
Abstract
Keywords
R
In striated muscle cells the myofibrillar components constitute the bulk proportion of proteins. In heart, the half-life of mixed muscle proteins is approximately 10 days (Lewis et al. 1984). Therefore, a considerable capacity to produce proteins is required. Studies on striated muscle cells in which in situ hybridization was applied have shown that myosin mRNA are localized in the subsarcolemmal and intermyofibrillar sarcoplasm (Dix and Eisenberg 1988; Eisenberg et al. 1991). Correspondingly, ribosomes demonstrated in striated muscle cells by conventional electron microscopy (Larson et al. 1969) or immunocytochemical techniques (Horne and Hesketh 1990; Larsen et al. 1994) are mainly accumulated between the myofibrils and in the subsarcolemmal areas. These results, taken together, suggest that translation of the mRNAs of myofibrillar proteins in striated muscle cells, at least the thick filaments, occurs close to the myofibrillar sites at which the proteins are to be inserted.
Isolated cardiac myocytes from neonatal rats rapidly attain a spherical shape in suspension. In culture, these cells attach to the substrate and spread as they undergo considerable structural changes. Considerable reorganization of the normal cellular architecture, including myofibrillar disassembly and reassembly, is apparent within the first hours and days of incubation. It is generally believed that the premyofibril becomes a mature myofibril via a nascent stage by incorporation of certain muscle-specific proteins (Rhee et al. 1994). Moreover, immunolabeling of such proteins reveals a close spatial association with polyribosomes located adjacent to myofibrils (Komiyama et al. 1993). The sequence of events that takes place in the formation of myofibrils in cardiac myocytes in vitro is comparable to that seen in vivo (Borisov 1991).
In this study we examined the localization of the 60S ribosomal subunit in freshly isolated and cultured cardiac myocytes, using immunofluorescence staining techniques and immunogold electron microscopy. Redistribution of the 60S ribosomal subunit was demonstrated, along with the considerable structural reorganization of the cells during attachment and spreading.
Materials and Methods
Isolation and Culture of Cardiac Myocytes
Hearts from neonatal rats were washed in Ca++-free Joklikmodified minimal essential medium (MEM) (Gibco Life Technologies; Paisley, UK) with 26.7 mM NaHCO3, 1.20 mM MgSO4, and 1.00 mM
Primary Antibodies
Polyclonal rabbit antibodies raised against 60S ribosomal subunits, prepared from rat liver as previously described (Horne and Hesketh 1990; Hesketh et al. 1991), were kindly provided by Dr. John E. Hesketh (the Rowett Research Institute, Aberdeen, Scotland). Monoclonal antibodies recognizing α-actinin were purchased from Amersham (Poole, UK).
Western Blotting
The protein samples were separated by SDS-PAGE electrophoresis on 10% gels. Protein fractions were then electrophoretically transferred onto a nitrocellulose sheet. Nonspecific binding sites were blocked with 5% BSA (w/v) in 0.1 M PBS containing 0.1% Tween 20 (v/v) before incubation with anti-60S antibodies (diluted 1:400 with 0.8% BSA in PBS/Tween) overnight at 4C. After washing in PBS/Tween, the sheet was incubated with peroxidase-conjugated anti-rabbit antibodies (Dakopatts; Copenhagen, Denmark). The antigen-antibody-peroxidase complex was visualized using the ECL chemiluminescence detection kit (Amersham). The specificity of the primary antibodies is shown in Figure 1.
Immunofluorescence Microscopy
For immunofluorescence microscopy, cardiac cells grown on coverglasses were fixed with 3.7% formaldehyde in 0.1 M PBS after various periods of incubation. In addition, suspensions of freshly isolated cells were fixed and cytospun onto poly
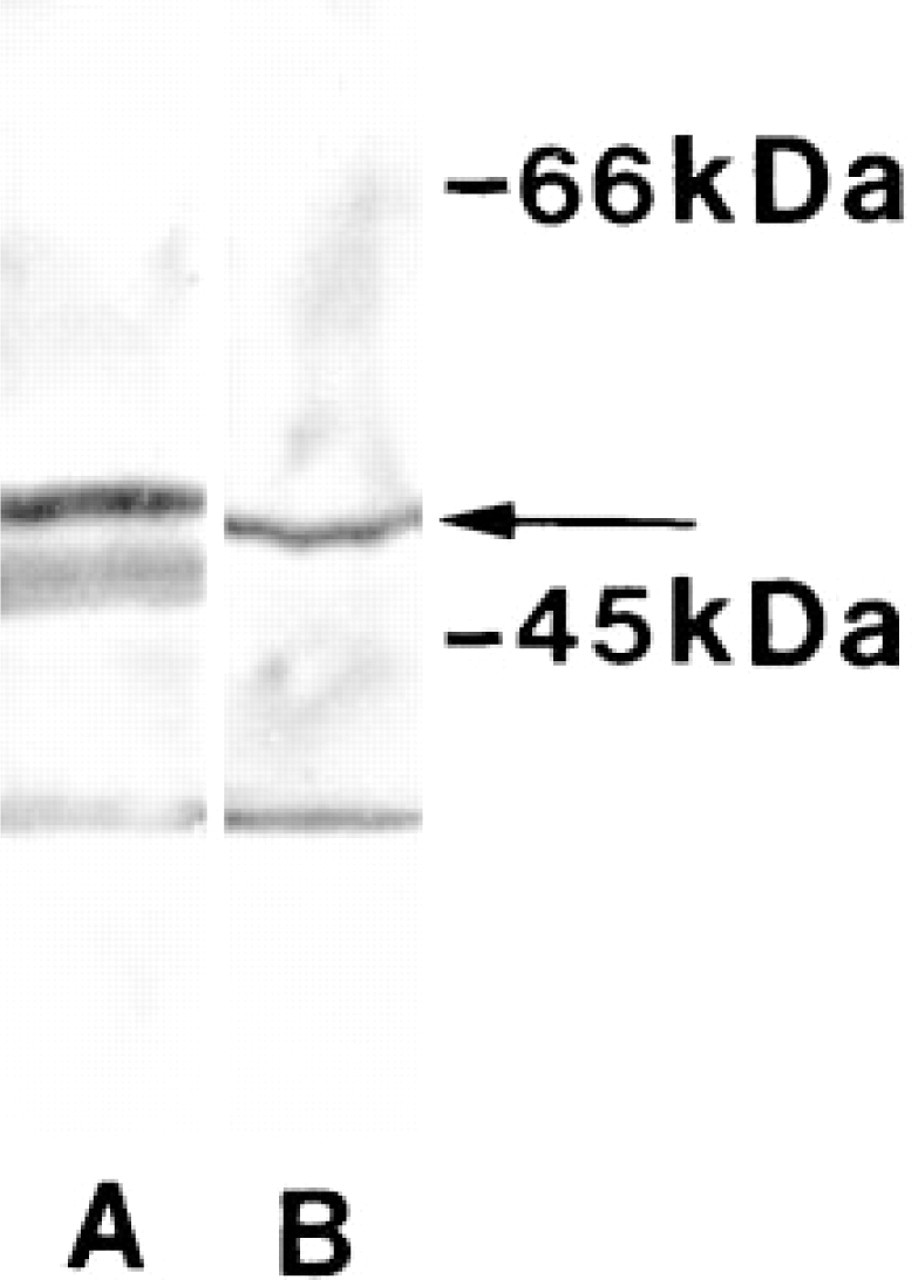
Western blot analyses of the specificity of the 60S antibodies. Protein fractions were transferred to a nitrocellulose sheet after SDS-PAGE and probed with the specific antibodies. Immunolabeling of purified 60S ribosomal subunit (Lane A) and whole-cell homogenates from cultured cardiac cells (Lane B) is demonstrated. A specific band (arrow) is identified in the cell homogenate (
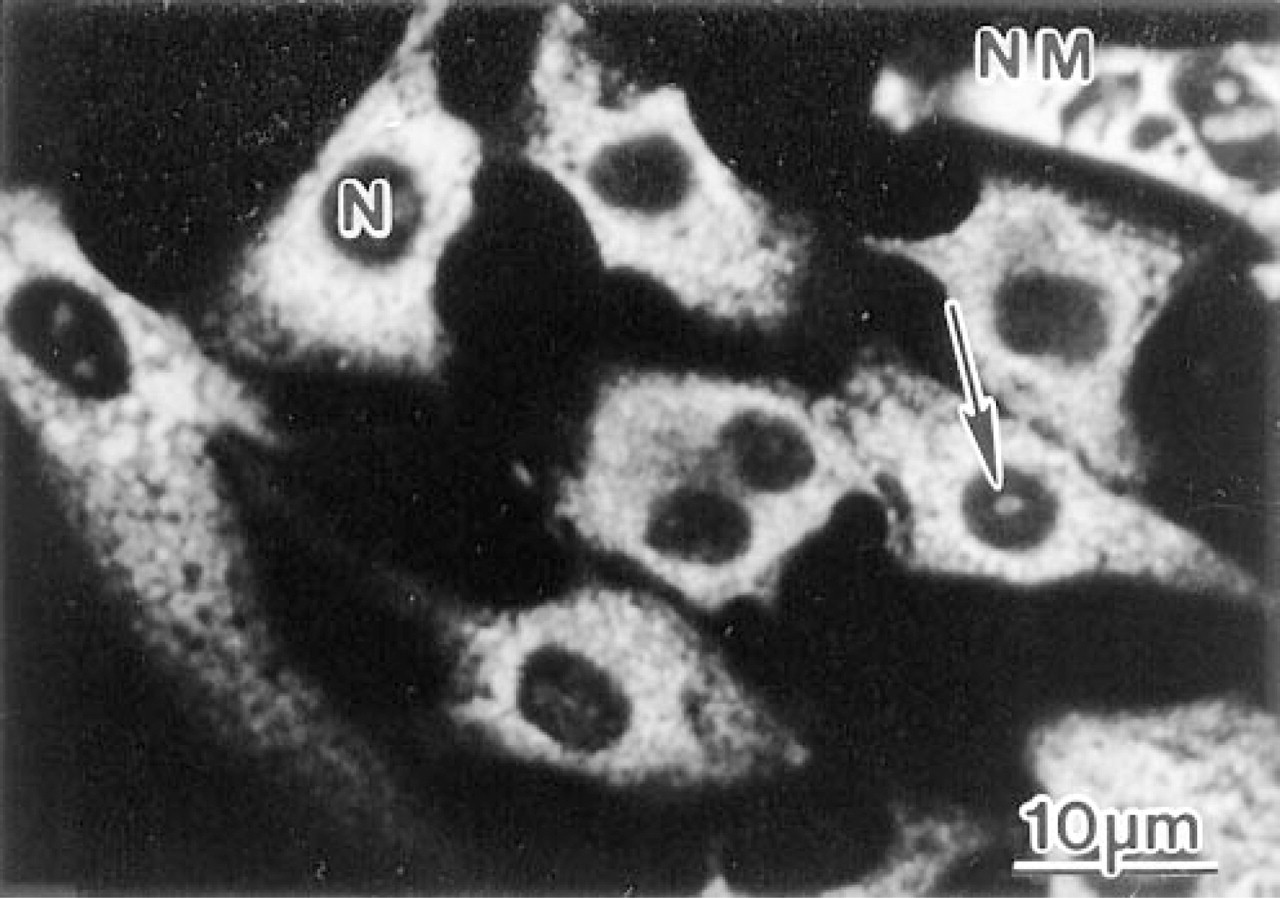
Confocal laser scanning microscopy of cardiac cells cultured for 1 day before fixation and immunostaining with anti-60S antibodies. In the noncardiac myocytes (NM) the staining is widely dispersed in the cytoplasm. In the cardiac myocytes, characterized by a centrally localized circular nucleus (occasionally they are binucleated), positive labeling is observed in the sarcoplasm. In some cells weak immunostaining of the nucleoli (arrow) occurred.
Pre-embedding Immunogold Electron Microscopy
Cultured cardiac cells were fixed in situ in the culture dish with 3.7% formaldehyde and 0.1% glutaraldehyde in Hanks' balanced salt solution (Sigma) for 1 hr at room temperature. The cells were permeabilized with 0.1% saponin for 10 min, and 0.01% saponin was used in the buffers during the incubation steps with antibodies. They were treated with glycine, preincubated with normal swine serum, and incubated with primary anti-60S antibodies using a similar protocol to that employed for immunofluorescence staining. The cells were then exposed to secondary antibodies conjugated to ultrasmall gold probes (Aurion; Wageningen, The Netherlands), postfixed with 2% glutaraldehyde, and exposed to a silver enhancement solution as previously described in detail (Larsen and Feller 1996). The cells were dehydrated in ascending concentrations of ethanol and embedded in Epon. Ultrathin sections were collected on copper grids and stained with uranyl acetate and lead citrate before being examined in the electron microscope.
Results
Confocal Laser Scanning Microscopy
After seeding, the cardiac cells became rapidly attached to the substrate and started to spread. Initially, the proportion of cardiac myocytes was 80-90%, whereas after 6 days in culture the proportion of noncardiac myocytes considerably increased. Prominent sarcoplasmic anti-60S immunostaining was revealed, and in some cardiac myocytes positive labeling was also observed in nucleoli (Figure 2). Double labeling of freshly isolated cardiac cells with anti-60S and antia-actinin antibodies showed that only a few cells retained a rod shape (Figure 3A), whereas the majority of cardiac myocytes were spherical (Figures 3B and 3C). Concomitant with spherical transformation, the sarcomeric organization of myofibrils disappeared, as demonstrated by immunolabeling with antia-actinin antibodies. During this apparently rapid disassembly of the myofibrils, immunoreactive α-actinin was redistributed to the peripherial proportion of the sarcoplasm. Immunostaining with the anti-60S antibodies revealed specific labeling in the intermyofibrillar and subsarcolemmal areas in the rod-shaped cardiac myocytes. However, in the spherical cells the staining attained a wide distribution in the sarcoplasm without any detectable organization (Figures 3B and 3C). After attachment to the substrate (4 and 8 hr), the sarcoplasmic 60S immunostaining attained a granular appearance (Figures 3D-3F). At the sites of membrane attachment, α-actinin was accumulated, whereas little 60S immunoreactivity was evident in these areas as revealed by double immunolabeling.
During further stages of culturing (12-24 hr), sarcoplasmic lamellae emerged concomitantly with the appearance of a centrally occurring sarcomeric-like α-actinin staining pattern (Figures 3G-3I). Moreover, strands of α-actinin immunostaining, extending from the centrally occurring myofibrils towards the cell surface, were co-localized with labeling of the 60S ribosomal subunit (Figures 3H and 3I). At later stages of spreading (2-6 days), the numbers of mature myofibrils increased, along with a redistribution of the 60S ribosomal subunit. In these cardiac myocytes, 60S labeling was present in the intermyofibrillar sarcoplasm (Figures 3J and 3O). Moreover, in some instances an apparent sarcomere-associated immunostaining of the 60S ribosomal subunit occurred in cardiac myocytes containing mature myofibrils (Figure 3N).
Electron Microscopy
The sarcoplasm of the spreading cardiac myocytes contained an abundance of filamentous proteins and mitochondria (Figure 4A). Filaments attached to the substrate were arranged in bundles forming myofibrillike structures (Figure 4B). Silver-enhanced ultrasmall gold particles labeling the 60S ribosomal subunit were seen widely in the sarcoplasm. The specific labeling was frequently closely associated with filaments (Figure 4C) and developing myofibrils (Figures 4B and 5).
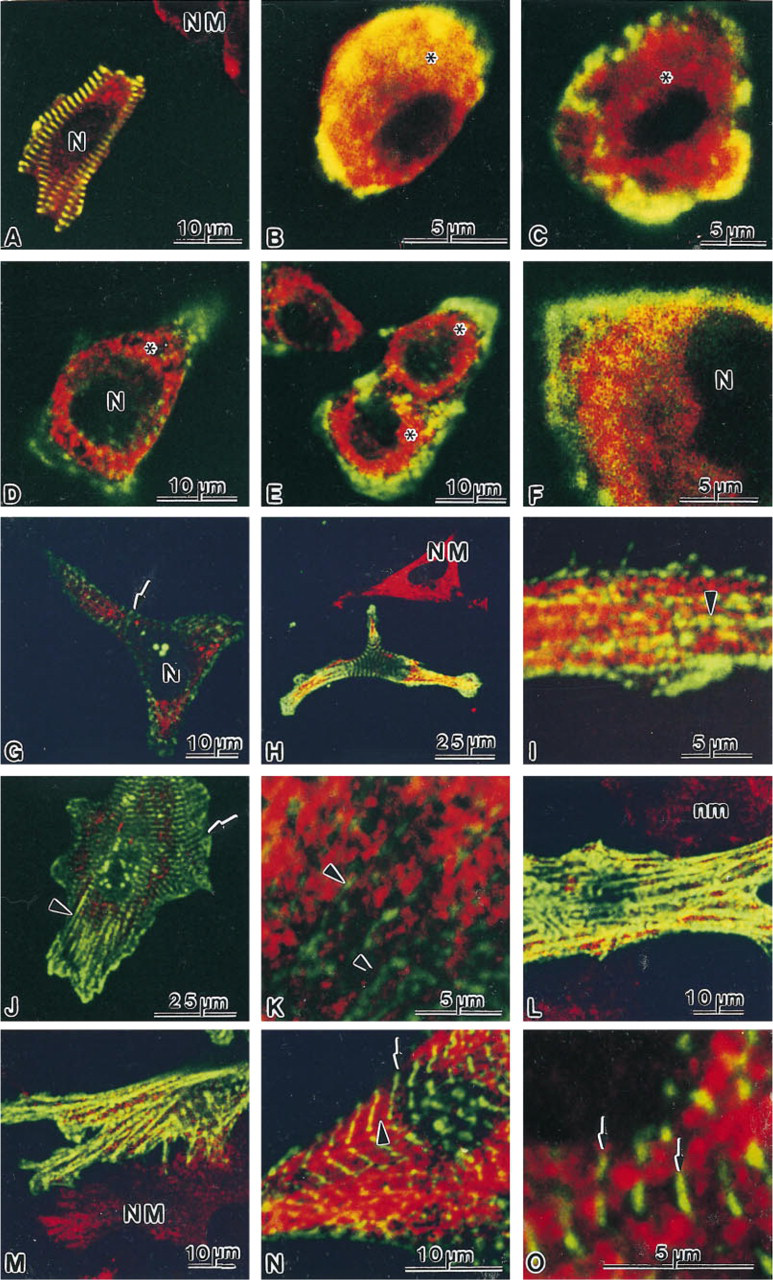
Double labeling experiments of 60S ribosomal subunit (TRITC, red fluorochrome) and α-actinin (FITC, green fluorochrome) in spreading cardiac myocytes. Overlapping of the two fluorochromes is recognized by a yellow color. After isolation, only a few rod-shaped cardiac myocytes are seen (
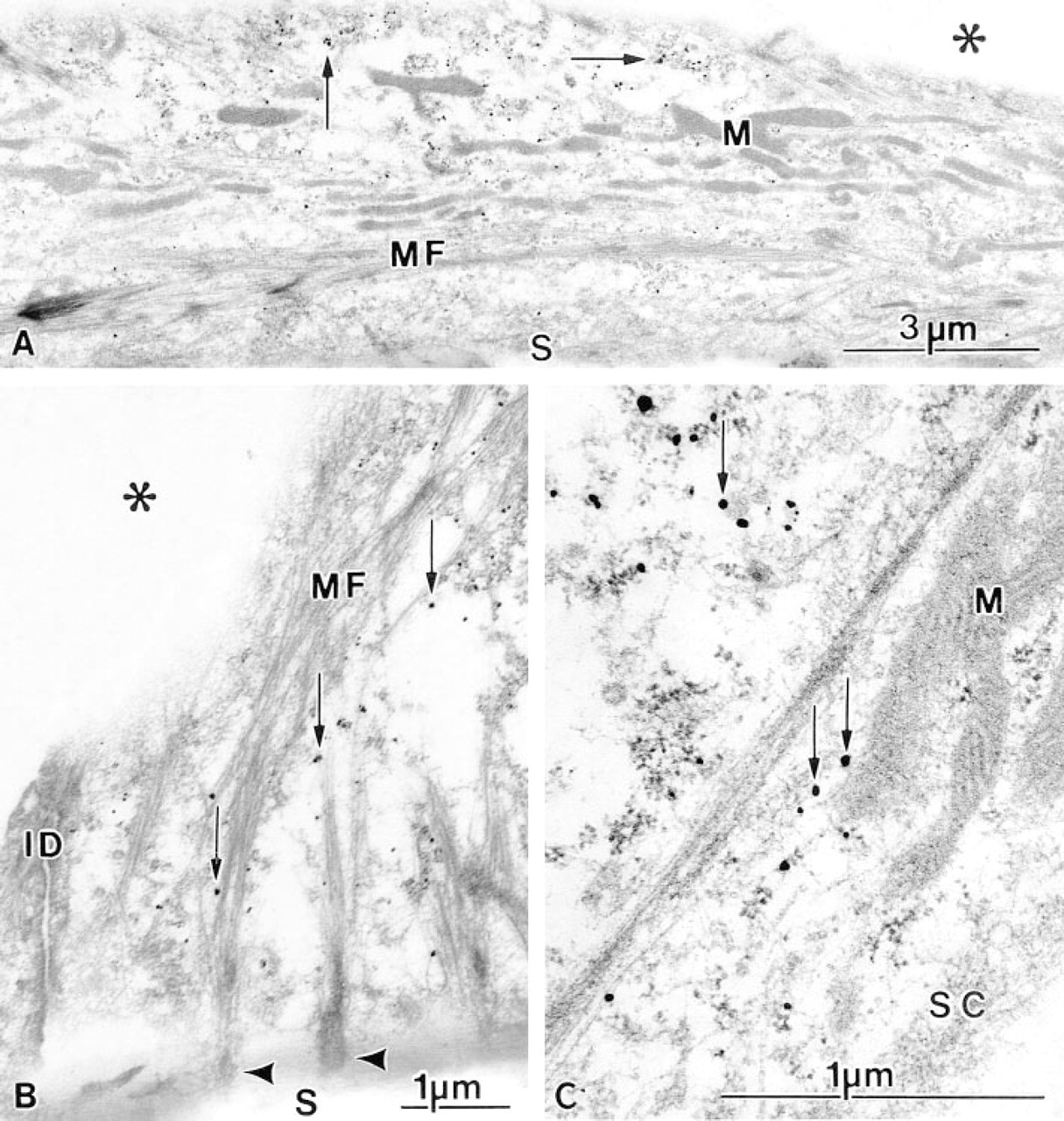
Pre-embedding immunogold electron microscopic detection of the 60S ribosomal subunit in spreading cardiac myocytes cultured for 3 days. The cells were fixed, incubated with specific antibodies, and labeled with ultrasmall gold probes followed by silver enhancement (arrows). Mitochondria (M) and myofibril-like structures (MF) are identified (
Discussion
The dramatic rearrangement of the cell architecture that occurs after isolation and attachment of the cardiac myocytes to the substrate is associated with major changes in the cytoskeleton (Guo et al. 1986). The initial redistribution of the 60S ribosomal subunit, as demonstrated in the present work, may therefore be connected with alteration of the cytoskeletal network, e.g., intermediate filaments, which coincide with the disassembly of the myofibrils. In cultures of chick cardiac myocytes it has been shown that the myofibrils disassemble through stages of transitory single sarcomeres, the cardiac polygons, that disappear after 5 days of incubation (Lin et al. 1989). During spreading of cardiac myocytes, premyofibrils are formed at the cell periphery (Lu et al. 1992). The fully mature myofibrils, however, are localized in the perinuclear areas, whereas the nascent myofibrils are positioned between the premyofibrils and the mature myofibrils (Rhee et al. 1994). During the initial stages of myofibrillogenesis, α-actinin containing dense bodies forms irregular Z-lines to which the thin actin filaments are anchored. Subsequently, muscle myosin II thick filaments as well as other sarcomeric proteins are incorporated into the preformed I-Z-I structures during maturation of the myofibrils (Schultheiss et al. 1990; Komiyama et al. 1993; Rhee et al. 1994). A similar sequence of filament incorporation occurs in the developing heart and in the adult hypertrophying heart (Sætersdal et al. 1976; Myklebust et al. 1978). In the present study, the 60S ribosomal subunit was accumulated in the perinuclear sarcoplasm during the early stages of spreading. However, after 2-3 days of culture a spatial relation between the 60S ribosomal subunit and the nascent and mature myofibrils emerged. Taking these findings together, it is indicated that the 60S ribosomal subunit has a particular role in the maintenance and growth of mature myofibrils rather than being involved in the initial formation of nascent myofirils.
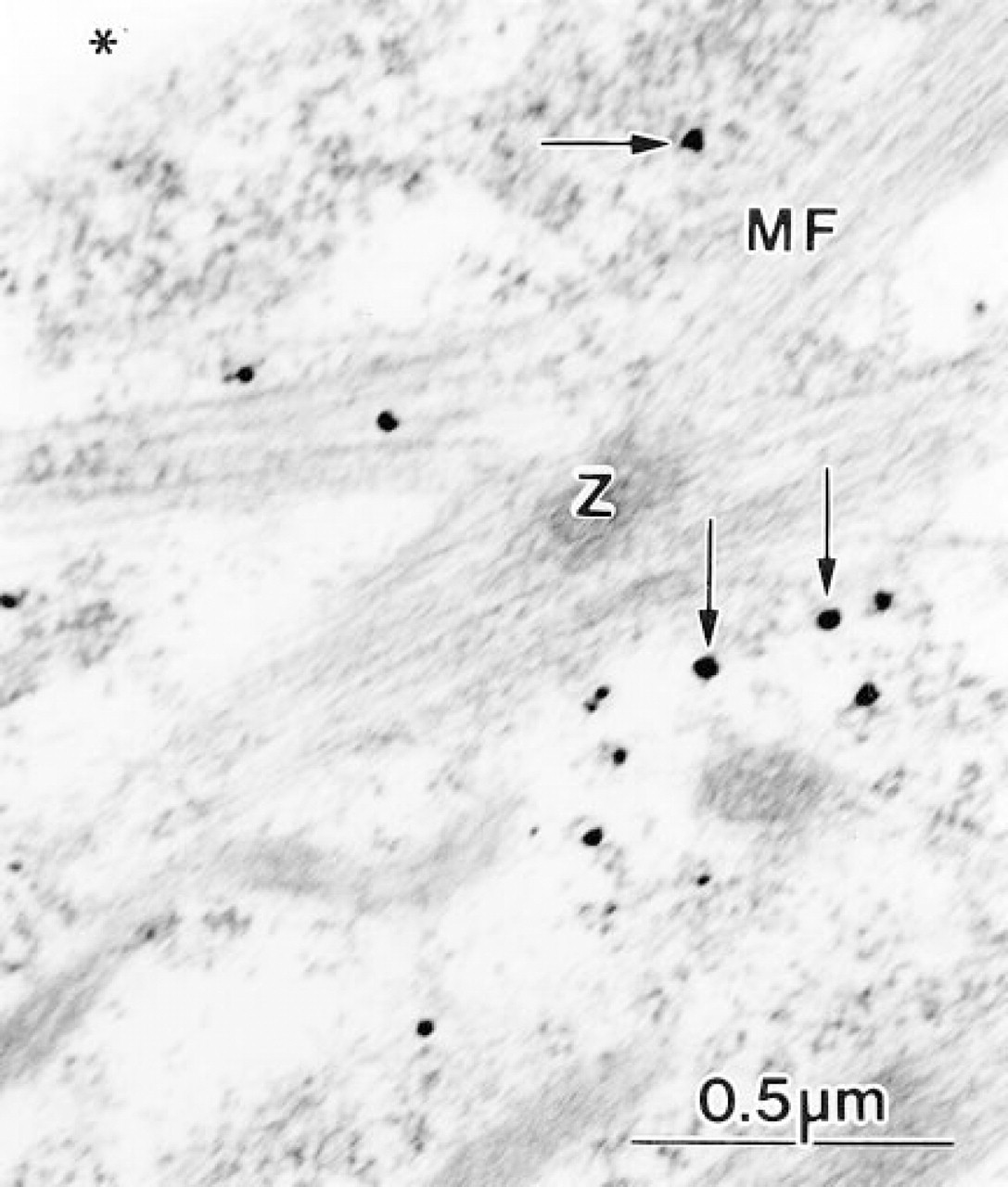
Immunolabeled cardiac myocytes incubated for 6 days. Silver-enhanced gold probes (arrows) are present in close spatial association with a developing myofibril (MF). Filaments are attached to the electron dense Z-line (Z).
A close relation of 60S ribosomal subunit staining along the myofibrils was demonstrated in cells colabeled with antia-actinin antibodies. The 60S ribosomal subunit appeared to be accumulated towards the Z-lines, which is consistent with the ribosome distribution observed in adult skeletal muscle by conventional electron microscopy (Larson et al. 1969). The synthesis of thick filaments has been shown to be connected with the presence of ribosomes (Myklebust et al. 1978). These findings are in accordance with that observed in cardiac myocytes in vivo in which the 60S immunostaining occurs as cross-striated bands in close spatial relation to the myofibrils (Larsen et al. 1994). Hence, the ribosome association to the filamentous proteins and the morphological features of cell differentiation confirm significant similarities between the developing and hypertrophying cardiac myocytes in vivo and the differentiation of isolated cardiac myocytes during culture. A large proportion of the existing pool of ribosomes are normally associated with steadystate synthesis of proteins (Luyken et al. 1996). Consequently, cardiac growth requires an increased synthesis of ribosomal proteins. Ribosome formation is regulated at the level of transcription of ribosomal RNA genes. Accelerated ribosomal RNA synthesis is accompanied by an increase of the ribosomal DNA transcription factor UBF, which is important for both initiation and maintenance of activated DNA transcription (Hannan et al. 1995,1996). Therefore, the presence of the 60S ribosomal subunit in the nucleoli of the spreading cardiac myocytes probably reflects an increased synthesis of ribosomal proteins, which are then assembled into ribosome complexes in the nucleoli before being transported back to the sarcoplasm. Similar nucleolar staining has been identified in 3T3 fibroblasts (Hesketh et al. 1991). However, preceding the increase in cell size a sequence of cell responses occurs. After induction of immediate-early genes, e.g., c-fos, the transcription of genes encoding for structural proteins and for fetal isoforms of certain proteins is increased (Lompre et al. 1991). In regionally infarcted hearts, transient c-fos activation is seen at 1-48 hr in cardiac myocytes undergoing cellular growth (manuscript in preparation). These cardiac myocytes, localized adjacent to the infarcted areas, express high levels of immunoreactive atrial natriuretic factor after 2-3 days (Larsen and Sætersdal 1993), and the size of the cells increases significantly (Cox et al. 1991). Moreover, a significantly increased content of 60S ribosomal subunits is seen in these cardiac cells after myocardial infarction (Larsen et al. 1994). Similar signaling pathways and regulation of gene transcription have been found in hypertrophying cardiac myocytes in culture as well (Sadoshima et al. 1992; Yazaki et al. 1993). In summary, the neonatal rat model is a useful system in which to study redifferentiation and myofibrillogenesis in cardiac myocytes. The present data suggest a significant role for the ribosomes during the remodeling and growth of the developing myofibrils.
Footnotes
Acknowledgements
Supported by grants from the Norwegian Research Council and the Bergen Heart Foundation.
The skillful technical assistance of Edel Karin Frotjold and Kjellfrid Haukanes is gratefully acknowledged.