
Abstract
We evaluated an enzyme affinity-gold ultrastructural technique designed to identify RNA-rich structures, based on an RNase-gold (R-G) probe in human mast cells (HMCs). As expected, the R-G technique labeled RNA-containing ribosomes and nucleoli in HMCs. The heparin-rich secretory granules in HMCs were also labeled. Extensive studies revealed that HMCs isolated from lung or skin and sustained in short-term cultures, derived de novo in growth factor-supplemented cord blood cell cultures, or present in vivo in multiple sites all shared this property. We performed a large number of controls designed to examine the HMC granule binding characteristics of gold alone, of irrelevant protein- or enzyme-gold reagents, of the role of charge and enzyme activity after various enzyme digestions, after blocking with macromolecules, after exposure to inhibitors of RNase, of heparin, or to irrelevant enzyme inhibitors, including staining of macromolecule-containing test agar blocks and a variety of combined absorption and digestion experiments of the binding of R-G to HMC granules. These studies established that the R-G method detected heparin in this site in conventionally prepared, well-preserved electron microscopic samples. These findings demonstrate a new use for this enzyme affinity-gold technique in mast cell biology, based on the known property of heparin as an inhibitor of RNase.
P
Human mast cell (HMC) granules store proteoglycans that have been shown primarily to consist of heparin (and smaller quantities of chondroitin sulfate E) (Stevens et al. 1988; Thompson et al. 1988), a macromolecule said to be the most anionic substance in the body (Anderson and Wilbur 1951; Salmivirta et al. 1996) and to be unique to mast cell granules (Lindahl and Höök 1978; Salmivirta et al. 1996). Historically, the anionic charge of heparin and heparin-protein granule complexes is believed to be the basis for dye binding that facilitates the recognition of mast cells (Uvnäs et al. 1970; Lagunoff 1974; Wingren and Enerbäck 1983) and provides “false-positives” in imaging techniques based on highly positively charged avidin in light microscopic preparations (Bussolati and Gugliotta 1983; Tharp et al. 1985). In addition, this unique property of heparin has served to inhibit RNase in biochemical studies and to isolate or remove a wide variety of materials with heparin binding sites from mixed materials (Paff et al. 1947,1952; Lindahl and Höök 1978; Chirgwin et al. 1979; Jaques 1980).
We believed that these properties and the known affinity of heparin for RNase might prove useful in imaging heparin in optimally prepared electron microscopic samples. We chose to test this hypothesis in a unique subcellular site of heparin, the HMC granule. We selected this test structure because it contains a large amount of heparin and because it should not contain RNA, the known substrate for which the R-G method was developed. We found that the R-G method reliably labeled heparin in HMC granules with a degree of label specificity and visibility of ultrastructural detail equal to or surpassing existing ultrastructural methods that label mast cell granule heparin (McLaren and Pepper 1983; Craig et al. 1993; Skutelsky et al. 1995; Oliani et al. 1997). We also confirmed in HMCs the ability of the R-G method to label RNA-rich organelles (i.e., ribosomes, nucleoli) as originally reported for RPACs (Bendayan 1981). Our findings confirm and extend for a new use (e.g., to examine heparin in mast cell biology) the R-G enzyme affinity method to detect RNA (Bendayan 1981).
Materials and Methods
Materials
Sodium citrate, tetrachloroauric [III] acid, polyethylene glycol, heparin, histamine; heparinase I from Flavobacterium heparinum EC 4.2.2.7, histaminase (diamine-oxidase) EC 1.4.3.6, heparin type III agarose beads, Sepharose beads; DNA type I, DNase from bovine pancreas EC 3.1.21.1, bovine serum albumin (BSA), chondroitin sulfate (CS), proteinase K EC 3.4.21.64, pronase E from Streptomyces griseus E.C. 3.4.24.31, poly-
Source of Human Mast Cells
In vivo and in vitro sources of HMCs included biopsy specimens of human skin, breast, and lung, isolated, purified human lung or skin mast cells [some samples were maintained in short-term (6-hr) tissue cultures], and HMCs that developed de novo in culture systems from their precursors in cord blood cells. Methods of obtaining these varied sources of HMC as biopsy samples, as isolated, purified, and shortterm cultured lung or skin mast cells, and as newly emerging HMCs in culture systems have all been reported in detail (Schulman et al. 1982; Dvorak 1987; Dvorak et al. 1985, 1986,1987,1991,1992; Furitsu et al. 1989).
Fixation
Small blocks of human tissue biopsy specimens were fixed by immersion in a mixture of 2% paraformaldehyde, 2.5% glutaraldehyde, pH 7.4, in 0.1 M sodium cacodylate buffer containing 0.025% calcium chloride, for either 2 hr at room temperature (RT) (lung, breast) or 5 hr at RT (skin). Isolated, purified human lung or skin mast cells and mast cells arising in cultures from cord blood cells were fixed in suspension in a mixture of 1% paraformaldehyde, 1.25% glutaraldehyde, pH 7.4, in 0.1 M sodium cacodylate buffer containing 0.025% calcium chloride, for 1 or 2 hr at RT.
Tissue Processing
Samples were washed in 0.1 M sodium cacodylate buffer, then either postfixed in 0.2 M Sym-collidine-buffered 1.33% OsO4, pH 7.4, in osmium potassium ferrocyanide-reduced osmium tetroxide [OPF method (Dvorak 1987)] or not exposed to OsO4. Some of the 0.2 M Sym-collidine-buffered 1.33% OsO4-postfixed samples were stained with uranyl en bloc [OCUB method (Dvorak 1987)]. All samples were dehydrated in a graded series of alcohols, infiltrated in a propylene oxide-Epon 812 sequence, and embedded in Epon 812, which was polymerized at 60C for 16 hr. Thin sections were recovered on either copper, nickel, or gold uncoated 200-mesh grids and air-dried before staining.
Preparation of Enzyme-Gold Complexes
Colloidal gold suspensions (Frens 1973) were prepared as follows. Four milliliters of a 1% aqueous solution of sodium citrate were added to 100 ml of a boiling 0.01% aqueous solution of tetrachloroauric acid and boiled for 5 min. The RNase-gold (R-G) complex was prepared according to the method of Bendayan (1981). Briefly, the colloidal gold suspension was adjusted to pH 9.0 with 0.2 M K2CO3 and 10 ml of this solution was added to a siliconized tube containing 0.1-0.7 mg RNase A dissolved in 0.1 ml distilled H2O. A DNase-gold complex was prepared similarly, except that the colloidal gold suspension was adjusted to pH 6.0 before adding 10 ml of it to the siliconized tube containing 0.15-0.5 mg DNase I dissolved in 0.1 ml distilled H2O. An albumin-gold complex was also prepared as described (Bendayan 1981). BSA 0.5 mg dissolved in 0.1 ml distilled H2O was placed in a siliconized tube to which 10 ml of colloidal gold suspension, pH 7.3, was added. The three mixtures were centrifuged at 25,000 rpm for 30 min at 4C, using a Beckman 50.2 Ti rotor. The dark red sediment of each reagent (and one of colloidal gold only) was resuspended in 3 ml of PBS, pH 7.5, containing 0.2 mg/ml polyethylene glycol. A diamine-oxidase-gold complex was prepared as described (Dvorak et al. 1993b).
RNase-Gold Staining Procedure
As reported (Bendayan 1981), section-containing grids were inverted and floated section side down on a PBS drop for 5 min followed by incubation on a drop of RNase-gold for 1 hr at 37C, pH 7.5. The grids were washed sequentially in PBS and distilled water and stained with lead citrate for 3-10 min before examination in either a Philips 300 or a Philips 400 electron microscope.
Cytochemical Controls
Specificity controls [as described (Bendayan 1981)] and additional controls were done to determine the nature of HMC granule labeling with RNase-gold, as follows. Samples were stained with the uncomplexed colloidal gold suspension alone, with an irrelevant protein-gold complex (BSA-gold), with an irrelevant enzyme-gold complex (DNase-gold), or with an inactivated RNase-gold complex that had been heated at 100C for 10 min. The effect of temperature, pH, and time on HMC granule staining with RNase-gold was examined by staining at 4C, 20C, and 37C, by staining at variable pHs from 4.5 to 8.7, and by staining for 5 or 60 min. Agar blocks containing macromolecules RNA 16 mg / ml, heparin 20-30 mg/ml, poly U 10 mg/ml, poly A 20 mg/ml, chondroitin sulfate (CS) 26 mg/ml, histamine 53 mg/ml, were fixed and processed similarly to the schedule for tissue blocks or cell suspensions and were stained with enzyme-gold complexes or gold suspension alone. Exposure of the substrate in samples and the RNase-gold complex to a variety of enzymatic or acidic digestions was done. These included incubations of grid-containing sections or RNase-gold reagent with RNase 0.5-1.0 mg/ml, DNase 1 mg/ml, heparinase I 1 mg/ml, histaminase 2.2 mg/ml, proteinase K 0.25 mg/ml, pronase E 0.5 mg/ml, or 0.1 N HCl. In some cases, incubations of samples with enzymes together or in sequential order preceded staining with RNase-gold.
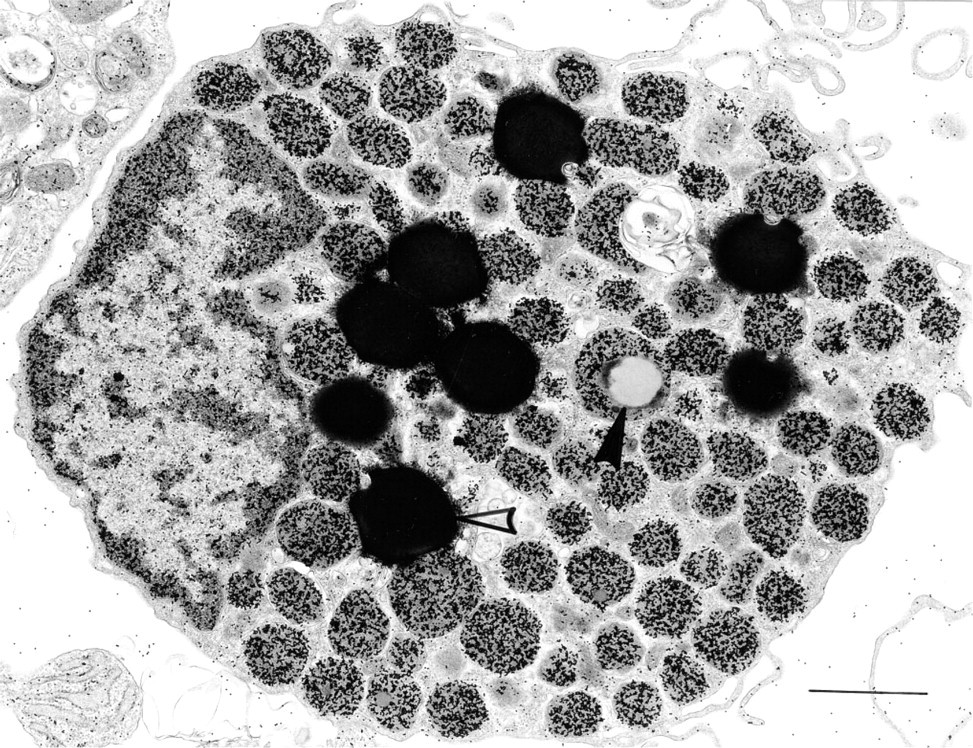
Human lung mast cell ex vivo (6-hr culture) stained with R-G shows gold-labeled cytoplasmic granules and nuclei. The large, osmiophilic cytoplasmic lipid bodies (open arrowhead) and an intragranular collection of non-osmiophilic lipid (closed arrowhead) are not labeled. Bar = 1 μm.
R-G labeling density in human mast cell granules and lipid bodies
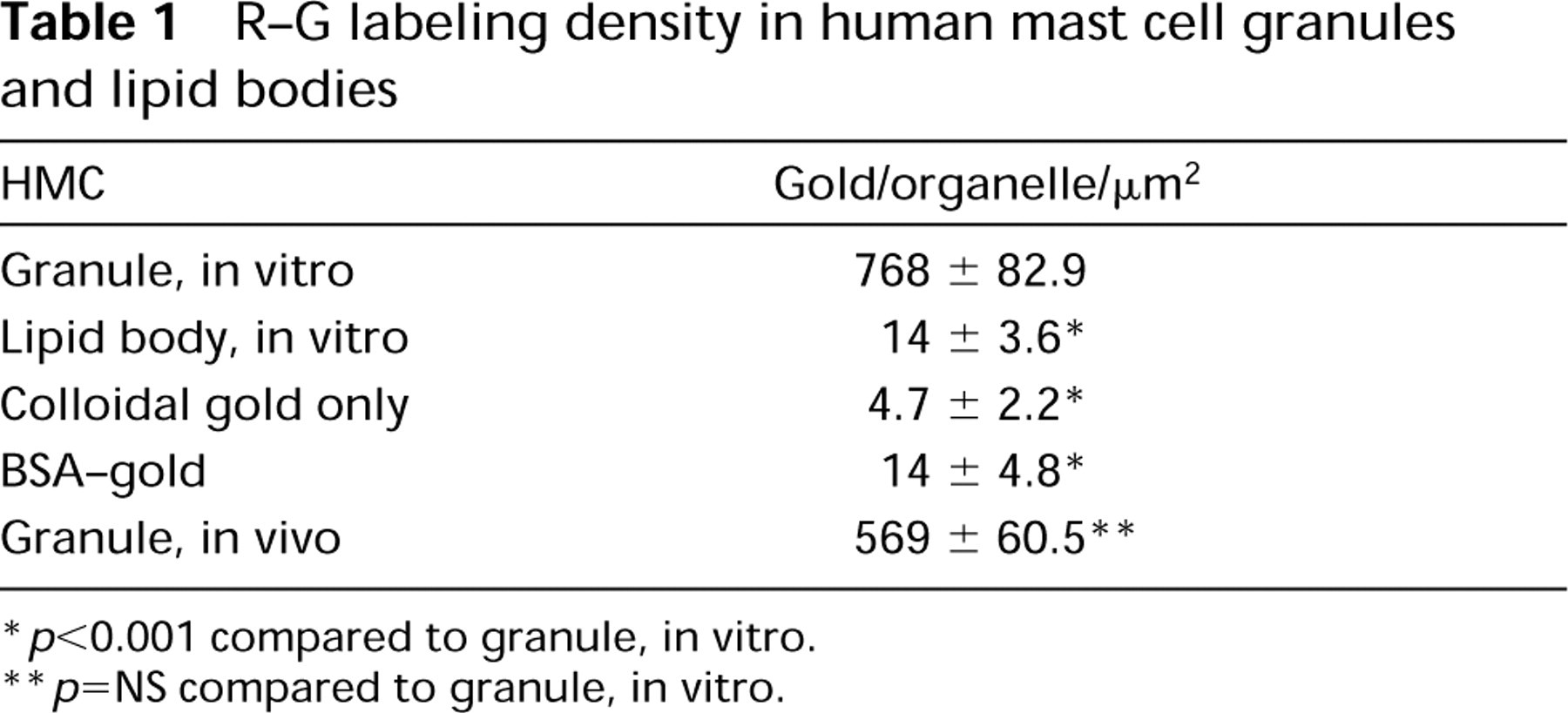
p<0.001 compared to granule, in vitro.
p=NS compared to granule, in vitro.
Cationized ferritin (CF) (0.5 ml in 10 ml Hanks' balanced salt solution) and cationized colloidal gold (poly-
Quantitation
Gold particles were counted on HMC granules of samples stained with multiple different preparations of RNase-gold as well as a variety of controls, and were expressed as the density of gold particles /μm2 of granule area. Statistical evaluations were done by the Mann-Whitney, Newman-Keuls, or Kruskal-Wallis test.
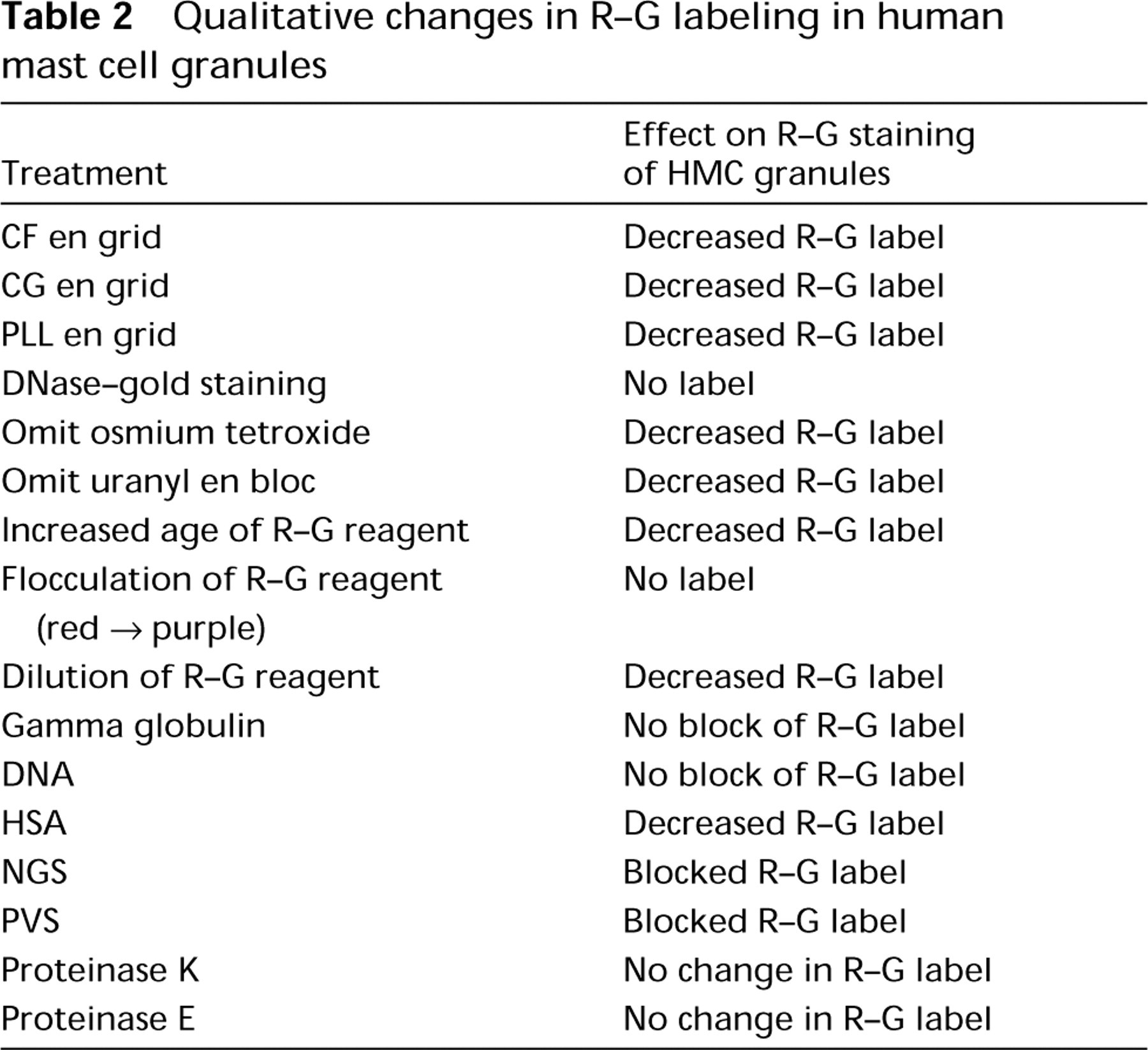
Qualitative changes in R-G labeling in human mast cell granules
Results
RNase-Gold-labeled Subcellular Sites in Human Mast Cells
HMCs stained with R-G revealed gold-labeled granules, nuclei, nucleoli, and ribosomes (Figure 1). These labeled sites were evident in HMCs of diverse origin (Table 1), including human lung mast cells (HLMCs) in vivo and after isolation, purification ex vivo, and short-term culture intervals in vitro (Figure 1) and human skin mast cells (HSMCs) in vivo and after isolation and purification ex vivo and development of HMCs in vitro from agranular precursors in human cord blood cells that were cultured with recombinant human stem cell factor (rhSCF) (Dvorak et al. 1993a).
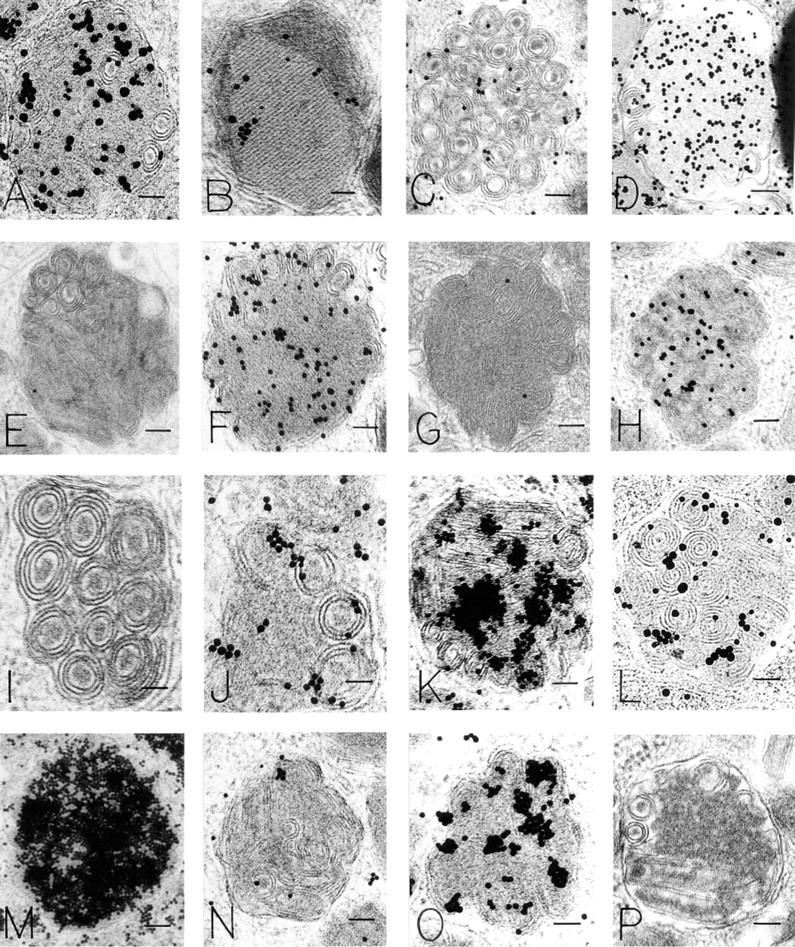
HMCs have large, electron-dense secretory granules with a variety of ultrastructural patterns. In general, these include scrolls, particles, reticular threads, crystals, homogeneous electron-dense material, and mixtures of these patterns (Dvorak 1992). All such variegated patterns in HMC granules bound R-G (mean = 452.5/μm2 HMC mature granule R-G label in 15 experiments that were quantitated) (Figures 2A and 2B). In contrast, colloidal gold alone had a mean granule label of 2.02/μm2 in six experiments (Figure 2C). An irrelevant protein-gold conjugate, BSA-gold (Figure 2D), did not label HMC granules (Table 1), nor were HMC granules stained with an irrelevant enzyme (DNase)-gold complex (Figure 2E; Table 2). Colloidal gold alone did not stain HMC granules compared to HMC granules stained with R-G (Figure 2C; Table 1). Immature granules in developing HMCs displayed electron-dense central nucleoids surrounded by a less dense matrix (Dvorak et al. 1992,1993a). In these developing granules, R-G stained the electrondense nucleoids but not the less dense matrix (data not shown). Lipid bodies, cytoplasmic subcellular organelles which are frequently observed in HLMCs (Dvorak 1991), generally were not significantly stained with R-G (Figure 2F) and served as a negative control in the same cells for granules that were stained with R-G (Table 1).
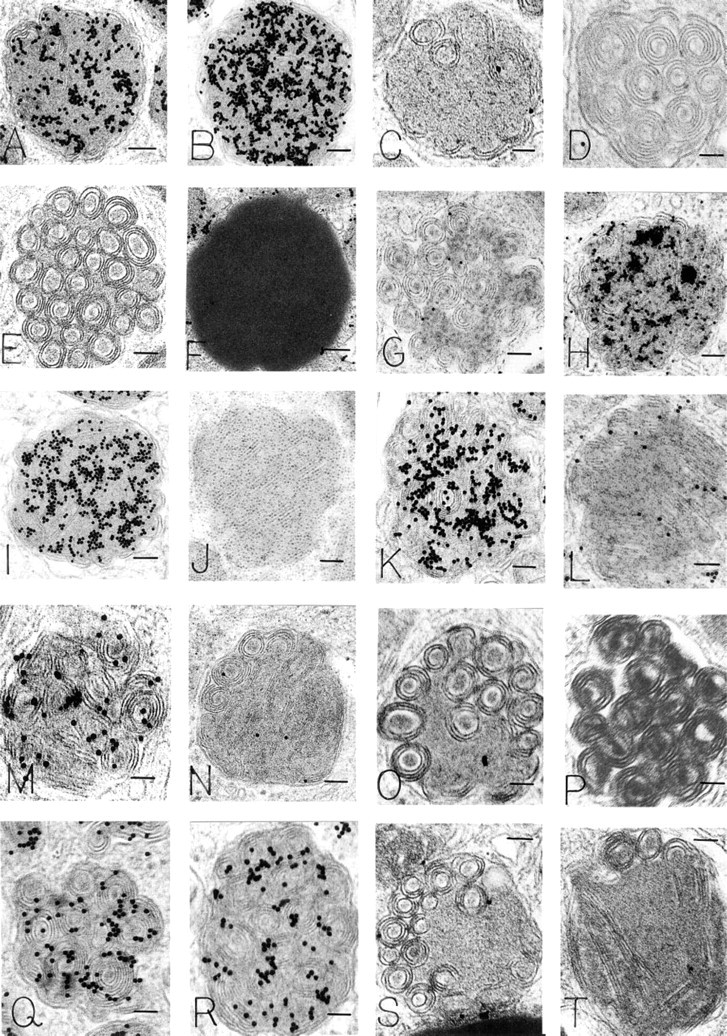
Human mast cell scroll-filled granules (all panels except
Experiments to Determine the Basis of R-G Staining of HMC Granules
Electron-dense Reagents En Grid. Two reagents were examined: cationized ferritin (CF) (Danon et al. 1972) and cationized gold (CG) (Skutelsky and Roth 1986). High-magnification views revealed that both reagents bound extensively to electron-dense substructural materials within mature HMC granules (Figure 2J). Prior staining en grid with either CF (Figures 2K and 2L) or CG, followed by R-G staining, resulted in reduction of R-G labeling of HMC granules (Table 2). CG is prepared by binding the cationic reagent poly-
Sample Preparations. When postfixation with OsO4 and staining with uranyl acetate en bloc were omitted from sample preparation of HMCs, the intensity of R-G staining was diminished but exceeded background levels for the samples.
Physical Parameters Related to the R-G Reagent. The degree of HMC granule staining was related to the age of the R-G reagent. In general, the R-G staining level for granules up to 2 weeks after preparation of the enzyme-gold complex was intense but dropped extensively after this time. Flocculation of the R-G reagent, with a color change from red to purple (Horisberger 1979), was associated with failure to stain HMC granules. Dilution of the R-G reagent up to 1:100 provided decreased stain levels that persisted, however, above background for the same samples. Heating the R-G reagent at 100C for 10 min did not alter the red color of the reagent, but HMC granule staining was eradicated (Figures 2T and 3).
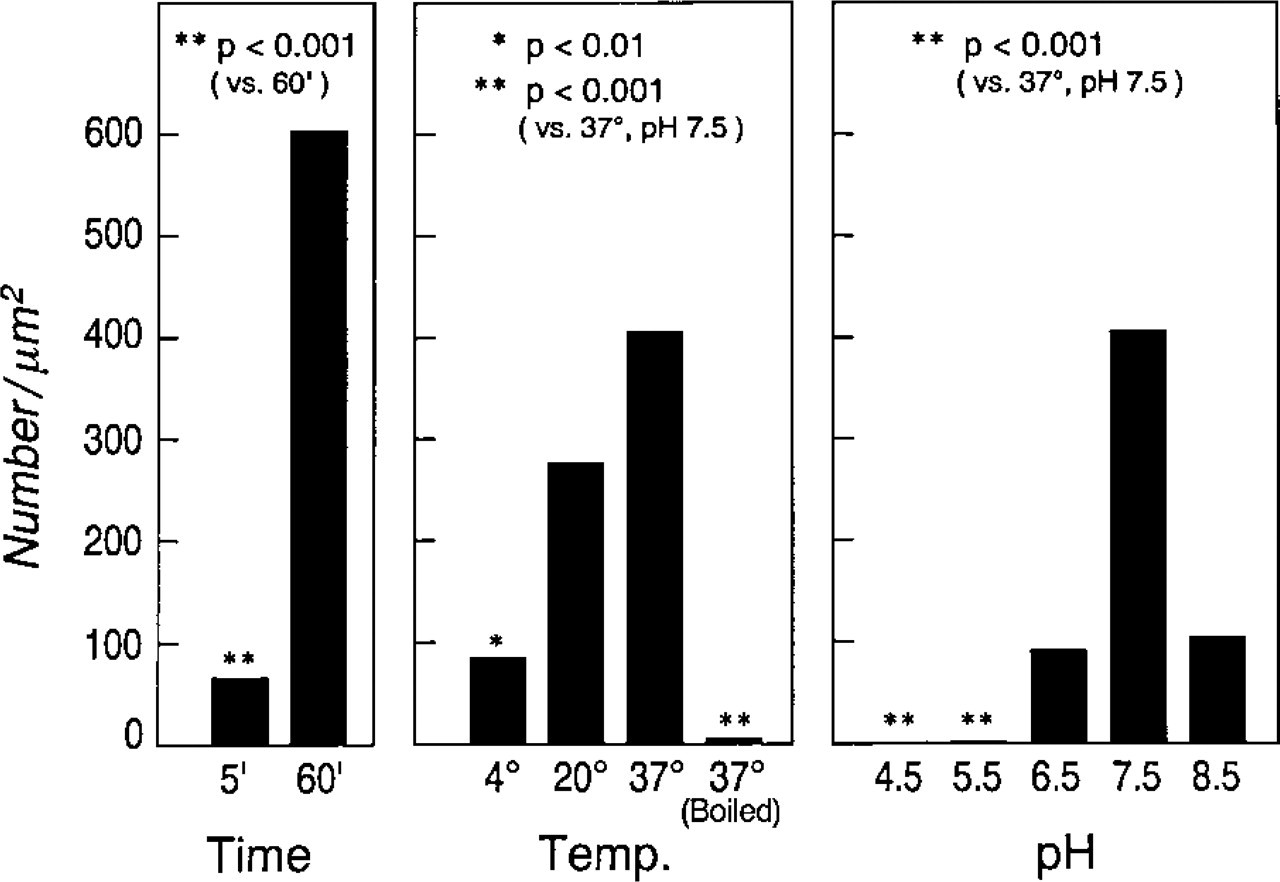
The number of gold particles/μm2 of human lung mast cell granule area representing R-G binding was established for staining time, temperature and pH. These values indicate staining for 60 min at 37C, pH 7.5, as the optimal conditions. Significantly diminished staining occurred between 5- and 60-min staining time. Significant reductions were also found when staining was done at 4C vs 37C or when the pH was 4.5 or 5.5 compared to 7.5. Boiling the RNase reagent before staining at 37C for 60 min at pH 7.5 resulted in virtual abrogation of staining.
Staining Parameters Affecting R-G Labeling of HMC Granules. The optimal time for staining grids with R-G was 1 hr and the optimal temperature was 37C. When very short staining times were used (e.g., 5 min), R-G labeling of HMC granules was evident but diminished compared to standard R-G labeling of HMC granules (Figure 3). Similarly, reductions in R-G staining were noted when temperatures of 20C and 4C were used for a 1-hr staining interval (compared to R-G staining of granules at 37C) (Figure 3). The density of R-G/μm2 of HMC granules was determined over a pH staining range spanning 8.5 to 4.5 (Figure 3). In general, the optimal pH for R-G staining of HMC granules was pH 7.5, a value used for comparative and standard purposes. Significant quantitative reductions in R-G label/μm2 granule occurred as the staining pH was increased or decreased (Figure 3).
Effect of General Inhibitors, Blockers, and Enzyme Digestions on R-G labeling of HMC Granules.
We performed experiments in which the effect on R-G granule labeling of general inhibitors, blockers, and enzyme digestions (by incubating the samples en grid in the test reagent or incubating the R-G reagent in solution with the test reagent) was assessed. In addition, a single reagent was tested before, with, or after R-G staining of samples. Furthermore, combinatorial studies of multiple reagents were done together, in sequential and reverse-sequential order. In these experiments, labeling of HMC granules was blocked and reduced, respectively, with exposure to NGS or HSA, but exposure to gamma globulin (Figure 2H) or DNA (Figure 2I) did not block R-G staining of HMC granules (Table 2). BSA placed en grid before R-G staining quantitatively reduced HMC granule label (Table 3) (Figure 2S). When BSA and R-G were incubated in solution before staining, a reduction (but not significantly so) in R-G labeling of HMC granules occurred (Figure 2R) (Table 3). Blocking with CS by incubation in solution with R-G before staining (Figure 2M) or en grid before staining with R-G qualitatively reduced but did not significantly change the R-G label of granules (Table 3). Blocking R-G with PVS abrogated HMC granule R-G staining altogether (Figure 2N; Table 2).
R-G labeling density of human mast cell granules
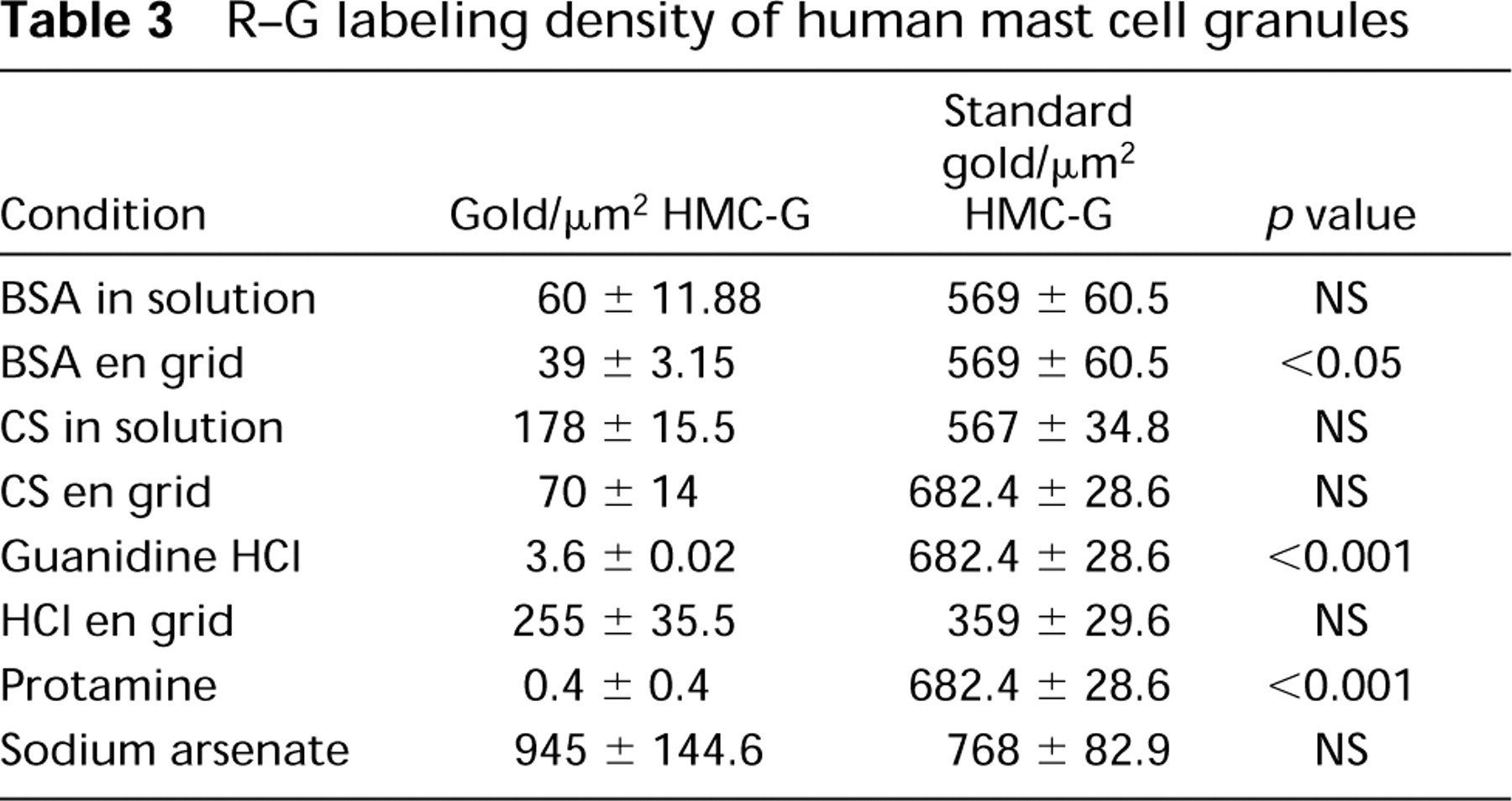
Inhibition of R-G staining of HMC granules occurred with exposure of the R-G reagent to guanidine- HCl (Figure 2O) or protamine before en grid staining (Figure 2P; Table 3); no significant change in R-G granule staining was seen with sodium arsenate (Figure 2Q; Table 3).
Digestion of samples en grid with pronase E, proteinase K, or HCl (Tables 2 and 3) did not significantly reduce R-G labeling of HMC granules.
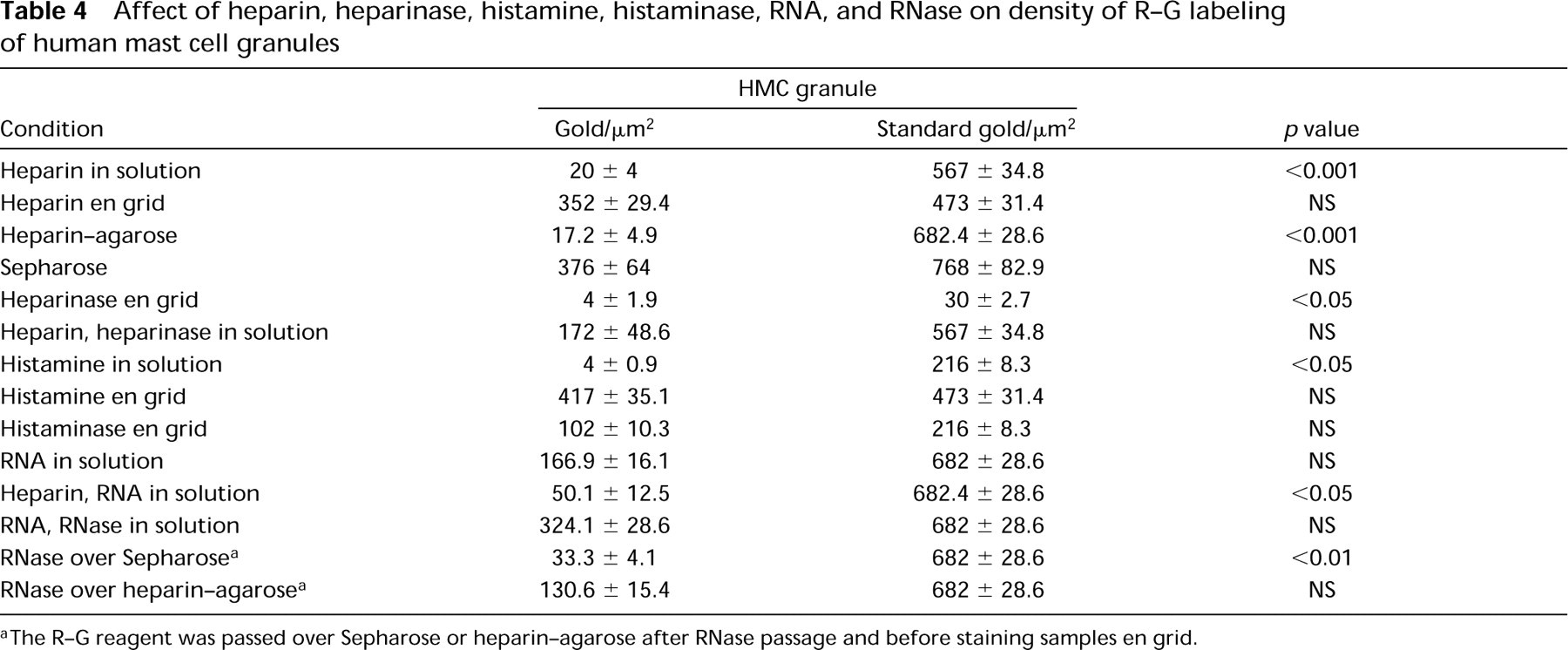
Effect of Specific Blocking and Enzyme Digestions on R-G Labeling of HMC Granules. Blocking of the R-G reagent in solution with heparin before R-G staining yielded a marked reduction in granule R-G label (Figure 4C; Table 4), but large aggregates were often present. When the heparin was placed en grid before R-G staining, no significant change in label density occurred (Figure 4D; Table 4). When the R-G reagent was absorbed by passage over solid-phase heparin before en grid staining, a marked reduction in granule staining occurred (Figure 4I; Table 4) (and the heparin-agarose turned red) compared to the retention of granule R-G staining when R-G was passed over Sepharose beads alone (Table 4) (the Sepharose beads remained white). Heparinase digestion of the grid before R-G staining resulted in diminished granule staining (Figure 4E; Table 4), whereas incubation of heparin and heparinase together with the R-G reagent before staining the grid abrogated the reduction of staining effected by heparinase (Table 4). Histamine blocking in solution resulted in decreased R-G labeling of granules (Figure 4G; Table 4), whereas histamine en grid did not affect R-G staining (Table 4). Digestion of the grid with histaminase before R-G staining did not significantly reduce HMC granule labeling with R-G (Figure 4H; Table 4).
Blocking of the R-G reagent in solution or by incubating the grid with RNA simultaneously, or before R-G incubation, yielded variable results, from a reduction in granule R-G staining that did not achieve statistical significance (Figure 4J; Table 4) to the presence of large aggregates on the grid. When RNA en grid preceded the R-G staining, no change in label density occurred (data not shown), and aggregates were excessive and random in distribution. RNase digestion of the grid before R-G staining sometimes resulted in diminished granule staining but sometimes increased it and generally produced large, random aggregates of R-G staining [as did incubation of the R-G reagent with RNase before staining the grid (Figure 4K)]. Combining digestions of grids with either proteinase or pronase before RNase digestion and R-G staining markedly increased HMC granule label (Figure 4M). Digestion of fixed HLMCs with RNase before further processing for electron microscopy and followed by R-G staining en grid resulted in no change in granule label (data not shown). Therefore, RNase digestions resulted in decreased or increased, positive as well as negative, staining in variable approaches of 10 experiments. Moreover, the presence of large aggregates impeded accurate quantitation (Figure 4K).
Affect of heparin, heparinase, histamine, histaminase, RNA, and RNase on density of R-G labeling of human mast cell granules
The R-G reagent was passed over Sepharose or heparin-agarose after RNase passage and before staining samples en grid.
Human mast cell scroll-containing (
We next evaluated a number of combined blocking and digestion experiments (Table 4). When the R-G reagent was combined in solution with heparin and RNA before staining, it resulted in a significant decrease of granule staining (Table 4) compared to standard R-G staining. However, compared to heparin blocking in solution only (Table 4) or to RNA blocking in solution only (Table 4), significant differences were not seen. When RNA and RNase were incubated in solution for 60 min before a second 60-min incubation with the R-G reagent, followed by en grid staining, random aggregates formed and the granule label was undiminished (Table 4).
In general, solid-phase heparin was most effective in absorbing the HMC granule labeling ability of the R-G reagent. We therefore incorporated its use into several combination schemes to assess the R-G reagent. When R-G was passed over Sepharose beads before staining, the granule label was retained and the beads remained white (Table 4). Passage of the R-G reagent over heparin-agarose beads before staining yielded a marked reduction of granule label and the beads turned red (Figure 4I; Table 4). Combination in solution of R-G and RNA for 1 hr, then passage over Sepharose beads and en grid staining, resulted in an insignificant reduction of gold label in granules and the beads remained white. Combination in solution of R-G and RNA for 1 hr, then passage over heparin-agarose beads and en grid staining, resulted in a significant reduction of gold granule label, and the beads were red. When R-G and heparin were incubated for 60 min in solution, passaged over Sepharose beads, and grids stained, the granule gold label was significantly reduced and the beads remained white. A double absorption with heparin, by first incubating R-G and heparin together for 60 min, followed by passage over heparin-agarose and en grid staining, resulted in the lowest density of granule gold label in the heparin absorption experiments, and the beads were stained red. We also evaluated the initial passage of RNase over either Sepharose or heparin-agarose, preceding the passage of R-G over each type of beads before en grid staining. In this case, granule R-G label after Sepharose was significantly reduced (Figure 4N) but not after passing over heparin-agarose (Figure 4O; Table 4).
R-G Staining of Agar Blocks Containing Heparin, RNA, or Histamine. Standard reagent-containing agar blocks were prepared, fixed, and processed for electron microscopy identically to the osmium collidine uranyl en bloc schedule used for the majority of cell and tissue samples. Thin sections containing these test materials were stained en grid with the R-G reagent. Agar blocks containing heparin or RNA stained with the R-G reagent. Agar alone, Epon alone, and agar blocks containing chondroitin sulfate or histamine did not bind the R-G reagent.
Discussion
We provide an examination of the properties of an enzyme affinity-gold ultrastructural technique designed to detect RNA-rich structures (Bendayan 1981,1989; Brooks and Binnington 1989) in human mast cells. These studies document a “new” use for this “old” technique.
In HMCs of wide provenance, immature and mature secretory granules known to contain heparin (Jorpes et al. 1937) were richly labeled with the R-G reagent. Mature HMCs are usually ribosome-poor (Dvorak 1992), but focal collections of free ribosomes and RNA-rich nucleoli of human mast cells also stained with the R-G method. These new experiments document an affinity of R-G for heparin in HMC granules and for RNA-containing ribosomes and nucleoli in HMC.
The findings that we present are supported by extensive controls and quantitation, which define the use of this enzyme affinity technique to image heparin in human mast cells. Heparin has been used to inhibit RNase in biochemical experiments for years (Paff et al. 1947,1952; Chirgwin et al. 1979; Jaques 1980). Therefore, it is not surprising that we have found an imageable affinity between R-G and HMC granule heparin that should prove useful in future studies of mast cell biology. Several other ultrastructural techniques for imaging mast cell heparin, based on different principles, are also available (McLaren and Pepper 1983; Craig et al. 1993; Skutelsky et al. 1995; Oliani et al. 1997).
The controls that were done in our work demonstrated that HCM granules were not labeled with gold alone, with an irrelevant protein-gold (BSA-gold), or with an irrelevant enzyme-gold (DNase-gold). An active enzyme was necessary for a positive binding reaction to HMC granules, as indicated by physical parameters that facilitated granule staining, suggesting that the binding to granules may not rely on charge alone. Charge was investigated further by directly visualizing two cationic electron-dense probes (cationized ferritin, cationic gold) bound to HMC granules that blocked subsequent R-G staining. The cationic macromolecule PLL (used to render the gold cationic) alone also blocked R-G granule staining, implicating charge in the binding reaction.
A series of enzyme or acidic digestions of samples en grid or of the R-G reagent were done. These showed specificity for heparin, because digestion of the sample with heparinase [an enzyme specific for heparin (Linker and Hovingh 1972)] removed subsequent R-G staining of HMC granules, whereas digestion with histaminase, proteinase, pronase E, and DNase did not. Digestions with RNase were technically difficult and resulted in diverse findings (see below), but when proteins were first removed from granules by proteinase or pronase E digestions, followed by RNase digestion, the residual granule heparin stained with increased intensity. Hydrolysis of the sample with HCl did not remove R-G stainable material from HMC granules. On balance, these studies supported the ability of R-G to stain HMC granule heparin.
A series of macromolecules was used to block R-G staining of human mast cell granules. Statistically significant reduction of granule label with R-G was present with heparin and polyvinyl sulfate, both known to be polyanionic competitive inhibitors of RNase (Mendelsohn and Young 1978; Chirgwin et al. 1979). CS caused a reduction in staining that did not achieve statistical significance. This trend is consistent with biochemical data documenting that the primary proteoglycan in isolated human lung mast cells is heparin but that a smaller amount of CS is also present (Stevens et al. 1988; Thompson et al. 1988). When heparinase was added to the heparin that was used to block the R-G reagent before staining, HMC granule staining was equal to standard stains, thus demonstrating that specific inactivation of heparin abrogated the heparin blocking of granule label with R-G. Neither DNA nor gamma globulin blocked R-G binding to HMC granules; RNA caused reductions that did not achieve statistical significance (see below).
Several inhibitors of RNase or heparin were examined. These showed that protamine [a heparin inhibitor that binds to heparin (Linker and Hovingh 1972; Uvnäs and Åborg 1976; Jaques 1980; Rossmann et al. 1982; Taylor and Folkman 1982)] eradicated R-G binding to human mast cell granules, as did guanidine- HCl [an inhibitor known to solubilize proteoglycans (Lindahl and Höök 1978; Hay 1981; Vogel and Peterson 1981; Rapraeger and Bernfield 1983; Carlson and Wight 1987; Thompson et al. 1988; Schick and Eras 1995)]. Incubation of the R-G reagent with sodium arsenate [a DNase inhibitor (Taper 1979)] did not reduce subsequent HMC granule binding, thus illustrating that an irrelevant enzyme inhibitor does not interfere with binding of the R-G reagent to heparin.
Histamine, a cation and a major component of HMC granules (Riley and West 1953; Lagunoff et al. 1961; Login et al. 1992; Dvorak et al. 1993b), is bound to granule heparin (Sanyal and West 1956; Kobayashi 1962; Lagunoff 1974; Jaques 1980), the most anionic substance present in mammalian cells (Anderson and Wilbur 1951; Salmivirta et al. 1996). We found that histamine successfully competed with R-G for binding to granules. Whereas heparinase treatment of the sample removed R-G granule staining, treatment with histaminase did not. Previously, we reported that another enzyme affinity-gold technique, based on the specificity of histaminase (diamineoxidase) for its substrate, histamine, specifically labels histamine in HMC granules and that prior digestion with histaminase removes the DAO-gold staining from them (Dvorak et al. 1993b).
Test blocks were prepared in agar and processed identically to the electron microscopic samples (Dvorak et al. 1993b). Blocks were thin-sectioned and subsequently stained with R-G. Blocks containing heparin or RNA bound R-G; those containing CS or histamine did not. Histamine test blocks previously were shown to bind the DAO-gold reagent (Dvorak et al. 1993b). Therefore, the R-G reagent binds to heparin in HMCs granules and to RNA, as previously reported (Bendayan 1981).
We sought to determine whether the R-G method was detecting RNA in HMC granules as well as heparin. Some of the earliest subcellular fractionation studies of another secretory granule, the pancreatic zymogen granule, reported small amounts of RNA in purified fractions (Meldolesi et al. 1971). The RNA this detected was considered a contaminant (Meldolesi et al. 1971), an interpretation that is supported by the extremely close alignment of rough endoplasmic reticulum with cytoplasmic granules in pancreatic acinar cells and the presence of some rough endoplasmic reticulum in granule fractions (Meldolesi et al. 1971). Purified preparations of HMC granules have not been reported. Therefore, biochemical composition studies of HMC granules are not available. Although an R-Gstainable component in HMC granules (attributable to RNA) is unexpected, we used a mixture of methods based on enzyme digestions, single, sequential, and combined macromolecular blocking with solid substrates to further evaluate this possibility. Before adding solid heparin-agarose to these experiments, we determined that RNA blocking in solution did not significantly reduce granule label, whereas heparin and RNA together in solution did block it, but quantitatively only to the same extent as heparin in solution alone. When we added the solid substrate heparin-agarose, we were able reliably and significantly to block HMC granule staining compared to passage of the R-G reagent over Sepharose beads alone as a control. Altogether, these experiments showed significant reduction in HMC granule label when R-G was passed over heparin-agarose before staining. Blocking of the R-G reagent with RNA, followed by passage over Sepharose, did decrease granule binding, but this did not achieve statistical significance. Either adding heparin to the RNA blocking solution or using a prestaining heparin-agarose step significantly reduced granule binding. Thus, we were unable to show an additional significant loss of R-G staining in granules that was attributable to RNA when we blocked with this macromolecule.
A number of RNase digestion experiments were done to further address the issue of RNA in HMC granules. In general, digestion of the samples on grids (or exposure of R-G to RNase) resulted in the production of aggregates of gold that were poorly quantifiable. In addition, their qualitative interpretation was often so variable as to be unhelpful. When RNase was passed over Sepharose before the R-G reagent was similarly passed over Sepharose, a significant reduction in granule staining occurred, compared to the same procedure using heparin-agarose. This, of course, could only be interpreted to mean that either heparin or both heparin and RNA are present in granules. This finding does not discriminate between these two possibilities, because RNase successfully competed with R-G binding to heparin (or RNA) in the granule in the first case. In the second case, unlabeled RNase binding to the heparin-agarose abrogated this competition, allowing the R-G reagent to stain heparin and/or RNA in granules. Altogether, these findings document the imaging of HMC granule heparin by bound RNase-gold. Other methods, including subcellular fractionation studies, will be necessary to determine whether RNA is also present in granules.
In summary, we demonstrate a new use for the ultrastructural enzyme affinity-gold method originally designed to detect RNA, i.e., to detect heparin in human mast cell granules. We also confirm that this method, as reported (Bendayan 1981), detects RNA in ribosomes and nucleoli of human mast cells. These findings provide a new use for this enzyme affinity-gold method in mast cell studies one derived from the known biochemical properties of heparin as an RNase inhibitor.
Footnotes
Acknowledgements
Supported by PHS grant AI-33372.
We thank Peter K. Gardner for editorial assistance in the preparation of the manuscript.