
Abstract
The cystatin superfamily of cysteine protease inhibitors and target cysteine proteases such as cathepsin B have been implicated in malignant progression. The respective cellular/extracellular localization of cystatins and cysteine proteases in tumors may be critical in regulating activity of the enzymes. Confocal microscopy has enabled us to demonstrate the differential localization of cystatins and cathepsin B in an embryonic liver cell line and an invasive hepatoma cell line. In both, stefins A and B were distributed diffusely throughout the cytoplasm, whereas cystatin C was distributed in juxtanuclear vesicles. Stefin A and cystatin C, but not stefin B, were present on the cell surface. Cystatin C was found on the top surfaces of both cell lines, whereas stefin A was found only on the top surface of the embryonic liver cells. Cathepsin B staining was concentrated in perinuclear vesicles in the embryonic liver cells. In the hepatoma cells, staining for cathepsin B was also present in vesicles adjacent to the cell membrane and on localized regions of the bottom surface. Such a disparate distribution of cathepsin B and its endogenous inhibitors may facilitate proteolysis by the hepatoma cells and thereby contribute to their invasive phenotype.
P
The subcellular localization of the stefins and cystatins can be inferred from sequence analysis of their cDNAs. Stefins A and B are presumed to be cytoplasmic because the amino acid sequences deduced from their cDNAs do not have signal peptides (Kartasova et al. 1987; Sato et al. 1990, 1992). Both stefins A and B have been isolated from supernatants of human spleen (Järvinen and Rinne 1982) and human liver (Green et al. 1984). On the other hand, they can also be isolated from human urine (Abrahamson et al. 1986). This may indicate a passive release from the cytoplasm of damaged or dying cells, or alternatively, active secretion from a vesicular compartment. The cDNA of cystatin C suggests that this inhibitor is secreted, because it contains a sequence coding for a signal peptide (Abrahamson et al. 1987). Cystatin C is a constituent of many biological fluids, the highest levels being in seminal plasma, cerebral spinal fluid, and synovial fluid (Abrahamson et al. 1986). The isolation of stefins A and B from cell supernatants and the presence of cystatin C in many biological fluids are consistent with the absence and presence, respectively, of signal peptides.
A membrane localization for stefins and cystatins would not be predicted from their cDNA sequences. However, in murine tumors, we have observed an enrichment of cystatin activity in a plasma membrane fraction along with a decrease in overall cystatin activity (Rozhin et al. 1990). Subsequently, we showed that the cystatin purified from the total membrane fraction of the invasive murine hepatoma Hepa cl 9 (utilized in this study) is immunologically related to stefin A (Moin et al. 1992). Therefore, there is evidence from biochemical studies for localization of low molecular mass cystatins that would not be predicted from their cDNA sequences.
Previous immunohistochemical studies have not shed light on these discrepancies because their primary goal had been to identify tissues positive for specific low molecular mass cystatins rather than to determine subcellular/extracellular localization. Therefore, in the present study, we employed immunofluorescent confocal microscopy to compare the localization of the cystatins and the target cysteine protease cathepsin B in BNL CL.2 murine embryonic liver cells and Hepa cl 9 murine liver tumor cells. This enabled us to assess whether there were any differences in the subcellular and surface distributions of stefins A and B, cystatin C, and cathepsin B. We were particularly interested in differences that might affect the invasive phenotype of Hepa cl 9 cells.
Materials and Methods
Materials
Dulbecco's modified Eagle's medium (DMEM) was purchased from Sigma (St Louis, MO); secondary antibodies (Texas Red-conjugated affinity-purified donkey anti-rabbit or anti-mouse IgGs and fluorescein-conjugated affinity-purified donkey anti-rabbit or anti-mouse IgGs) and normal donkey serum from Jackson ImmunoResearch (West Grove, PA); and the Slow Fade anti-fade reagent from Molecular Probes (Eugene, OR). Polyclonal antibodies to rat epidermis cystatin-α and mouse cystatin C were kind gifts from Dr. Kimie Fukuyuma (University of California at San Francisco, CA) and Dr. Magnus Abrahamson (University of Lund, Lund, Sweden), respectively. Monoclonal antibody to human stefin B was a kind gift from Dr. Vito Turk (Jozef Stefan Institute, Ljubljana, Slovenia). The specificities of the antibodies were established by immunoblot analysis (Moin et al. 1992; Hakansson et al. 1996). Polyclonal antibody to the double-chain form of human liver cathepsin B was developed and its specificity confirmed in our laboratory (Sloane et al. 1994b). All other materials were of reagent grade.
Cells and Culture Conditions
Hepa cl 9 is a subclone of Hepa cl 7, originally derived from a murine hepatoma (Forster-Gibson et al. 1988), and was a kind gift from Dr. Michael Dufresne, (University of Windsor, Windsor, Ontario, Canada). Hepa cl 9 was grown and maintained in DMEM containing 10% fetal calf serum. BNL CL.2, an embryonic liver cell line derived from a BALB/c mouse (Patek et al. 1978) (American Type Culture Collection; Rockville, MD), was grown and maintained in DMEM containing 10% fetal calf serum and 4.5 g/liter glucose.
Immunolocalization
Intracellular Labeling. Intracellular stefins A and B, cystatin C, and cathepsin B were localized using our published immunocytochemical protocols (Sloane et al. 1994b; Sameni et al. 1995). Cells were grown on glass coverslips to ∼ 70% confluency. After an overnight incubation in serum-free medium, cells were washed at 25C with PBS, pH 7.4, and fixed with 100% methanol at -20C or with 3.7% formaldehyde in PBS at 25C. The choice of fixatives was dictated by optimal preservation of the antigen in the cell line being studied. All further steps were performed at 25C. Cells were washed with PBS and blocked with PBS containing 2 mg/ml bovine serum albumin. The membrane permeant saponin (0.1%) was added to subsequent antibody and PBS wash solutions. Cells were incubated with primary antibodies (rabbit antirat cystatin-α IgG, which is crossreactive with mouse stefin A, rabbit anti-mouse cystatin C IgG, or mouse anti-human liver stefin B IgG) for 2 hr. In controls, either preimmune serum (rabbit or mouse) or PBS-saponin was substituted for the primary antibody. After washing, cells were blocked with normal donkey serum (5% in PBS-saponin) in the presence of the secondary antibody: either Texas Red-conjugated or fluorescein-conjugated affinity-purified donkey anti-rabbit IgG or mouse IgG at 20 μg/ml. After washing, the coverslips were mounted upside down on slides with Slow Fade anti-fade reagent and observed on a Zeiss LSM-310 confocal microscope.
Surface Labeling. Surface stefins A and B, cystatin C, and cathepsin B were localized on living cells by the following modification of the procedures described above. Incubation with the primary antibody was performed at 4C to prevent endocytosis of antibodies, followed by fixation with 3.7% formaldehyde, saponin was omitted, and only Texas Red-conjugated secondary antibodies were used.
Confocal Image Analysis. For each antigen, 40 consecutive optical sections were obtained utilizing the LSM-310 in confocal mode; individual sections were 0.25 μm and the 40 sections encompassed the entire cell volume. Three-dimensional images and galleries of the optical sections were analyzed to demonstrate the spatial distribution of cell surface cathepsin B and cystatins. The three-dimensional images were assembled from all 40 optical sections utilizing the software VoxelViewUltra from Vital Images (Fairfield, IA). Galleries of confocal images were assembled to encompass the entire cell volume. For cystatin C and stefin A, the galleries contained 25 images of 512 X 768 pixels and, for cathepsin B, 24 images of 512 X 512 pixels.
Results
Intracellular Localization
The staining for stefin A (Figures 1A and 1B) and stefin B (Figures 1C and 1D) was diffuse and was distributed throughout the cytoplasm of both the embryonic liver (Figures 1A and 1C) and hepatoma (Figures 1B and 1D) cells. In the hepatoma cells, the many vesicles presented a negative image because of an apparent concentration of stefins A and B at their membranes (see arrows, Figures 1B and 1D). In contrast, staining for cystatin C was punctate and thus presumably vesicular (Figure 2). The distribution of the vesicles staining for cystatin C was primarily juxtanuclear and was similar in embryonic liver (Figure 2A) and hepatoma (Figure 2B) cells.
One of the enzymes presumed to be a target of the low molecular mass cystatins is the lysosomal cysteine protease cathepsin B. We had established previously that cathepsin B is localized in perinuclear vesicles in normal cells. In transformed and tumor cells, cathepsin B is distributed both in perinuclear vesicles and in vesicles throughout the cytoplasm, including vesicles at the cell periphery and in cell processes (Sloane et al. 1994b; Sameni et al. 1995). Therefore, we compared the intracellular localization of cathepsin B in the embryonic liver and hepatoma cells. In both cell types cathepsin B staining was vesicular, but the distribution of vesicles staining for cathepsin B differed. In the embryonic liver cells, the vesicles staining for cathepsin B were concentrated in the perinuclear region (Figure 3A), whereas in the hepatoma cells the vesicles were found in both the perinuclear and peripheral regions of the cells (Figure 3B).
Surface Localization
Our previous observations of an enrichment of cystatin activity in a plasma membrane fraction of murine B16 melanomas and Lewis lung carcinomas (Rozhin et al. 1990), as well as the presence of a stefin A-like protein in the total membrane fraction of murine Hepa cl 9 hepatoma cells (Moin et al. 1992), led us to investigate whether cystatins (stefins A and B and cystatin C) might be present on the surface of nonpermeabilized murine embryonic liver and hepatoma cells. We did not observe any staining for stefin B on the surface of either the embryonic liver or the hepatoma cell line (data not shown). Staining for cystatin C was present in discrete patches on the surface of both cell lines (Figures 4C and 4D). Similar discrete patches of staining for stefin A were present on the surface of the embryonic liver cells (Figure 4A), but not on the surface of the hepatoma cells (Figure 4B). To determine whether the surface staining for cystatin C and stefin A on the embryonic liver cells was specific for the top or the bottom of cells, we obtained serial optical sections of the staining and used these to reconstruct three-dimensional images. The reconstructions show that the levels of surface staining on embryonic liver cells were similar for stefin A and cystatin C (Figures 5A and 5B). This assessment is subjective rather than based on objective measurements. The optical sections show that the surface staining for stefin A was primarily at the top of the cells (Figure 5C), whereas the surface staining for cystatin C was distributed on both the top and lateral surfaces of the embryonic liver cells (Figure 5D). Surface staining for stefin A or cystatin C was not present in the bottom sections of the embryonic liver cells.
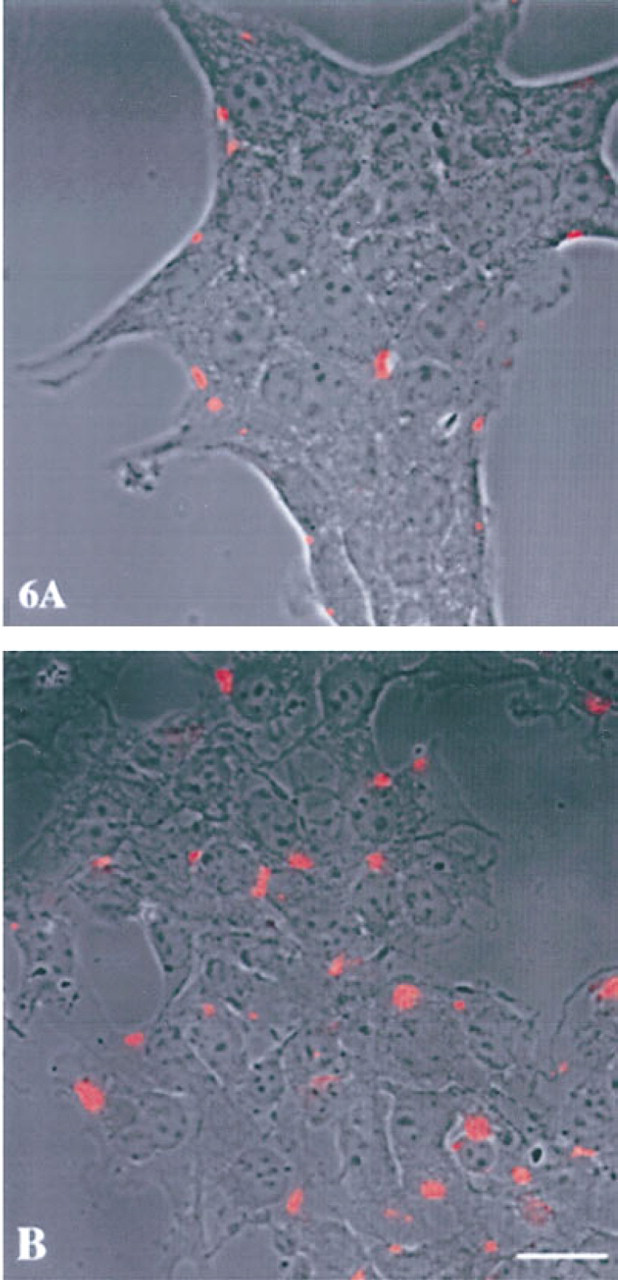
We had established previously that cathepsin B staining is localized on the bottom surface of transformed breast epithelial cells, breast tumor cells, and glioblastoma cells (Sloane et al. 1994b; Sameni et al. 1995; Sloane 1996). In contrast, we found little or no staining for cathepsin B on the surface of normal breast epithelial cells. Therefore, we compared the surface staining for cathepsin B on nonpermeabilized embryonic liver and hepatoma cells. We observed many discrete patches of staining for cathepsin B on the surface of the hepatoma cells (Figure 6B). There were also discrete patches of staining for cathepsin B on the surface of the embryonic liver cells (Figure 6A), but these areas were both fewer in number and smaller in size than those observed on the hepatoma cells. To determine whether the surface distribution of cathepsin B was similar to that of the cystatins, we obtained serial optical sections of the staining and used these to reconstruct three-dimensional images. Reconstructions and galleries of the surface staining for cathepsin B showed that there was substantially greater surface staining on hepatoma cells than on embryonic liver cells (not illustrated) and that staining for cathepsin B was on the lateral and bottom surfaces of both embryonic liver (not illustrated) and hepatoma (Figure 7D) cells. As indicated above, cystatin C was also present on the surface of hepatoma cells. We compared the surface localization of cystatin C (Figure 7A) and cathepsin B (Figure 7B) by using serial optical sections of the staining to reconstruct three-dimensional images. The reconstructions of surface staining on the hepatoma cells indicated similar levels of staining for cystatin C and cathepsin B. However, the optical sections showed that the distribution of surface staining differed. Cystatin C was present on the top surface (Figure 7C) and cathepsin B on the bottom surface (Figure 7D) of the hepatoma cells.
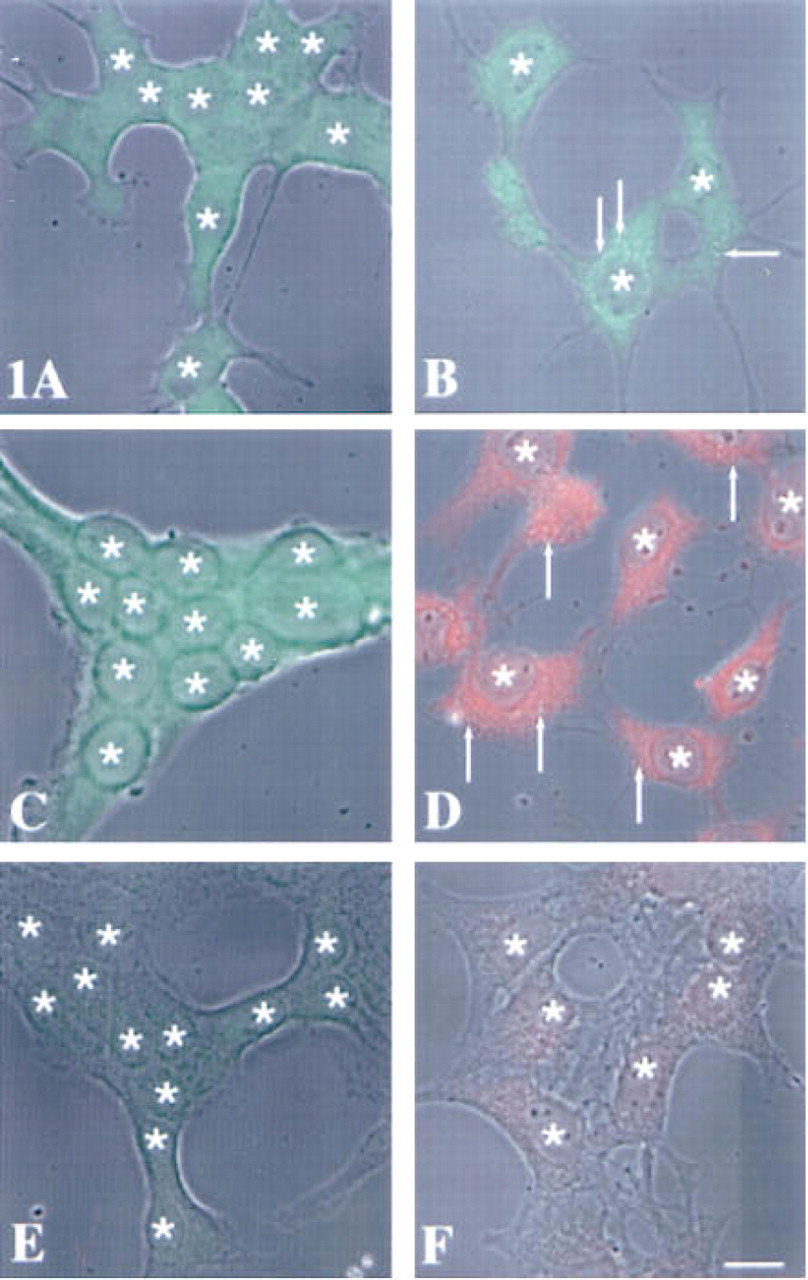
Superimposition of immunofluorescent staining for intracellular stefins A and B on phase-contrast images of murine embryonic liver
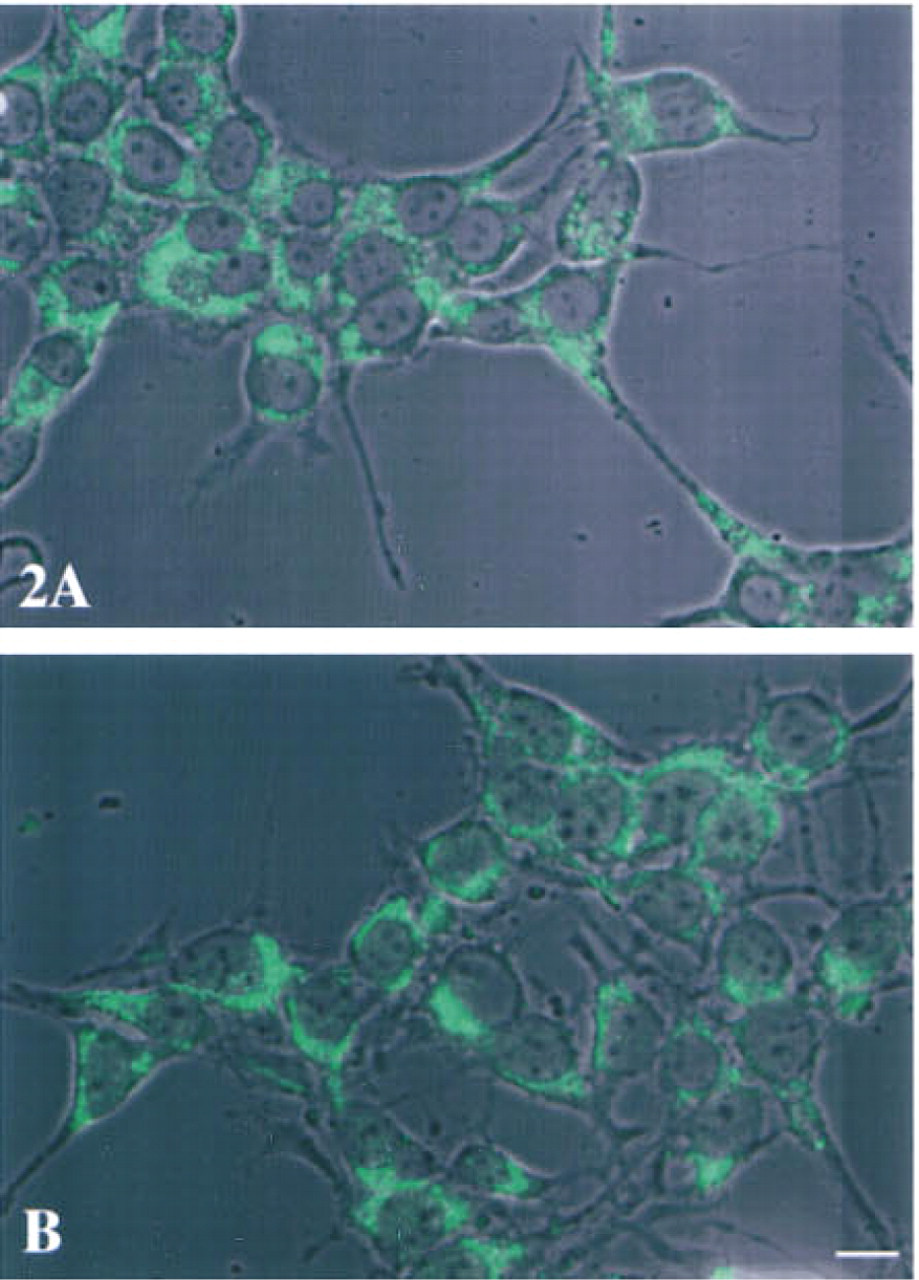
Superimposition of immunofluorescent staining for intracellular cystatin C on phase-contrast images of murine embryonic liver
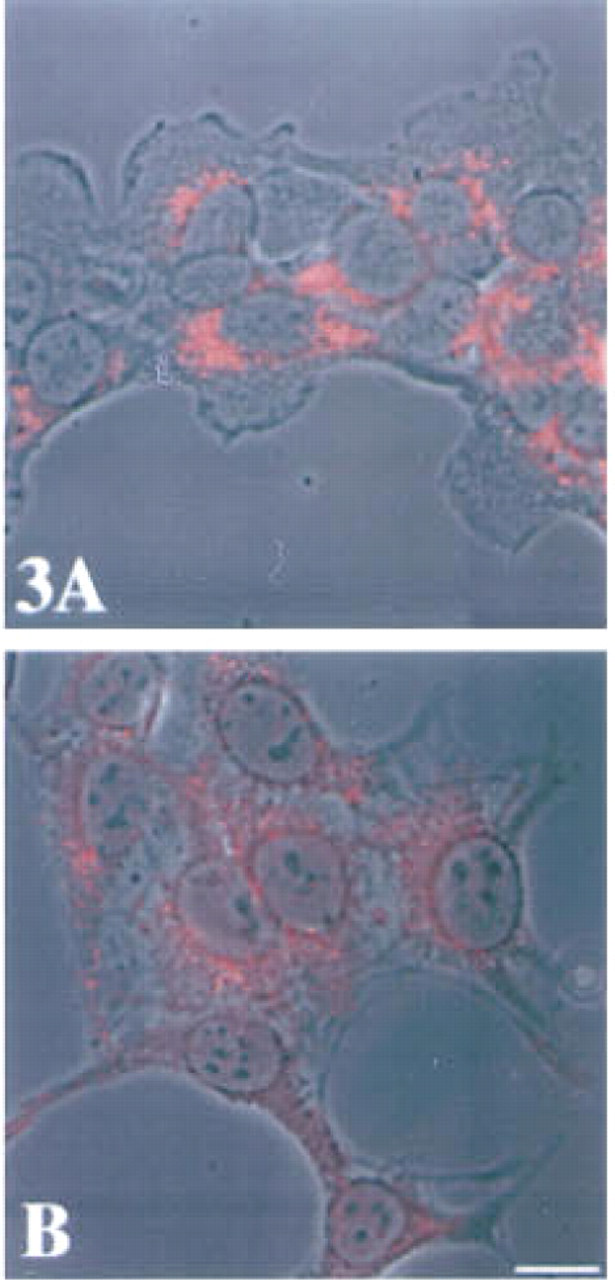
Superimposition of immunofluorescent staining for intracellular cathepsin B on phase-contrast images of murine embryonic liver
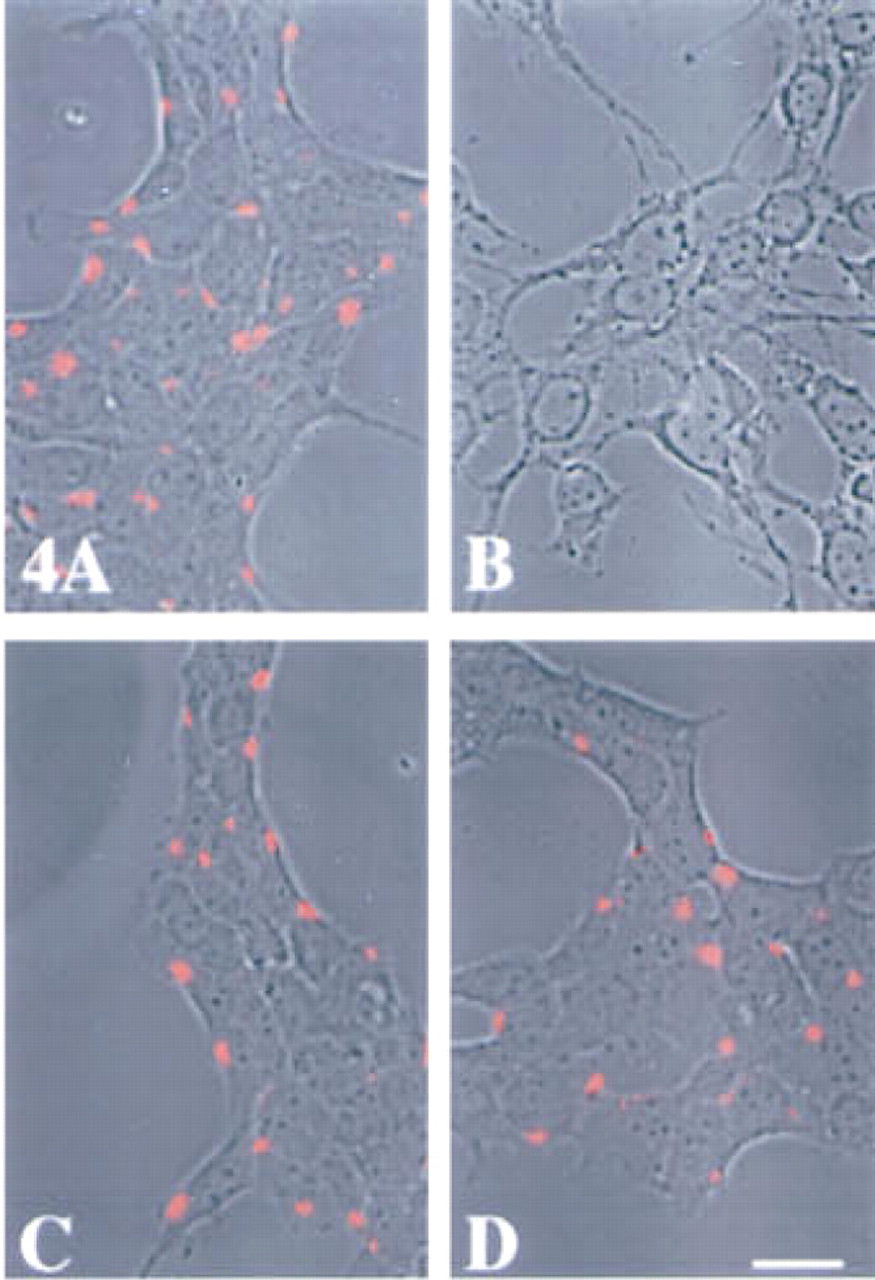
Superimposition of immunofluorescent staining for surface stefin A and cystatin C on phase-contrast images of murine embryonic liver
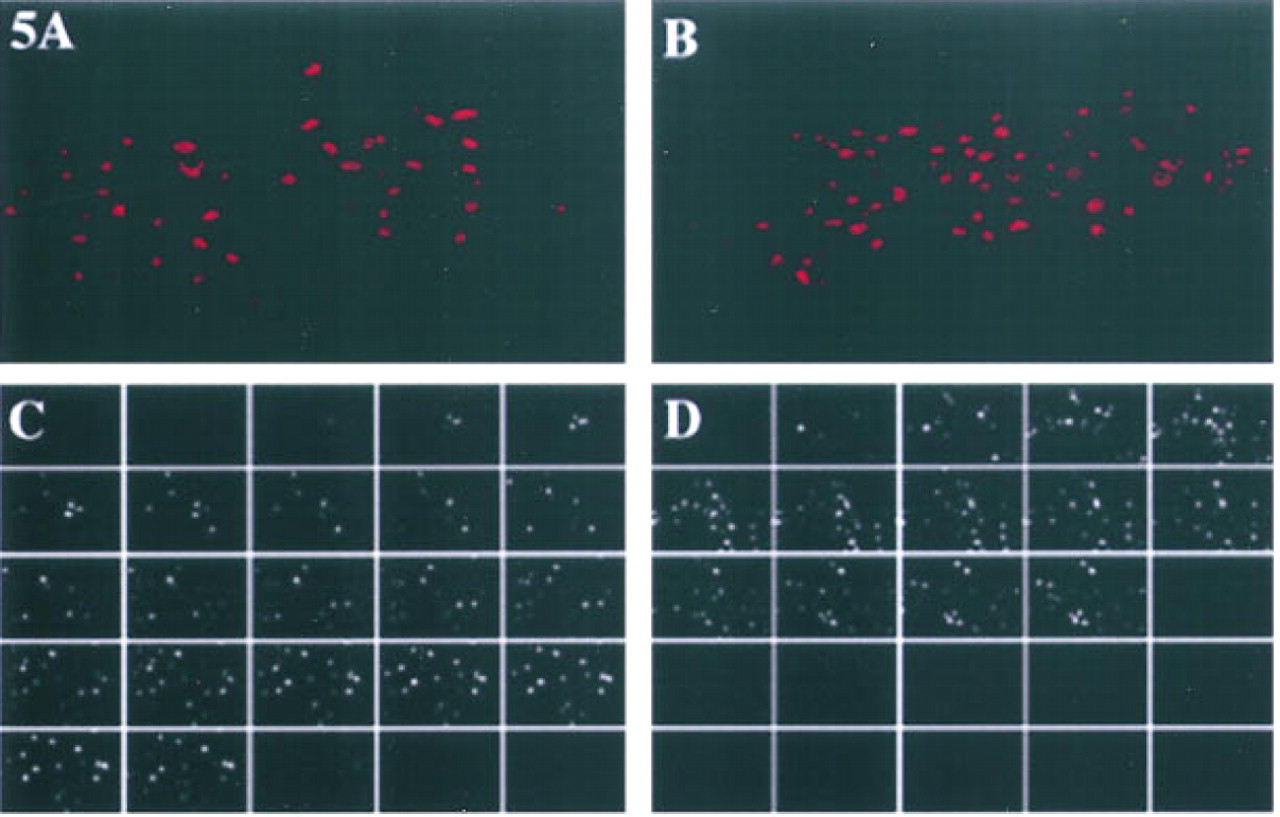
Three-dimensional reconstruction of immunofluorescent staining for stefin A
Superimposition of immunofluorescent staining for surface cathepsin B on phase-contrast images of murine embryonic liver
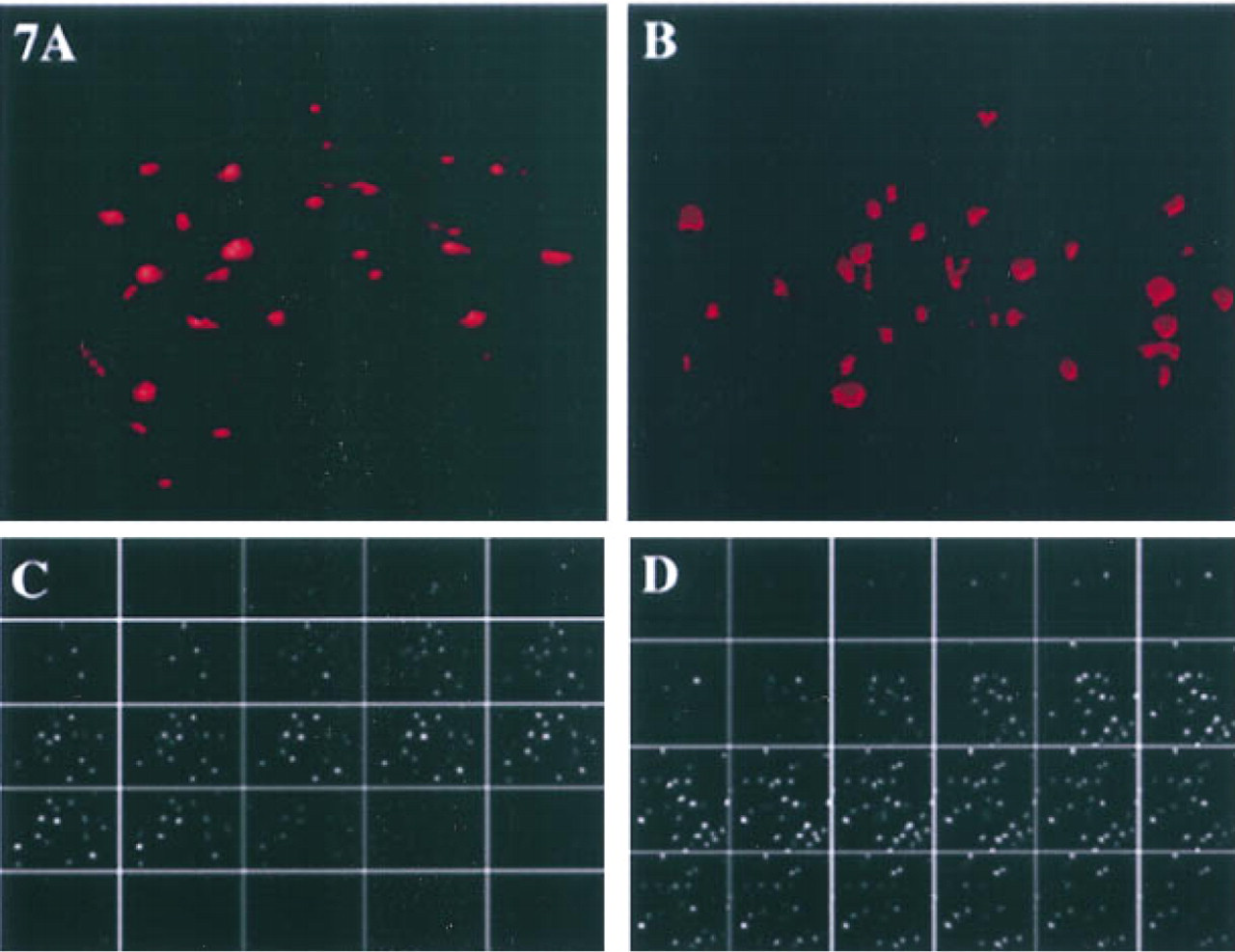
Three-dimensional reconstruction of immunofluorescent staining for cystatin C
Discussion
Our immunofluorescent studies confirmed a cytoplasmic localization for stefins A and B and a vesicular localization for cystatin C in murine cell lines of liver origin. A surface association of stefin A was observed only in the embryonic liver cells. In contrast, cystatin C was associated with the surface membranes of both cell lines. A cytoplasmic localization for stefin A and B and vesicular localization for cystatin C is consistent with the absence and presence of signal peptides in their respective cDNAs (Abrahamson et al. 1987; Kartasova et al. 1987; Sato et al. 1990, 1992). The target enzymes for these cystatins are cysteine proteases, e.g. cathepsins B, L, and S. These cysteine proteases are localized intracellularly in the vesicular compartment, specifically in the lysosomes and endosomes, and are not usually present on the cell surface. Therefore, although stefins A and B and cystatin C all inhibit cathepsins B, L and S in in vitro assays (Barrett et al. 1984; Green et al. 1984; Abrahamson et al. 1986; Bromme et al. 1991), the site at which cystatins interact with these cysteine proteases in situ has not been known.
In the present study, the staining for stefins A and B in the tumor cells appeared to be concentrated adjacent to vesicular membranes. The pattern of staining suggested that stefins A and B may interact specifically with certain vesicular membranes. There are previous reports of association of cystatins with vesicular membranes/ vesicles: cystatin activity with a lysosomal membrane fraction of rabbit liver (Pontremoli et al. 1983) and stefin A (also known as cystatin-α) with keratohylin granules of rat epidermis (Takahashi et al. 1992). The apparent association of stefins A and B with hepatoma vesicular membranes in the present study would be consistent with an increased association of cystatin activity with membrane fractions of murine tumors (Rozhin et al. 1990) and with our purification of a stefin A-like cystatin from a total membrane fraction of the hepatoma (Moin et al. 1992).
Association of the cytoplasmic stefin A with vesicular membranes/vesicles intracellularly might explain how stefin A becomes associated with the cell surface. There is precedent for translocation of cytoplasmic proteins to vesicular and cell surface membranes. For example, in chromaffin cells on nicotine stimulation, annexin 2, a cytoplasmic protein, has been shown to be translocated to a membrane fraction containing chromaffin granule membrane and plasma membrane (Sagot et al. 1997). However, surface staining for stefin A was not observed in the hepatoma cells. This may be the result of a differential trafficking of stefin A in the two cell types or of cleavage of surface stefin A by proteases secreted from the Hepa cl 9 cell line. Stefin A on the surface of normal cells may serve a protective, antiproteolytic role which is absent in the tumor cell. To our knowledge, the present study is the first to demonstrate indisputably a localization of stefin A on the external surface of cells. Kolar et al. (1989) did report apical staining for stefin A in normal and neoplastic breast epithelium; however, because their study examined tissue sections, the staining visualized therein may be on the external surface or inside the apical membrane. The confocal microscopy techniques used in the present study do confirm stefin A staining on the top surfaces of cells.
We and others have postulated that alterations in the balance between endogenous cystatins and cysteine proteases may contribute to the malignant progression of tumors (Rozhin et al. 1990; for review see Kane and Gottesman 1990; Sloane 1990; Sloane et al. 1990). We have observed a reduced inhibitory activity for stefin A purified from a human sarcoma (Lah et al. 1989), an enhancement of cystatin activity in tumor cell membranes (Rozhin et al. 1990), and a stefin A-like protein in total membranes of hepatoma (Moin et al. 1992). The mechanism(s) for an association of cystatin activity and a stefin A-like protein with tumor membranes is not known. One possibility is via binding to a cysteine protease. Such interactions have been reported for tissue inhibitors of matrix metalloproteinases (TIMPs) and their target proteases, the matrix metalloproteinases (MMPs). In HT-1080 cells, a complex between proMMP-2 and TIMP-2 is necessary for binding of the MMP to the cell surface (Emmert-Buck et al. 1995).
Cathepsin B has been localized to the inner and external aspects of the bottom surface of transformed and tumor cells (Campo et al. 1994; Sloane et al. 1994b; Sameni et al. 1995; Strohmaier et al. 1997). In human colon cancer, the movement of cathepsin B to the basal membrane occurs in late adenomas/early carcinomas (Campo et al. 1994), coincident with focal degradation of the underlying basement membrane. Although stable active complexes between one inhibitor (stefin B) and a cysteine protease (cathepsin L) have been reported (Dennison et al. 1992), the present study suggests that the association of cathepsin B with the tumor cell membrane did not occur via binding to inhibitors. In fact, our results demonstrate a differential localization of the inhibitors and cathepsin B that would favor proteolysis at the bottom surface of tumor cells. Cystatin C, the most effective member of the cystatin superfamily against cathepsin B, was not associated with the bottom surface of hepatoma cells and is therefore not available to modulate the activity of cathepsin B on the cell surface adjacent to underlying matrices. The mechanism(s) by which stefin A, cystatin C, and cathepsin B becomes associated with cell surfaces and the functional consequences of cell surface stefins and cystatins and their target enzymes will require further study.
Footnotes
Acknowledgements
Supported by US Public Health Service grant CA36481. The Confocal Imaging Core is supported in part by US Public Health Service Center grants P30 ES06639 from the National Institutes of Environmental Health Sciences and P30 CA22453 from the National Cancer Institute.
We thank Dr Edith Elliott (University of Natal, South Africa) for critical review of the manuscript and Ms Grace Ziegler for technical assistance.