
Abstract
Ganglion cells and topographically related nerves in the vallate papilla/von Ebner gland complex were investigated in rat tongue by cytochemical, immunocytochemical, and ultrastructural methods to evaluate the possible presence of different neuronal subpopulations. Immunostaining for neurofilaments and protein gene product 9.5 revealed ganglionic cell bodies and nerve fibers. A large part of the neurons were positive at immunostaining for neuronal nitric oxide synthase (NOS), vesicular acetylcholine transporter (VAChT), or vasoactive intestinal peptide (VIP). A small subset of nerve fibers revealed immunoreactivity for cholecystokinin. Axons traveling under the lingual epithelium were evidenced by their content of calcitonin gene-related peptide (CGRP) or substance P (SP). Cell bodies positive for SP or CGRP were not detected. Using methods of co-localization, three different neuronal classes were detected. The main population was composed of AChE/NADPH-diaphorase (NADPHd)-positive cells. Small groups of acetylcholine esterase (AChE)-positive/NADPHd-negative cells were visible. Isolated neurons were AChE-negative/NADPHd-positive. The results of co-localization experiments for VAChT/NOS were consistent with those obtained by cytochemical co-localization of AChE and NADPHd. Experiments of co-localization for peptidergic and nitrergic structures revealed CGRP- and SP-immunoreactive fibers in the vallate papilla/von Ebner gland ganglion. In conclusion, the results demonstrated in the VP/VEG complex peptidergic, cholinergic, and nitrergic neurons. The presence of different neuronal subclasses suggests that a certain degree of functional specialization may exist.
T
In the past, several authors described the presence of ganglion cells in the connective tissue below the VP of the tongue. Multipolar and bipolar nerve cells under the VP of mammals were first described by Remak (1852), Drasch (1887), and Fusari and Panasci (1889). More recently, Kane and Law (1950) in pig, Gairns and Garven (1952) in rat, hedgehog, and human, and Baradi and Bourne (1959) in rabbit described nerve cells under the VP and foliate papillae. Further data on these cells of different mammals have also been reported by Bradley et al. (1985) and by Fitzgerald and Alexander (1969). The ultrastructure of ganglion cells in the VP of rat and mouse was described by Graziadei and Monti Graziadei (1978). We demonstrated that the neurons are mainly nitrergic elements, as revealed by light microscopy evaluation of neuronal nitric oxide synthase (NOS) immunoreactivity, and NADPH-diaphorase (NADPHd) cytochemical positivity (Sbarbati et al. 1999). The organization of the nitrergic cells suggested that NO has a role in the regulation of the VP/VEG complex mediating interactions between the gustatory epithelium and the secretory cells in the VEG. The nitrergic fibers were located in the chemoreceptor-free mucosa, below the gustatory epithelium, in VEG, and around blood vessels. To date, it is not clear if all the cells of the VP/VEG complex are nitrergic or if different neuronal subpopulations exist. The demonstration of different neuronal types could strengthen the hypothesis that neurons of the VP/VEG complex are not simple parasympathetic relays but might have a possible integrative role. In this work we studied the ganglion cells and topographically related nerves of the VP/VEG complex by cytochemical and immunocytochemical methods. As general nerve markers, we used immunocytochemistry for protein gene product (PGP) 9.5 and neurofilaments (NF200). Nitrergic structures were studied by cytochemistry for NADPHd and immunocytochemistry for NOS. Peptidergic fibers were stained by immunocytochemistry for calcitonin gene-related peptide (CGRP), substance P (SP), cholecystokinin-8 (CCK), and vasoactive intestinal peptide (VIP). Cholinergic structures were evidenced by cytochemistry for acetylcholine esterase (AChE), and using immuno-cytochemistry for vesicular acetylcholine transporter (VAChT), which is an ideal marker for acetylcholine-containing neurons (Arvidsson et al. 1997).
The aim of the work was to correlate the distribution of nitrergic cells in the VP/VEG complex with cholinergic and peptidergic structures, evaluating the possible presence of neuronal subpopulations.
Materials and Methods
Tissue Preparation
Wistar rats (150–200 g; Morini Company, Reggio Emilia, Italy) were anesthetized with ether, sacrificed by dislocation of cervical vertebrae, and perfused via the aorta with 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4). The lingual tissue was postfixed by immersion for 1 hr in the same fixative. The tissue was transferred into a 30% sucrose solution in phosphate buffer. Blocks of lingual tissue containing the VP as well as adjacent epithelium, muscles, and glands were dissected and sectioned on a freezing microtome. Free-floating sections (30–40 μm) were collected in PBS (pH 7.4) and washed in this solution. In other rats, the tongues were removed after sacrifice and fixed by immersion in 4% neutral buffered formalin from 2 to 6 hr at 4C, dehydrated by alcohol steps, transferred to xylol, embedded in paraffin, sectioned (10–12 μm) and placed on poly-L-lysine-coated slides. Free-floating sections were processed without pretreatment, while paraffin sections were deparaffinized and rehydrated.
Immunohistochemistry
Immunostaining was performed using the avidin-biotin complex (ABC) technique. Specificity, working dilution, and sources of the primary antibodies used in this work are summarized in Table 1. Free-floating sections were immuno-stained for VIP, CGRP, SP, NOS, VAChT, and CCK. Paraffin sections were used for PGP and NF-200. Briefly, sections were incubated for 30 min with 0.3% hydrogen peroxide in methanol to inhibit endogenous peroxidase activity. Sections were then incubated for 60 min with 5% normal serum diluted in PBS. The sections were incubated overnight with the primary antibody. The immunoreaction was revealed by incubating sections for 2 hr with biotinylated anti-rabbit (Dako; Carpinteria, CA), anti-goat (Vector Laboratories; Burlingame, CA) or anti-mouse (Dako) secondary antibody diluted 1:400 in PBS and by application of ABC Elite Reagent kit (Vector Laboratories) for 45 min. Peroxidase activity was revealed by diaminobenzidine (Sigma; St Louis, MO) alone or with 0.025% nickel chloride. Controls were performed replacing the primary antibody by 10% non-immune serum or by 3% PBS-BSA. Further controls were done by omitting the secondary antibody.
For NOS and VAChT double immunocytochemistry, the sections were sequentially incubated with anti-NOS antibody as described above, followed by biotinylated swine anti-rabbit IgG (Dako), ABC complex, and DAB development. The sections were then incubated with anti-VAChT antibody followed by biotinylated donkey anti-goat secondary antibody (Jackson Immunoresearch; West Grove, PA). This antibody was chosen for the absence of crossreactivity with swine proteins. Sections were then processed with avidin-biotinylated horseradish peroxidase and reacted with 3-amino-9-ethylcarbazole (AEC). The specimens were rinsed and mounted in aqueous medium.
Primary antibodies used
For VAChT and CGRP double immunocytochemistry, the sections were sequentially incubated with anti-VAChT antibody as described above, biotinylated donkey anti-goat IgG (Jackson Immunoresearch), ABC complex, and DAB development. The sections were then incubated with anti-CGRP antibody following by biotinylated swine anti-rabbit secondary antibody (Dako). This antibody was chosen for the absence of crossreactivity with donkey proteins. Sections were then processed with avidin-biotinylated horseradish peroxidase and reacted with AEC. The specimens were rinsed and mounted in aqueous medium.
For electron microscopic immunohistochemistry, free-floating sections were processed using the pre-embedding ABC complex method as previously described. The immuno-stained sections were postfixed in 1% osmium tetroxide, dehydrated in graded concentrations of acetone, and embedded in a mixture of Epon and Araldite. Semithin sections were examined by light microscopy to locate areas containing immunoreactivity. Ultrathin sections were observed unstained in an EM 10 electron microscope (Zeiss; Oberkochen, Germany).
NADPHd Cytochemistry
Animals (n = 6) were anesthetized with ether and perfused with 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4). Blocks of lingual tissue were processed as previously described (Sbarbati et al. 1999).
AChE Cytochemistry
Enzymatic staining of AChE for light microscopy was performed according the direct thiocholine method of Karnowsky and Roots (1964). Adult Wistar rats (n = 8) were used. Tongues were removed from animals and VP was excised and fixed in 4% paraformaldehyde in 0.4 M phosphate buffer for 1 hr at room temperature. Tissues were put into 20% sucrose in phosphate buffer for 1 hr at 4C and sectioned on a freezing microtome (Reichert-Jung). Free-floating sections (thickness 30 μm) were preincubated with 0.1 M acetate buffer, pH 6, for 30 min and incubated for 30 min in AChE staining solution containing 5 mg acetylthiocholine iodide, 0.5 ml 0.1 M sodium citrate, 1 ml 30 mM copper sulfate, 1 ml 5 mM potassium ferricyanide, 1 ml distilled H2O, 6.5 ml 0.1 M sodium acetate buffer, pH 6.0. Control sections were incubated without acetylthiocholine iodide. After rinses in acetate buffer, free-floating sections were mounted on glass slides, dehydrated, coverslipped with Entellan, and observed in a Orthoplan 2 photomicroscope (Leitz; Wetzlar, Germany).
Cell Counts
We have evaluated the numerical ratio between AChE- and NADPHd-positive cells by a morphometrical approach. Quantitative evaluations of AChE- and NADPHd-positive cells were performed on serially sectioned VP (AChE n = 3; NADPHd n = 3) after cytochemical staining using sections cut at a 34-μm setting. The thickness of the whole VP ranged from 646 to 850 μm. Positive cells were counted in the area under the VP, in the region between two parallel lines passing through the lumen of the vallum. Considering that the neurons are distributed in small ganglia that could be missed in a systematized sampling protocol, we performed the counts in all the sections. Using this approach, the possibility exists that neurons are counted twice on adjacent sections. To reduce the overestimation involved in this approach, only nucleated cells were counted.
Co-localization of AChE or Peptides with NADPHd
In co-localization experiments, the method was based on two further steps of staining. In a first step we performed AChE staining or immunocytochemistry for peptides on floating slices with the above described methods. The sections were subsequently mounted in glycerol and observed by light microscopy (Leitz). Images were recorded using a KY-F58 CCD camera (JVC). In each section, all the positive elements were identified and photographed. The images were analyzed and stored using the software Image-ProPlus on a personal computer. In a further step, the same slices used for AChE detection or immunocytochemistry were washed in PB and processed for NADPHd cytochemistry. For light microscopy, the stained sections were mounted with Entellan. The images obtained after the first step were compared with those obtained from the same cells after the second staining. NADPHd reaction causes a very dark cytoplasmic precipitate. Therefore, the comparison of the image obtained from the same cell after the first and the second step enabled us to obtain information about the co-localization patterns.
Results
General Morphology
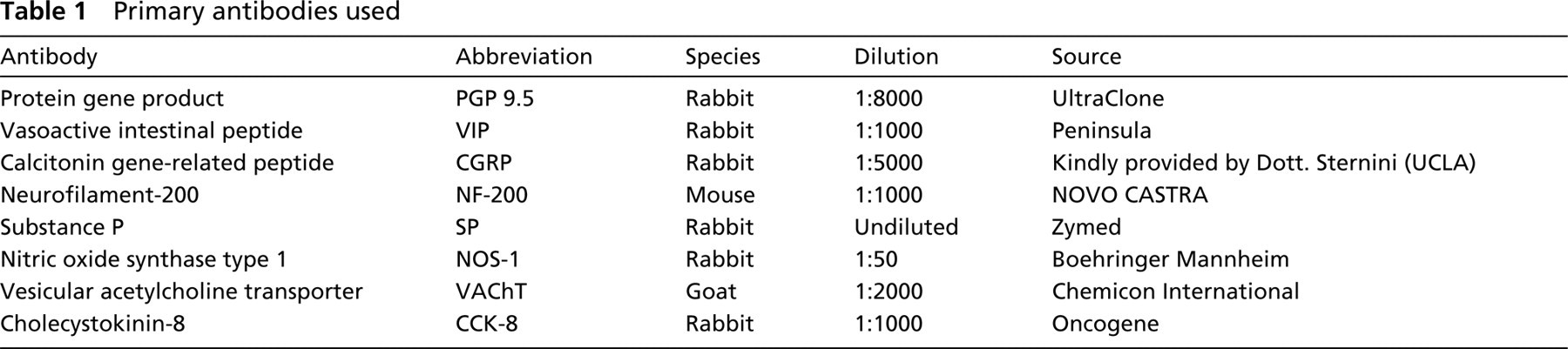
The VP/VEG complexes are composed of an epithelial papilla (i.e., the VP) and an associated gland, i.e., the VEG (Figure 1). This latter structure (also called posterior lingual gland) is a serous salivary gland. In each papilla the taste buds are strictly associated with the terminal ends of the VEG duct system. A specialized portion of the peripheral nervous system is associated with the structure. In the VP/VEG complex, a main ganglion is located at the base of the VP and is visible using NADPHd (Figure 1b), AChE (Figure 1c) cy-tochemical staining or immunocytochemistry for a neuronal marker (Figure 1d). In previous works, the main ganglion was called Remak's ganglion (Remak 1852). However, this term may generate confusion with the Remak's ganglion in the duodenum of birds. Therefore, the term VP/VEG ganglion will be used in the present work. Nerve fibers originating from this ganglion penetrate the tongue, reaching more deeply located ganglia.
Co-localization of AChE/NADPHd
A detailed anatomic description of the intrinsic nervous system of the VP/VEG complex was obtained by cytochemical staining for AChE (Figures 1c and 2a–2d). By this technique, the main ganglion located at the base of the VP was visible. AChE-positive nerve fibers were also located below the gustatory epithelium. Several neurons were also scattered in the core of the VP, around the trench and in the VEG. Data about the NADPHd staining of the VP/VEG complex have been previously reported (Sbarbati et al. 1999). The staining pattern obtained by NADPHd was basically similar to that obtained by AChE. In each VP, 291 ± 11 NADPHd-positive cells (mean ± SD) and 192 ± 12 AChE-positive cells were found. Therefore, the two populations seem to be in a 1:1.5 ratio.
General morphology of the VP/VEG complex. (
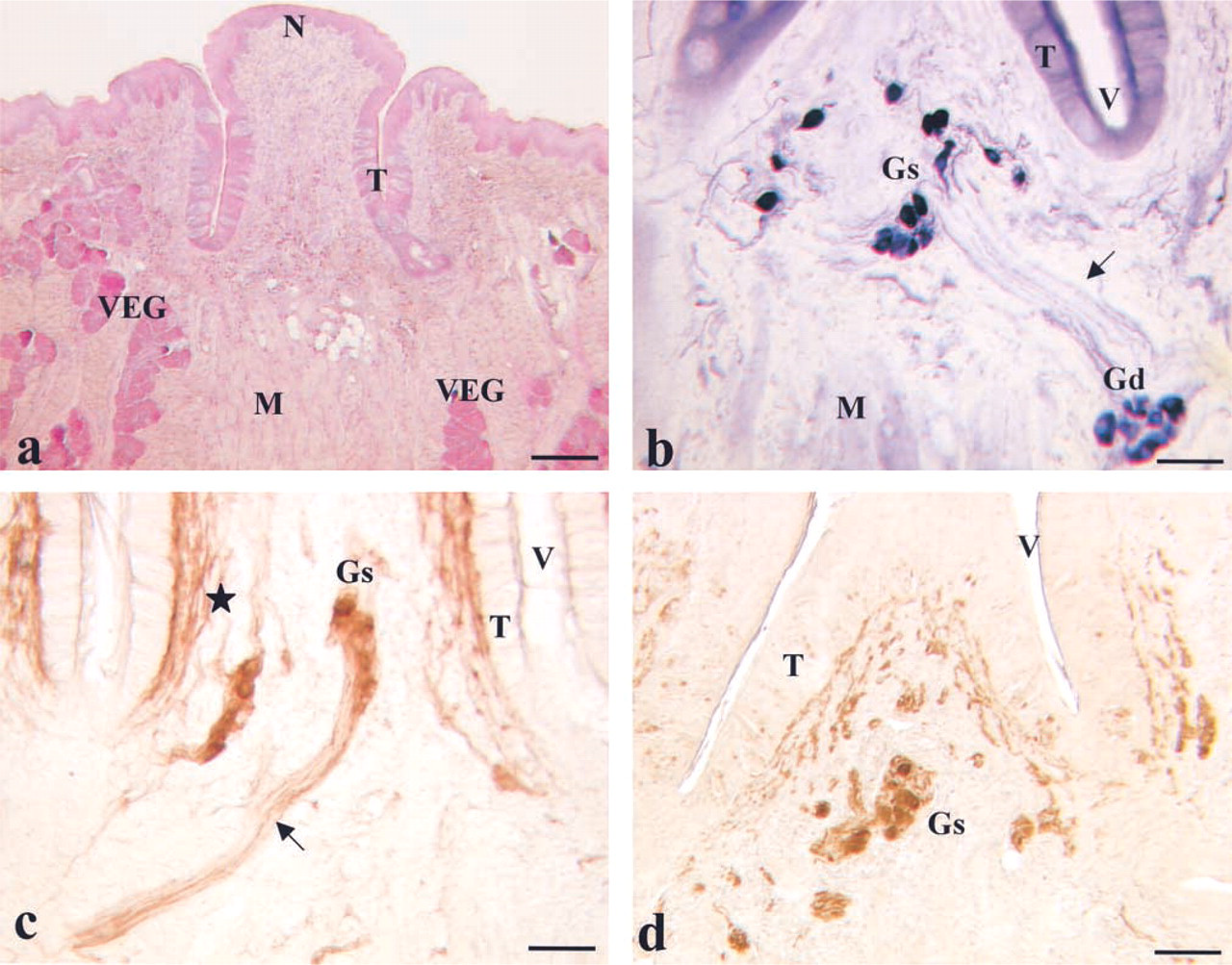
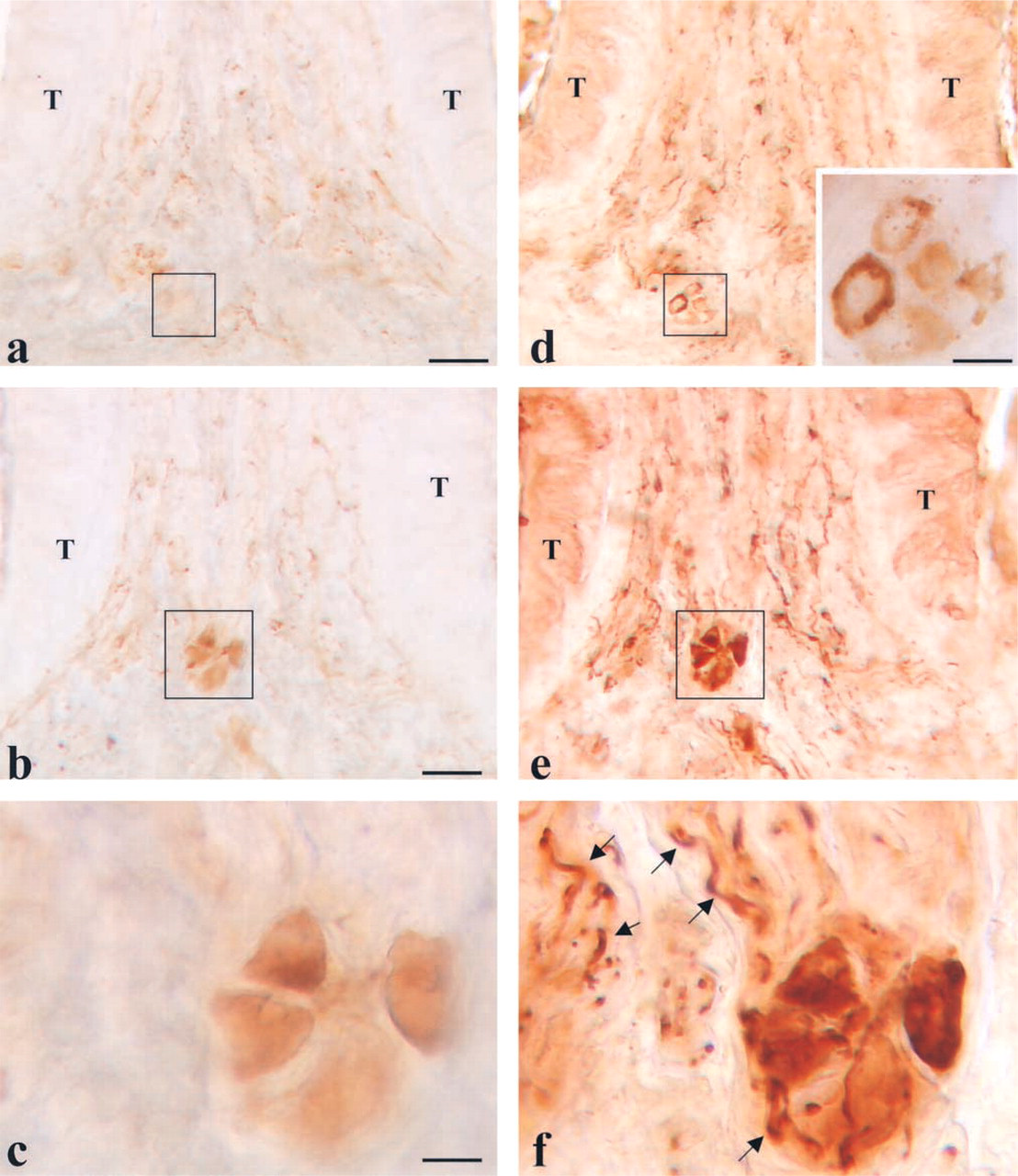
The presence of possible differences between the two staining procedures was studied on the same slice (Figures 2e–2h). Using this paradigm, three different neuronal subpopulations were detected in the VP/VEG ganglion. The main population was composed of AChE+/NADPHd+ cells. Small groups of elements that were AChE+/NADPHd— were visible in the VP/VEG ganglion. They regularly showed a large diameter and a round shape. Isolated AChE—/NADPHd+ cells were visible at the periphery of the VP/VEG ganglion. These cells were small neurons with an ovoid shape.
Co-localization of AChE (brown) and NADPHd (dark blue). (
Immunocytochemical Analysis
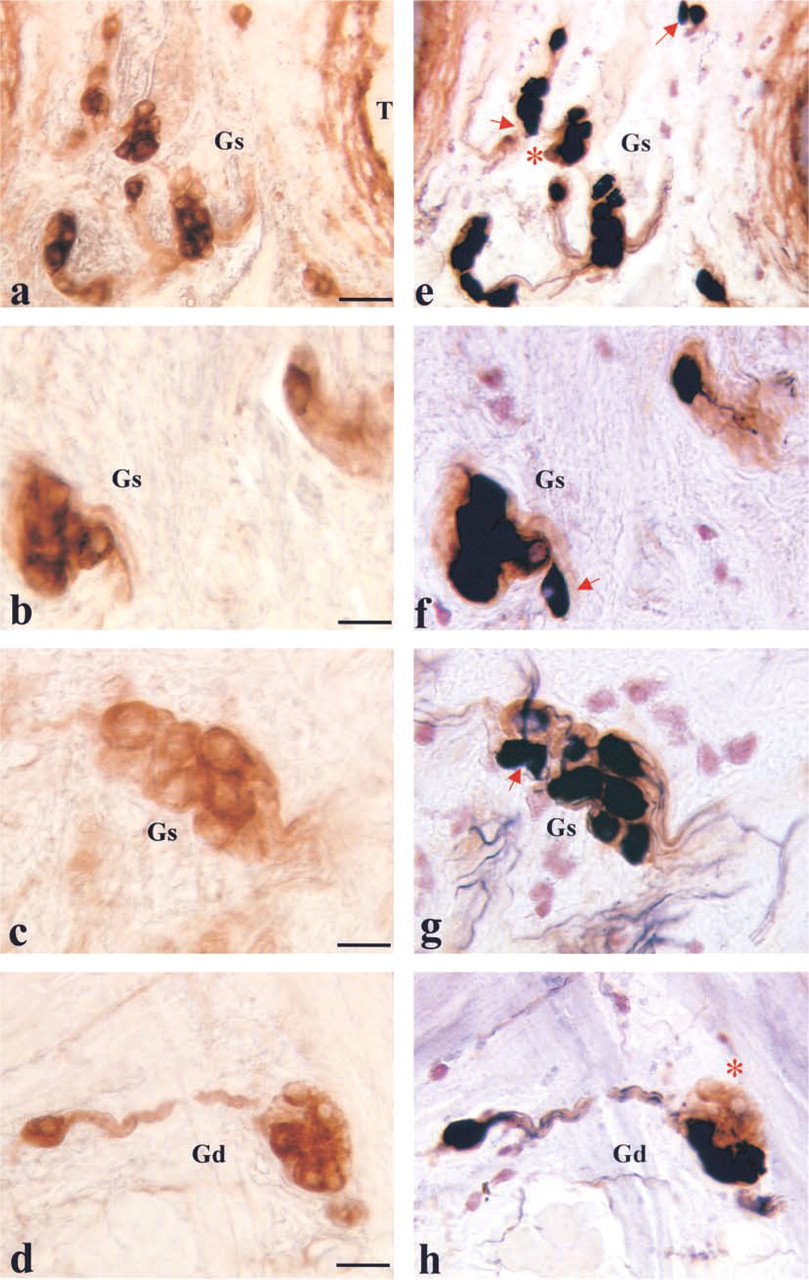
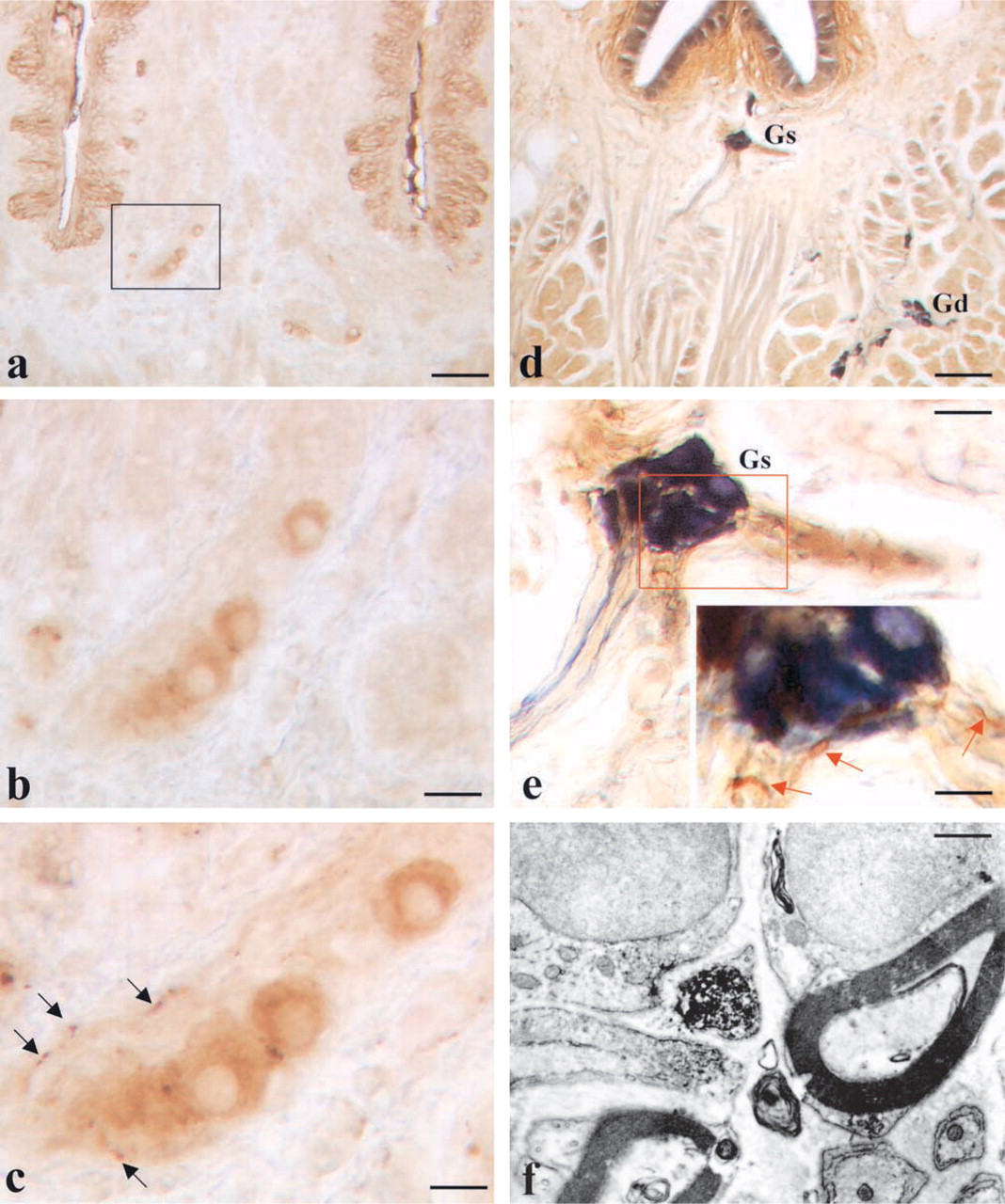
The immunocytochemical specificity of the intrinsic nervous system was studied by a panel of antibodies. NF200 immunostaining weakly stained the cell bodies in the ganglia and the nerve fibers (Figure 3a). The cell bodies of the ganglionic neurons were also immuno-stained by antibodies against PGP 9.5, a general nerve marker (Figure 1d).
A large part of the neurons of the intrinsic system were positive at immunostaining for VIP (Figure 3b), NOS (Figure 3c), or VAChT (Figure 3d). Only a small subset of nerve fibers revealed immunoreactivity for CCK (Figure 3e). In the VP/VEG complex, varicose axons traveling in the lamina propria under the lingual epithelium can be evidenced by their content of neuropeptides such as SP (Figure 3f) or CGRP (Figure 3g). In the intrinsic nervous system, no cell bodies positive for SP or CGRP were detected either in the superficial or in the deep portion. Control specimens were always negative (Figure 3h).
Co-localization of VAChT/NOS
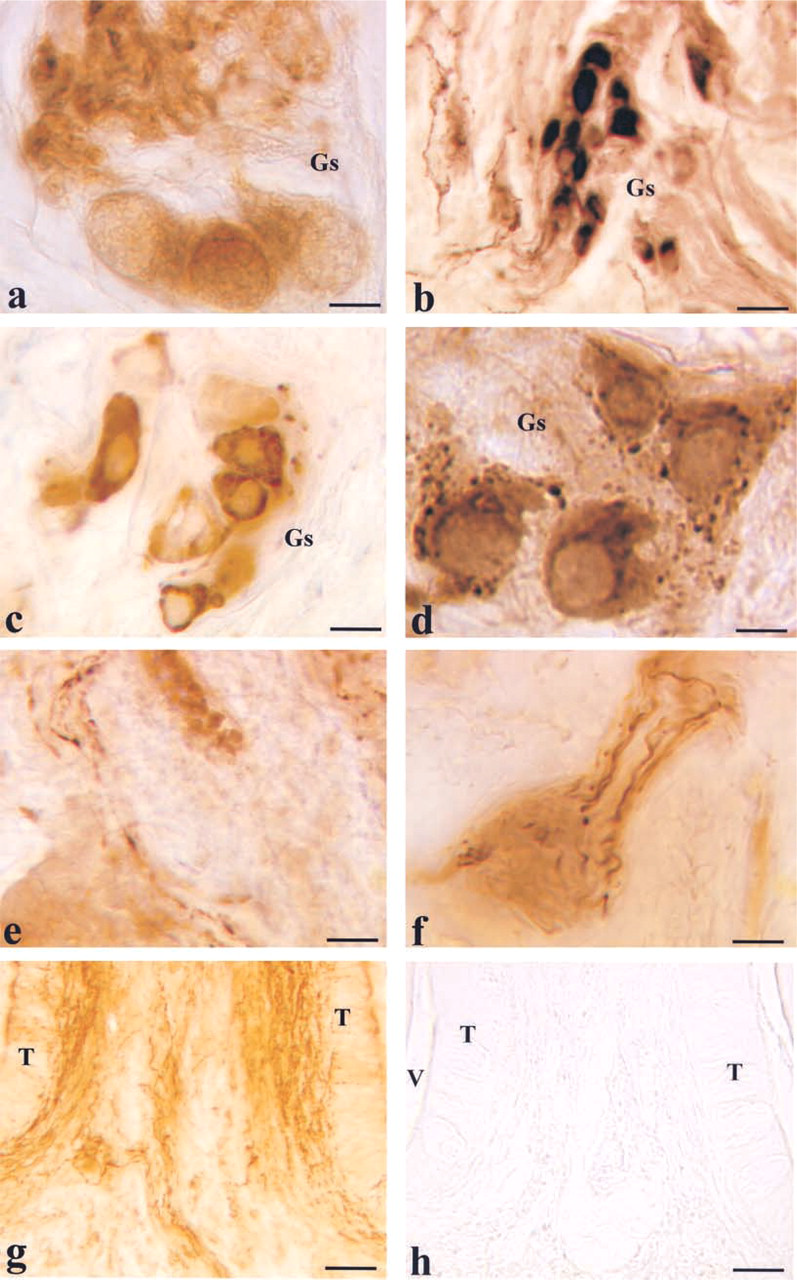
To further characterize the reciprocal relationship between cholinergic and nitrergic elements of the intrinsic nervous system, we performed co-localization experiments for VAChT and NOS that can be considered specific markers of the two components. The presence of possible differences was studied on the same slice by the same computer-aided method of analysis used to compare cytochemical staining. The findings obtained by this paradigm were consistent with those obtained by cytochemical co-localization of AChE and NADPHd. The staining pattern obtained by the two antibodies was basically similar. However, in the main ganglion VAChT+/NOS — cell bodies were visible (Figures 4a and 4d). In addition, a contingent composed of VAChT+/NOS— fibers was found associated with the groups of neurons (Figures 4b, 4c, 4e, and 4f).
Co-localization of VAChT/CGRP
Co-localization experiments for VAChT and CGRP revealed that the VP/VEG/VAChT+ ganglion is surrounded by CGRP-immunoreactive fibers (Figure 5c).
Co-localization of Peptides (CGRP or SP)/NADPHd
Co-localization experiments for peptidergic and nitrergic structures revealed that the VP/VEG ganglion is surrounded by CGRP-immunoreactive fibers (Figures 5d and 5e). Substance P-immunoreactive fibers were also found in the ganglion (not shown).
Ultrastructural Immunocytochemistry
Ultrastructural analysis also confirmed the presence of CGRP- (Figure 5f) or SP-immunoreactive (not shown) nerve fibers in the VP/VEG ganglion. In the specimens examined, no clear images of synapses between CGRP-immunoreactive fibers and neurons were detected.
Discussion
In the past, several studies described neurons associated with the VP (Remak 1852; Drasch 1887; Fusari and Panasci 1889; Kane and Law 1950; Gairns and Garven 1952; Baradi and Bourne 1959; Brizzi 1969; Fitzgerald and Alexander 1969; Graziadei and Monti Graziadei 1978; Bradley et al. 1985). We have recently described the anatomic organization of these neurons (Sbarbati et al. 1999) and their development (Sbarbati et al. 2000). No specific studies have neuro-chemically characterized these neurons and described the presence of different subclasses. In addition, previous immunocytochemical studies of the VP focused mainly on the gustatory system, paying scarce attention to the intrinsic neurons. Therefore, a precise identification of the chemical coding of the intrinsic nervous system annexed to the VP and VEG is missing. The present work provides further data on the morphology and chemical specificity of its intrinsic neurons.
The results demonstrated the presence of peptidergic, cholinergic, and nitrergic intrinsic neurons. The ganglia show SP- and CGRP-immunoreactive fibers. Such fibers are also located in the gustatory epithelium (Luts et al. 1990), and SP and CGRP are known to coexist in primary sensory neurons innervating several organs (Luts et al. 1990). In other locations, SP- or CGRP-immunoreactive fibers in association with sympathetic and parasympathetic ganglia are known, and their possible origin has been suggested to be collaterals from primary sensory nerve fibers (Suzuki et al. 1998). Our data appear to confirm findings obtained in the anterior two thirds of the dog tongue (Hino et al. 1993; Hu et al. 1996) describing peptide-containing sensory fibers in NADPHd-positive ganglia and suggesting an intrinsic peripheral nerve reflex mechanism in the regulation of lingual vascular function (Hu et al. 1996). Further studies are necessary to evaluate if the peptidergic fibers that we have demonstrated in the VP/VEG ganglion are collaterals of gustatory afferent fibers mediating reflex actions involving the gland. In the present work, we have performed a large series of experiments of immunostaining and co-localization and the results demonstrate that different types of neurons can be identified in the VP/VEG complex. The finding obtained by experiments of co-localization for AChE/NADPHd and those of co-localization for VAChT/NOS are consistent and both suggest that nitrergic/cholinergic, nitrergic/non-cholinergic, and cholinergic/non-nitrergic neurons exist in the VP/VEG ganglion. The presence of different neuronal subclasses suggests that in the VP/VEG complex a certain degree of functional specialization may also exist. However, further data about projections and chemical coding of intrinsic neurons of the VP/VEG complex must be obtained for a definitive confirmation of this hypothesis.
Immunostaining of the VP/VEG complex for different antibodies. (
(
(
Morphological and neurochemical results must not be overestimated in the absence of data on function and connectivity of the different components of the intrinsic nervous system. However, the description of neuronal subpopulations obtained in the present work suggests that further studies must be performed with the aim of evaluating the presence of intrinsic primary sensory neurons and interneurons in the VP/VEG complex.