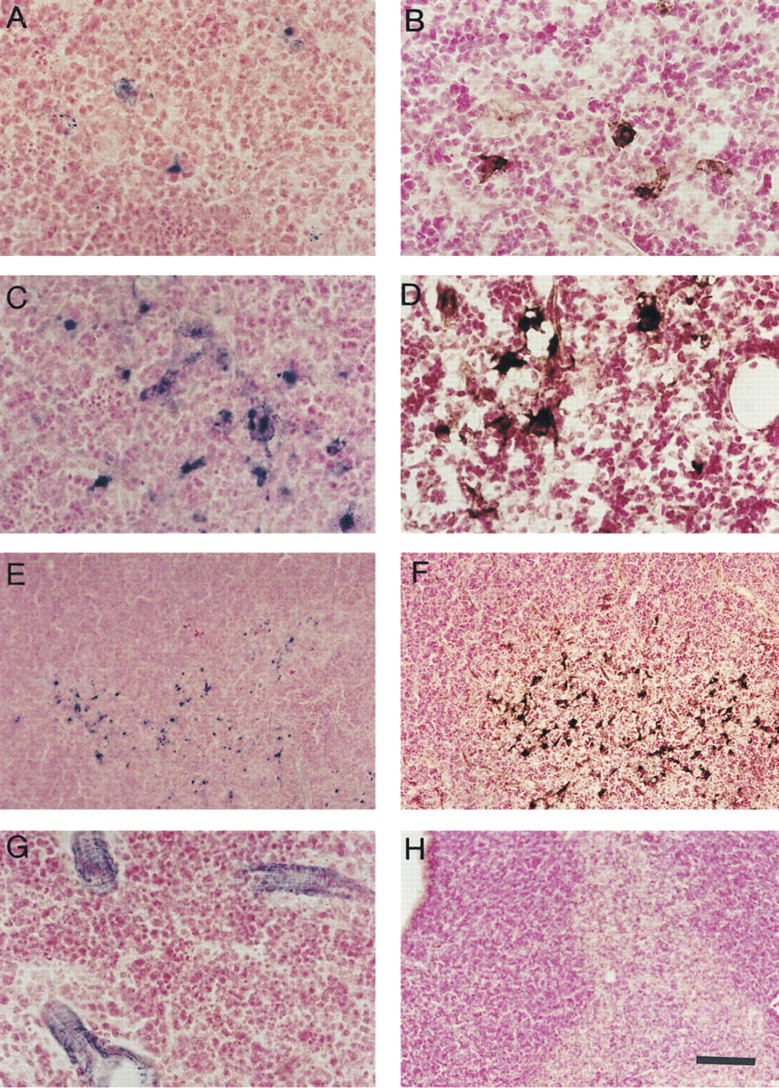
NADPH diaphorase histochemistry reveals iNOS-expressing cells in the thymi of naive and SEB-stimulated mice.
Discussion
The key observations of this report can be summarized as follows. (a) We have provided evidence for NADPHd+ and iNOS+ cells in the naive mouse thymus. (b) We have demonstrated a massive increase in the number of NADPHd+ and iNOS+ cells in response to SEB exposure. (c) We have shown the complete absence of NADPHd+ and iNOS+ thymic medullary cells in the thymi of iNOS knock-out animals. Based on the parallel changes in the amount of NADPHd+ and iNOS+ medullary cells in response to SEB and on the absence of these cells in the thymi of iNOS-deficient mice, we propose that NADPHd staining primarily detects the presence of iNOS in the thymus, both under resting conditions and under conditions of immunostimulation.
We have detected the expression of iNOS in the medulla of the naive mouse thymus. Although this may seem contradictory to the term “inducible” NOS, we hypothesize that, as has been proposed by Tai et al. (1994, 1997) iNOS expression by dendritic cells in the naive thymus is induced by thymocyte-derived cytokines and is a result of a cross-talk between thymocytes and antigen presenting cells during the negative selection process. This may be initiated by the high-affinity binding of the T-cell receptors of potentially autoreactive thymocytes to self antigens presented by dendritic cells, which in turn signals thymocytes to produce interferon-γ. Thymocyte-derived interferon-γ, in turn, activates stromal cells leading to iNOS expression, NO secretion, and NO-mediated apoptosis of thymocytes (Lu et al. 1996). Alternatively, increased iNOS expression may result from the direct effect of SEB on IFNγ prestimulated stromal cells (Fast et al. 1991).
The wide range of physiological and pathophysiological effects of NO and NO-derived reactive species set up a claim for simple and sensitive techniques to detect NOS activity in tissue sections. NADPHd histochemistry, a fast and simple reaction often used by neuroscientists to visualize the expression of bNOS in the nervous system, offered a good alternative to the expensive and time-consuming immunohistochemistry. Although many enzymes have diaphorase activity which may cause specificity problems, pretreatment of tissues with 4% p-formaldehyde has been shown to eliminate non-NOS-derived activity (Matsumoto et al. 1993). The assay procedure of our presently used NADPHd reaction takes 1.5 hr compared to 16 hr required for immunohistochemistry. Moreover, the reaction is highly cost-effective, because the chemicals used are cheap and no antibodies are needed. On the basis of our finding, we propose that the NADPHd reaction offers a simple, fast, and sensitive screening method for detection of changes in the expression of iNOS in thymic sections and possibly also in other peripheral tissues.