
Abstract
We have developed a novel approach for in situ labeling and detection of nucleic acids in cultured cells. It is based on in vivo incorporation of chlorouridine (ClU) or iododeoxyuridine (IdU) into Chinese hamster ovary cells with the aim of labeling RNA and DNA, respectively. The halogenated nucleotides are immunolabeled on ultrathin sections with anti-bromodeoxyuridine (BrdU) monoclonal antibodies that specifically react with either IdU or ClU. Furthermore, we combined ClU and IdU incubation to label simultaneously RNA and DNA in the same cell. Both were visualized by means of anti-BrdU antibodies exhibiting strong affinity for one of the two halogenated epitopes. Confocal imaging of interphase nuclei and electron microscopic analysis showed evidence of a partial colocalization of newly synthesized DNA and RNA inside the cell nucleus. RNase and DNase digestion of ultrathin sections after formaldehyde fixation and acrylic resin embedding confirmed the specificity of incorporation. Consequently, this method allows us to differentially label DNA and RNA on the same section. Using short pulses with the precursors, we could show that newly synthesized DNA and RNA both preferentially occur within the perichromatin region at the border of condensed chromatin domains.
T
Although structural characterization of these compartments has been extensively studied during the last four decades, especially thanks to ultrastructural cytochemical studies (Monneron and Bernhard 1969), we still look for high-resolution tools, allowing one to better approach the occurrence of different functions within the architecture of the cell nucleus.
The application of halogenated precursors in ultrastructural in situ analyses of DNA replication and RNAtranscription is now well established. Cl- and I-containing deoxyuridine has been used for DNA double labeling in pulse-chase experiments (Jaunin et al. 1998). Moreover, bromodeoxyuridine (BrdU) has been shown to be suitable for labeling of chromatin domains after a long incorporation period followed by several cell cycles, giving rise to multiple chromatid segregations (Visser et al. 2000). Bromouridine (BrU) and 5-bromouridine 5′-triphosphate (Br-UTP) were especially used for short labeling of RNA and detection of transcription sites (Cmarko et al. 1999; Trentani et al. 2003).
In this study, we analyzed the location of newly replicated and transcribed molecules with regard to chromatin structure. It was achieved in Chinese hamster ovary cells (CHO) by detecting newly synthesized DNA and RNA using incorporation of different halogenated precursors by means of immunoelectron cytochemistry. This high-resolution technique for specific nucleic acid detection in ultrathin sections is based on the use of specific antibodies against bromo-deoxyuridine.
Halogenated nucleosides do not necessarily need cell permeabilization for their incorporation, and they are readily accepted by endogenous enzyme machineries and incorporated into DNA or RNA. In this work, we made use of this labeling mode to explore the possibility of visualizing newly synthesized DNA and RNA labeled simultaneously during the same incubation period. Our observations show that, using DNA and RNA precursors containing different halogenated atoms, one succeeds to specifically visualize fractions of both nucleic acids in the same cell and to analyze their possible colocalization within different architectural domains of the same nucleus.
Materials and Methods
CHO cells were grown in plastic flasks containing MEM supplemented with 10% fetal calf serum, glutamine, and penicillin/streptomycin. In the single-labeling experiments, cells in culture were incubated for 15 min with the following: 1 mM bromouridine (BrU; Sigma-Aldrich, St. Louis, MO); 50 μM chlorouridine (ClU; Biolog, Bremen, Germany); or 25 μM iododeoxyuridine (IdU; Sigma-Aldrich).
In some experiments, cells were labeled for 5 or 30 min with BrU. Moreover, a pulse-chase experiment consisting of 5-min BrU labeling followed by 30-min incubation in the absence of the halogenated precursor was performed.
In double-labeling experiments, CHO cells were incubated simultaneously with 25 μM IdU and 50 μM ClU for 15 min. In another series of experiments, cells were first prelabeled with ClU for 25 min, IdU was added to the culture medium, and the cells were further incubated in the presence of the two precursors for another 5 min.
Microinjection Experiment
In other experiments, CHO cells were microinjected with 100 mM Br-UTP (Sigma-Aldrich) and allowed to grow for 10 min.
Microinjection of Br-UTP was performed on cells cultured on microgridded Cellocate cover slips (Eppendorf; Hamburg, Germany) at 50–70% confluence, essentially as previously described (Cmarko et al. 1999). Briefly, living cells were injected into the cytoplasm with 100 mM Br-UTP in 140 mM KCl and 2 mM piperazine-N,N 9-bis(2-ethanesulfonic acid), pH 7.4. After microinjection, cells were cultured for 10 min at 37C. The injection time and the position of injected cells were recorded to determine the exact time interval between injection and fixation.
Ultrastructural Immunocytochemistry
For electron microscopy, the cells were fixed with 4% paraformaldehyde in 0.1 M Sörensen phosphate buffer, pH 7.4, for 60 min on ice, dehydrated in ethanol, and embedded into LR White resin. Ultrathin sections were collected on formvar and carbon-coated nickel grids.
Antibodies used for this study
BrdU, bromodeoxyuridine; IdU, iododeoxyuridine; Br-UTP, 5-bromouridine 5′-triphosphate; GAM, goat anti-mouse; ClU, chlorouridine; GARa, goat anti-rat.
Antibodies used for immunoelectron and immunofluorescence microscopy are listed in Table 1.
For BrU and Br-UTP detection, the sections were incubated on a drop of normal goat serum (NGS; Nordic Immunology Laboratories, Tilburg, The Netherlands) diluted 1:100 in PBS for 3 min. The incubation with monoclonal antibodies, diluted in PBS-0.05% Tween 20-0.1% BSA, was performed at 4C for 17 hr. After rinsing with PBS-Tween and PBS, the grids were incubated again with NGS as above. For mouse monoclonal antibodies, we used the secondary goat anti-mouse IgG + IgM coupled with 12-nm colloidal gold (Jackson ImmunoResearch Laboratories; Baltimore, MD), diluted 1:10 in PBS. The incubation was carried out for 30 min at room temperature.
For DNA detection, ultrathin sections were treated for DNA denaturation with 1 N HCl for 15 min at room temperature (Jaunin et al. 1998). After washing in H2O, sections were incubated on a drop of 10% NGS in PBS for 10 min and immunoreacted at room temperature for 1 hr with an anti-BrdU antibody (Becton Dickinson; Mountain View, CA) diluted 1/10 in PBS containing 1% BSA and 0.1% Tween 20. After rinsing with PBS/Tween and incubating with PBS for 15 min, grids were treated for 10 min with 10% NGS in PBS and incubated with a goat anti-mouse antibody conjugated with 12-nm colloidal gold particles (Jackson Immuno-Research Laboratories), diluted 1/10 in a solution of 1% BSA in PBS. The grids were finally rinsed with PBS and distilled water and air-dried.
For simultaneous immunodetection of IdU and ClU, preparations were processed as previously described (Jaunin et al. 1998) using an NGS/BSA blocking solution and washing with Tris high-salt buffer containing 1% Tween 20 to minimize cross-reactivity of the anti- BrdU antibodies recognizing either IdU or ClU. Sections were first incubated for 17 hr at 4C with a mixture of mouse-anti-BrdU (B+D; Becton Dickinson), which exhibits high affinity for IdU, and rat-anti BrdU (Seralab; Crawley Down, UK) that recognizes ClU. Afterward, a mixture of goat anti-mouse (GAM, 1:10; Jackson) and goat anti-rat (GARa, 1:3; Aurion, Wageningen, The Netherlands) antibodies (conjugated to 12- and 6-nm gold particles, respectively) was used for 30 min at room temperature.
All the grids were stained with the EDTA regressive technique preferential for nuclear ribonucleoproteins (Bernhard 1969) adapted for acrylic resin (Cmarko et al. 1999).
Specimens were observed with a Philips CM10 electron microscope equipped with a 40- to 50-μm objective aperture and operating at 80 kV.
Controls
To show the specificity of the labeling, several controls were performed.
Negative Controls. Some grids were floated on the incubation mixture without the primary antibody, treated as above, and incubated with the appropriate secondary antibody.
Controls of Cross-reaction. To determine the specify of the antibodies and a possible degree of cross-reactivity, the mouse monoclonal antibody, which preferentially recognizes the iodinated precursor, was incubated with ultrathin sections of cells labeled with ClU. Similarly, the rat monoclonal antibody that preferentially reacts with the chlorinated precursor was tested on the IdU-labeled cells.
RNase Digestion. To confirm the RNA nature of Cl-labeling, some sections were submitted to RNA digestion with 0.2% RNase (type IA; Sigma-Aldrich) in 1 mM triethanolamine–acetic acid buffer, pH 7.3, for 18 hr at 37C before the immunocytochemical visualization.
Specificity of halogenated precursor incorporation after 15-min labeling. (
DNase Digestion. To verify the DNA nature after IdU labeling of cells, some grids were floated on 0.1% DNase solution for 5 hr at 37C to digest DNA and used for immunoelectron microscopic assays.
Cross-digestion Controls. To further prove the high affinity of the antibodies to one of the halogens, the samples of cells labeled with IdU were incubated with the rat monoclonal antibody after DNase digestion. Moreover, sections of cells allowed to incorporate ClU were treated with RNase and incubated with the antibody that preferentially recognizes the iodinated precursor.
To further substantiate the type of nucleoplasmic RNA, some cells were treated with 10 μg/ml of α-amanitin for 12 hr to specifically inhibit RNA polymerase II. ClU was added to the culture medium for 25 min, and finally IdU was added, and the incubation continued for another 5 min.
Quantitative Evaluation
Statistical analyses were carried out on electron micrographs to determine the incorporation efficiency of halogenated precursors. Fifteen to 30 randomly selected electron micrographs from each labeling experiment were collected with a charge-coupled device camera (SIS–Soft Imaging System, Morada Camera; Olympus-SIS, Munster, Germany) at a final magnification of ×31,000. The surface of different nucleoplasmic areas was measured, and the labeling density was expressed as the number of colloidal gold grains per micrometer squared. Significance was determined using the Wilcoxon signed ranks test, and the probability level was set at p<0.05.
Simultaneous double labeling for 15 min. (
Fluorescence Microscopy
For immunofluorescence microscopic analysis, cells grown on coverslips were labeled with halogenated precursors as mentioned above. They were rinsed in PBS and fixed in 4% paraformaldehyde for 1 hr and washed in PBS, followed by 70% ethanol at −20C overnight. After rehydration in PBS, they were incubated for 1 hr, in the dark and at room temperature, with the same probes (rat anti-BrdU, 1:50; mouse anti-BrdU, 1:100) as used for electron microscopy, and revealed either with an FITC-conjugated goat anti-mouse IgG antibody (Alexa Fluor; Invitrogen, Carlsbad, CA) or a rhodamine-conjugated goat anti-rat antibody (Alexa Fluor; Invitrogen), both diluted 1:100. As controls, some samples were processed as above but with the omission of primary antibodies. Furthermore, nuclease digestion controls have also been carried out. Images were acquired with a confocal laser scanning microscope (LSM 510 Meta; Zeiss, Jena, Germany) using a ×63 oil objective.
(
Results
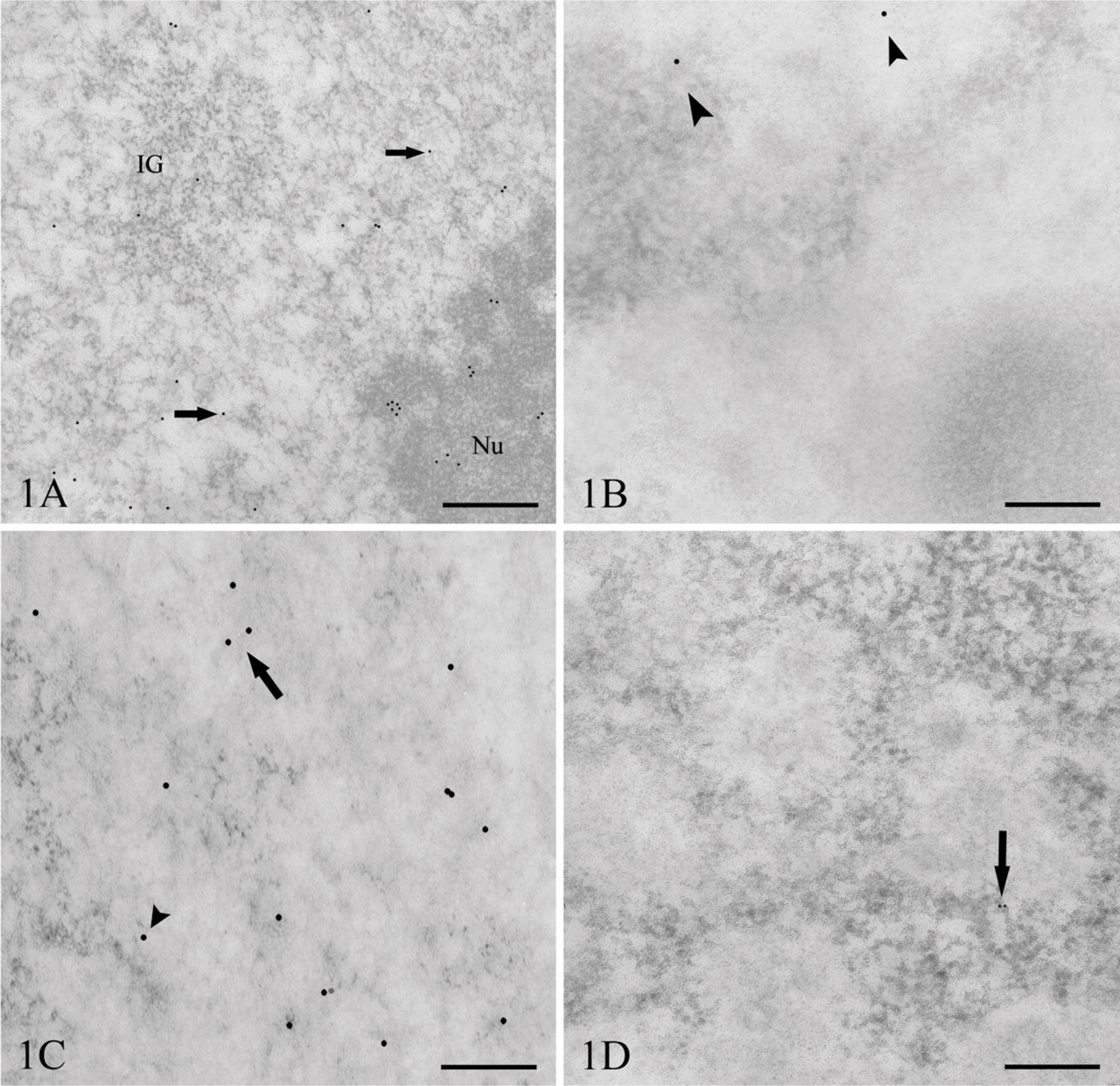
For the detection of RNA (Figures 1A and 1B), we were able to obtain a significant signal when using a rat monoclonal antibody displaying a strong affinity for ClU-labeled RNA. After 15 min of ClU incubation, the gold grains were located predominantly on the perichromatin fibrils (PFs) occurring on the border of condensed chromatin domains (Figure 1A). Some label is occasionally observed associated with the interchromatin granule clusters. The dense fibrillar component (DFC) of the nucleolus and the granular component (GC) are also labeled (Figure 1A).
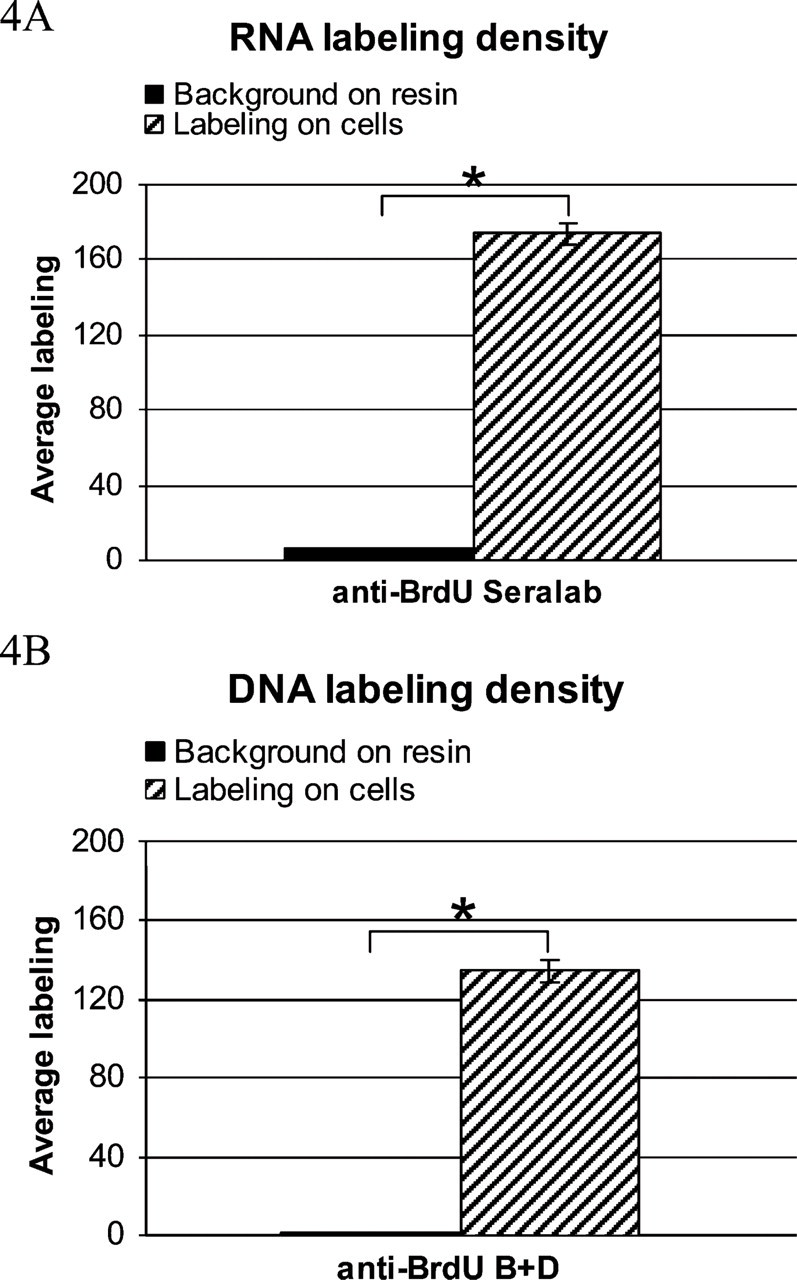
Histograms show the background on resin outside the cells. Asterisks show the significant difference between the columns. (

Background labeling on resin from a section with cells labeled for 15 min with ClU and IdU. Level of nonspecific labeling is negligible. Bar = 0.5 μm.
As for DNA detection (Figures 1C and 1D) by means of IdU, I-DNA was identified by a mouse monoclonal antibody. The signal was associated with chromatin domains located either in the proximity of the nuclear envelope (data not shown) or in the nuclear interior including the nucleolus-associated chromatin. After 15-min labeling of cells, the gold grains occurred on the periphery of condensed chromatin domains and on condensed chromatin itself (Figure 1C). Our observations showed that the signal for both the RNA and DNA is mostly represented by individual gold grains, and only a few grain clusters were detected.
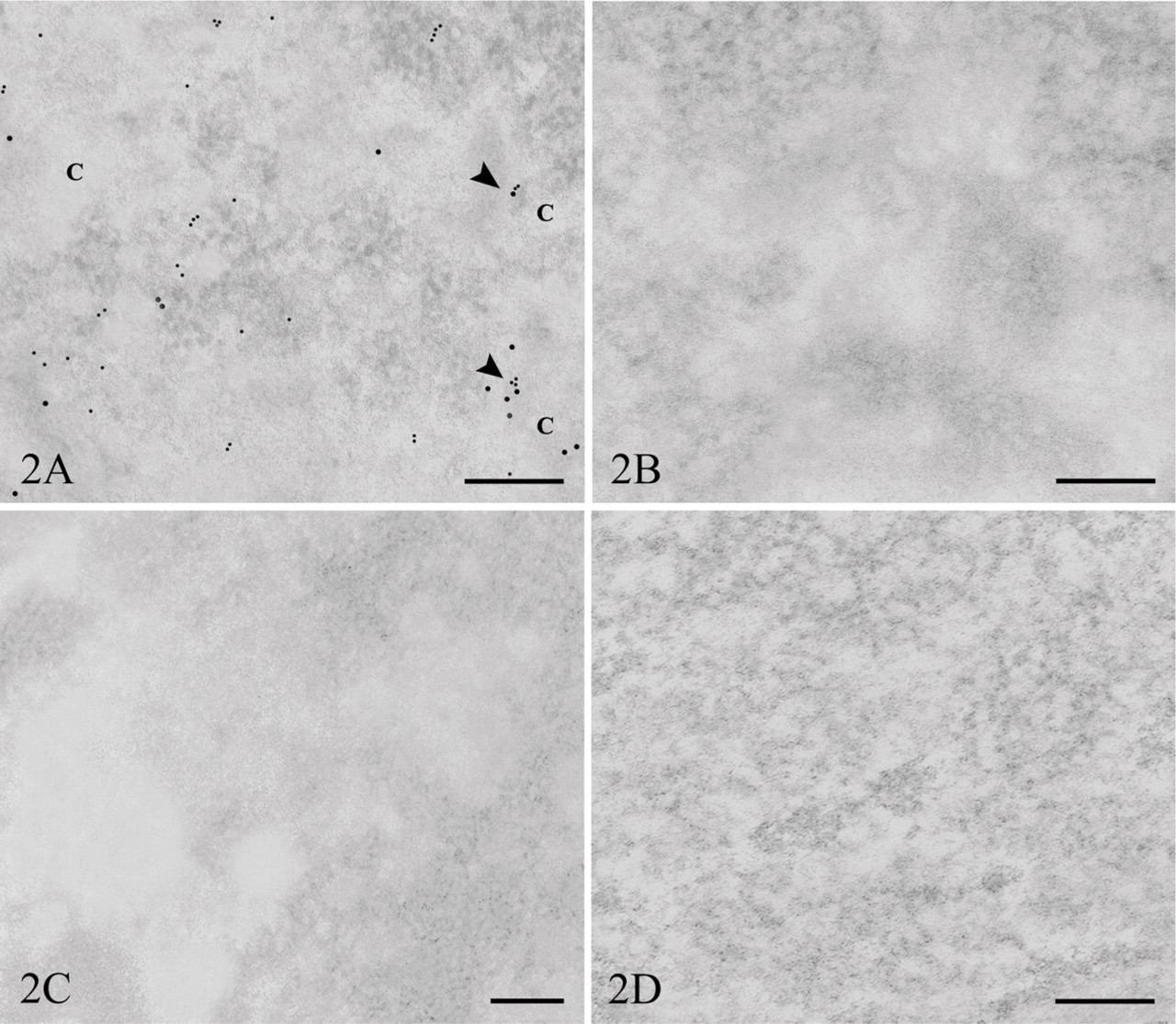
In the double-labeling experiments (Figures 2A–2D), colocalization of I-DNA and Cl-RNA can be observed (average 4.3 colocalization sites/cell nucleus) in the perichromatin region in the nucleoplasm (Figure 2A) and in the nucleolar dense fibrillar component. Moreover, HCl treatment needed for DNA denaturation does not seem to modify the intranuclear localization pattern of newly synthesized RNA.
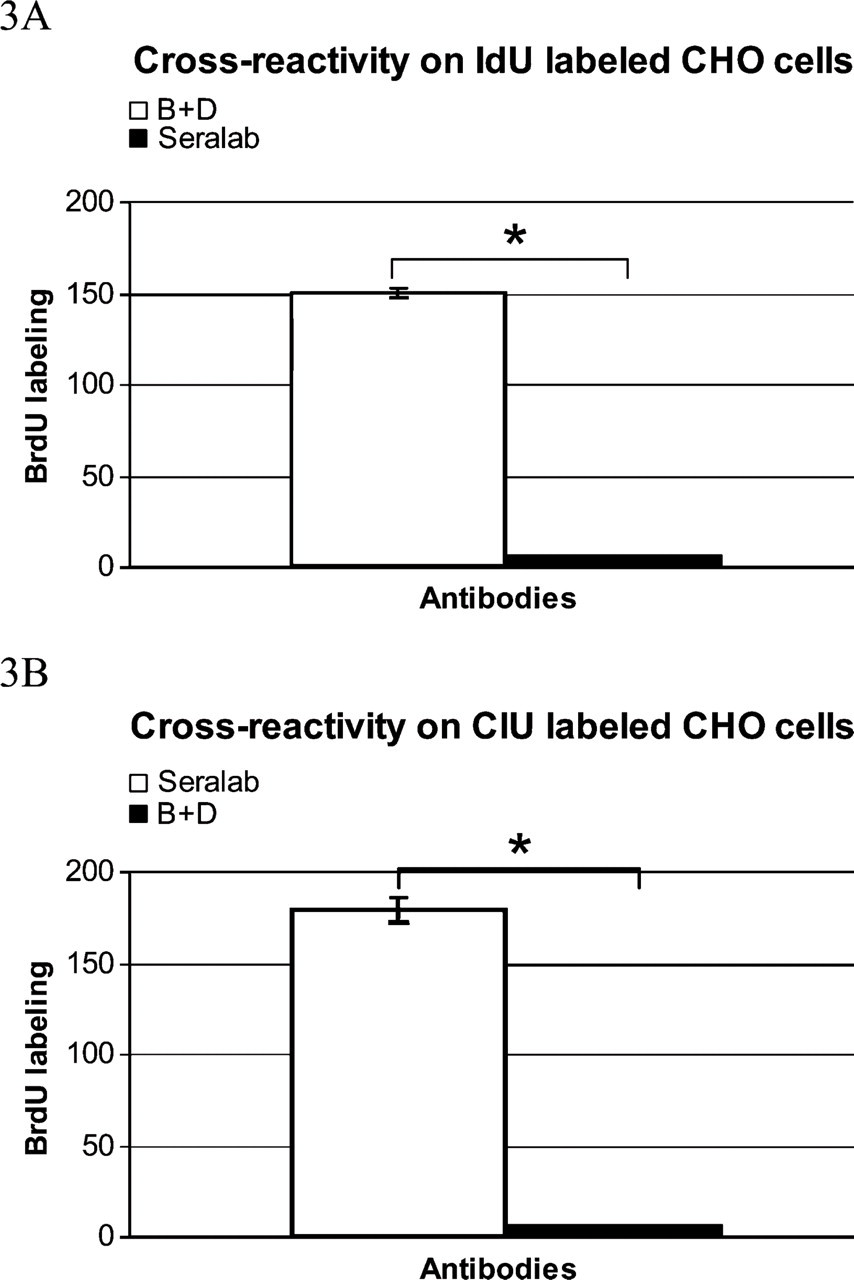
Because the anti-BrdU antibodies react with both the iodinated and the chlorinated uridine derivatives, we carried out control experiments to test the specificity of the probes used. For this aim, ultrathin sections from cells labeled with ClU were incubated with the mouse anti-BrdU that strongly binds iodinated molecules (Figure 1B), whereas a rat monoclonal antibody that preferentially reacts with chlorinated precursors was tested on the IdU-labeled cells (Figure 1D). Statistical analysis confirmed the occurrence of very low cross-reactivity for both probes: the rat monoclonal antibody exhibited a percentage of cross-reactivity corresponding to 4.45% (Figure 3A), whereas for the mouse anti-BrdU, it reached 3.34% (Figure 3B).
The background level determined on the resin outside cells was negligible for both antibodies used (Figures 4A, 4B, and Figure 5).
The tests of specificity of the DNA and RNA labeling gave rise to the following observation. Grids floated on the incubation mixture from which the primary antibodies were excluded presented a very low level of background label. When grids were pretreated with enzymes, the treatment with RNase gave rise to digestion of RNA (Figure 2C), and consequently, the loss of labeling. Similarly, DNase treatment of the thin sections of IdU-labeled cells removed the specific substrate, preventing the binding of the primary antibody (Figure 2D).
Another test of labeling specificity consisted of blocking RNA polymerase II activity responsible for most nucleoplasmic RNA synthesis by α-amanitin. Under such conditions, simultaneous double labeling with ClU and IdU gave rise to RNA inhibition and absence of ClU incorporation, whereas IdU labeling of DNA remained unaffected (Figures 6 and 7).

A double control of ClU incorporation specifity after α-amanitin treatment of cells. Cells were labeled first for 25 min with ClU and then with both ClU and IdU during the last 5 min of incubation. (
Halogenated precursor experiments also involved BrU application. After 15 min of BrU incubation, colloidal gold particles were present in the nucleoplasm, especially at the border of the condensed chromatin areas or on the PFs and on the DFC of the nucleolus (Figure 8A). Microinjection assays with Br-UTP carried out in parallel showed a comparable efficiency of incorporation and labeled RNA distribution pattern for these two brominated precursors (Figure 8B). However, ClU showed a significantly higher labeling efficiency compared with cells incubated with BrU, even though the concentration of ClU in the medium was lower.
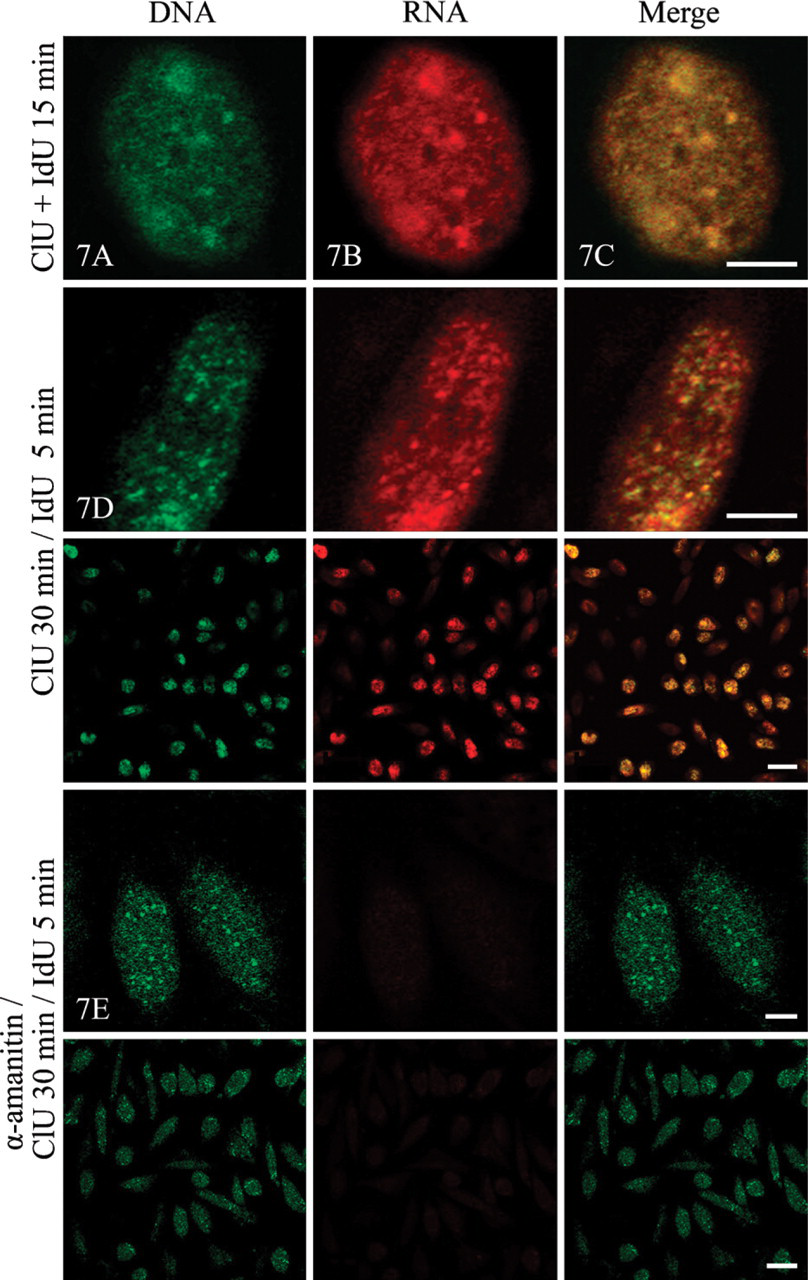
Immunofluorescence labeling of cells incubated in a mixture of ClU and IdU. (
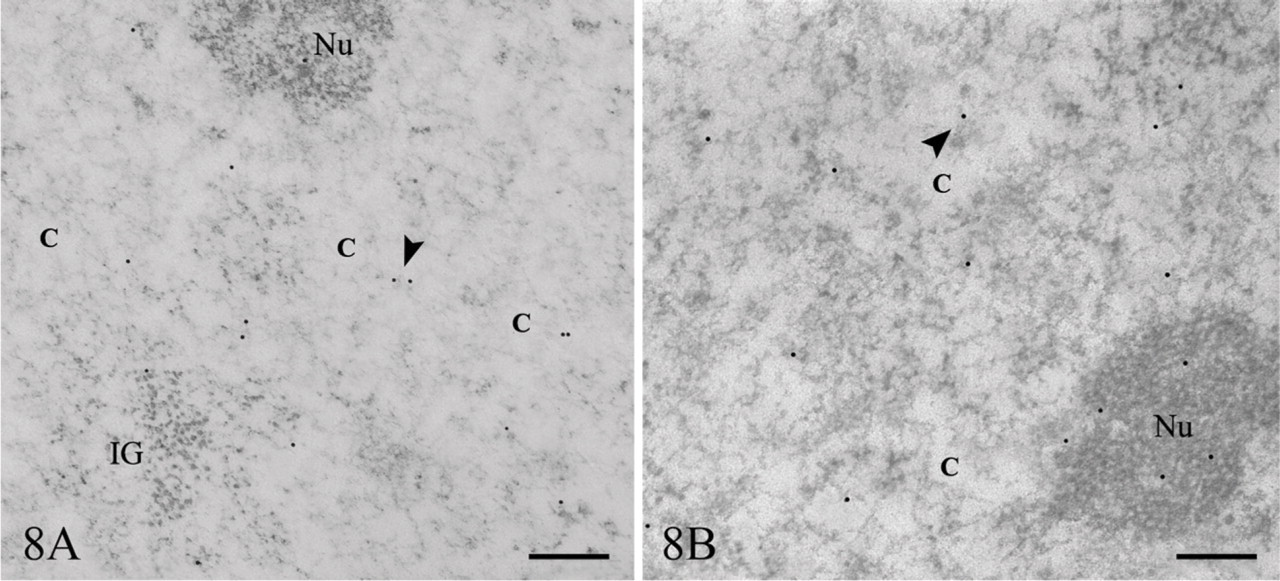
BrU and Br-UTP labeling. (
Short BrU labeling revealed a low but detectable signal in the nucleus (Figure 9A). A pulse-chase experiment (5 min + 30 min) showed some label occurring in the interchromatin space (Figure 9B), whereas after 30-min incubation with BrU, strong overall labeling in the nucleus was observed (Figure 9C).
Immunofluorescence
The confocal microscopic analysis completed the results obtained at the ultrastructural level. Optical section of a cell nucleus visualized IdU-labeled DNA after a 15-min incubation (Figure 7A). I-DNA immunodetection (green fluorescence) showed a diffuse distribution of signal interspersed with tiny spots throughout the nucleoplasm (Figure 7A). The Cl-RNA signal (Figure 7B, red fluorescence) exhibited diffuse labeling with a number of small spots, representing newly synthesized RNA. The merged image (Figure 7C) shows a high degree of colocalization (yellow fluorescence) at this level of resolution.
When α-amanitin pretreated cells were simultaneously double labeled with ClU and IdU, immunofluorescence observations clearly showed the suppression of ClU incorporation (Figure 7E), whereas IdU labeling of DNA remained unchanged compared with the cells incubated with the two precursors under control conditions (Figure 7D).
Discussion
The in vivo incorporation of halogenated precursors is a very useful way to detect and localize at high resolution in situ the sites of RNA transcription and DNA replication directly on ultrathin sections of resin embedded intact cells. Our studies showed that the addition of halogenated precursors to the medium is a valuable method for the simultaneous detection of RNA and DNA within the same cell nucleus; CHO cells show a variable yield with regard to the incorporation of precursors containing different halogens; and newly synthesized DNA and RNA do not occur in particular aggregation structures, but they are preferentially detected as individual gold grains.
The direct addition of halogenated precursors to the culture medium has shown to be a suitable technique for in situ ultrastructural RNA and DNA visualization. Although halogenated BdU has previously been widely used for detection of DNA (for review, see Jaunin and Fakan 2002), halogenated uridine directly added to the culture medium seems to be a simple and convenient way for labeling RNA in some cultured cells. Previous reports and our own experience indicated that some cell lines obviously incorporate very slowly or to a limited extent BrU from culture medium and require Br-UTP administration into cells by means of microinjection (Wansink et al. 1993; Cmarko et al. 1999), lipofection (Haukenes et al. 1997), or hypotonic shift (Koberna et al. 1999), whereas others are able to do so (Trentani et al. 2003). The possibility of a direct uptake of the halogenated precursor from the cultured medium therefore allows one to avoid any possibly invasive treatment to the cells. Furthermore, halogenated precursor detection requires the use of appropriate antibodies, keeping in mind that all halogenated nucleosides or nucleotides are to be revealed by means of anti-BrdU antibodies. The application of such antibodies can lead to an interference in the binding processes, giving rise to a cross-reaction when identifying a specific halogen-containing epitope. It is therefore essential to chose conditions where an antibody exhibits a very high affinity to a specific halogen site, thus ensuring a virtually specific detection. This can be achieved by the choice of an antibody and application of blocking agents, minimizing cross-reactivity. Our previous work showed a possibility to apply deoxyuridines containing two different halogen atoms for in situ visualizing of two different fractions of DNA in living cells (Jaunin et al. 1998). In this study, we developed a novel method allowing us to visualize a fraction of newly synthesized DNA and RNA in the same nucleus simultaneously, using a similar approach.

Pulse-chase experiment with BrU. (
Although previous studies have shown the presence of some breaks on DNA after CldU incorporation (Dillehay et al. 1984), there are no data suggesting alteration of RNA synthesis caused by ClU incorporation. Moreover, we observed a significantly higher labeling density for ClU compared with BrU (data not shown). This shows that the uptake from the medium is faster for ClU.
Although previous studies of Br-labeled RNA indicated an alteration of RNA processing events attributable to the presence of bromine atoms within the pre-mRNA molecule (Wansink et al. 1994; Fay et al. 1997), it obviously does not influence the synthetic process itself and the location of newly synthesized RNA with regard to its transcription sites (Cmarko et al. 1999). Moreover, it is interesting to note that different cell lines incorporate the BrU label with a different sensitivity. This is the case when we compare the labeling rate in CHO, HTC, and HeLa cells (unpublished data). Nevertheless, the nuclear localization exhibits the same distribution pattern in the different cell lines.
A short labeling with halogenated precursors allows one to detect the transcription sites at the periphery of condensed chromatin areas where perichromatin fibrils occur and on the DFC of the nucleolus (Hozak 1995; Cmarko et al. 1999, 2000; Koberna et al. 1999, Raska 2003).
On the contrary, a longer incubation period (e.g., 15 or 30 min) with ClU, BrU, or Br-UTP gives rise to a fraction of newly synthesized RNA located not only in the perichromatin region, but also in the interchromatin space and on both the dense fibrillar and the granular component of the nucleoli. A similar situation is observed after a pulse-chase experiment. Although we cannot estimate the percentage of halogenated substitution in our experiments, these findings may suggest some migration of halogenated RNA throughout the nucleus before its translocation to the cytoplasm, as well as between the nucleolar structural compartments.
Moreover, we noted that, after such exposure to the halogenated precursors, label for Br/Cl-RNA is mostly represented by individual gold grains without any striking accumulation of markers, suggesting that transcription mostly takes place in individual/discrete transcription sites rather than within “transcription factories” (Jackson et al. 1993, 1998; Iborra et al. 1996).
Our results point out a colocalization of signal for DNA and RNA, especially in nuclear regions where most replication and transcription occur. Fluorescence and ultrastructural observations showed a simultaneous occurrence of both newly synthesized DNA and RNA in the same nuclear region. A similar observation has been reported at the light microscopic level, combining biotin and Br-labeled precursors microinjected into cells (Hassan et al. 1994). Considering that after a 15-min incubation, at least a part of our labeling signal still represents replication and transcription sites, one can reasonably assume that colocalization of halogenated DNA and RNA in the perichromatin region can also include genes that could be engaged simultaneously in the replication and transcription process. Indeed, direct visualization of some genes during Drosophila embryo development showed this phenomenon after spreading of active chromatin by the Miller procedure (McKnight et al. 1977). In addition, it was shown in yeast that activators of transcription were involved in stimulating DNA replication by recruiting the RNA polymerase II transcription complex to DNA replication sites (Stagljar et al. 1999).
In conclusion, the halogenated precursor approach allows visualization, in the same nucleus, of the sites of both DNA replication and RNA elongation. The possibility of using chlorinated uridine in combination with iodinated deoxyuridine opens a new way of studying the intranuclear pathways of DNA and RNA in the nucleus in situ and to further approach the structural association of important functional processes in the cell.
Footnotes
Acknowledgements
This work was supported by the Swiss National Science Foundation (Grant 3100AO-109333/1) and Fondo di Ateneo per la Ricerca (FAR 2006).
The authors thank J. Fakan and F. Voinesco for excellent technical assistance and W. Blanchard and A. Spedito for photographic work. They are indebted to Dr. J. Rouquette for the critical reading of the manuscript.