
Abstract
Hepatocellular Na+,K+-ATPase is an important driving force for bile secretion and has been localized to the basolateral plasma membrane domain. Cholestasis or impaired bile flow is known to modulate the expression, domain specificity, and activity of various transport systems involved in bile secretion. This study examined Na+,K+-ATPase after ethinylestradiol (EE) treatment and after bile duct ligation (BDL), two rat models of cholestasis. It applied quantitative immunoblotting, biochemical and cytochemical determination of enzyme activity, and immunocytochemistry to the same livers. The data showed a good correlation among the results of the different methods. Neither EE nor BDL induced alterations in the subcellular distribution of Na+,K+-ATPase, which was found in the basolateral but not in the canalicular (apical) plasma membrane domain. Protein expression and enzyme activity showed a small (~10%) decrease after EE treatment and a similar increase after BDL. These modest changes could not be detected by immunofluorescence, immuno EM, or cytochemistry. The data, therefore, demonstrate that Na+,K+-ATPase is only slightly affected by EE and BDL. They suggest that other components of the bile secretory apparatus that take effect downstream of the primary basolateral driving force may play a more prominent role in the pathogenesis of cholestasis.
Keywords
I
In hepatocytes, a multitude of transport systems in the plasma membrane are involved in the regulation of intracellular ion homeostasis, membrane potential, intracellular pH, and ion transport (Graf and Häus-singer 1996). Many of these carriers are dependent on an Na+ gradient and therefore are coupled to Na+, K+-ATPase.
Whereas functionally active Na+,K+-ATPase is localized in the basolateral domain of most polarized epithelial cells, controversial immunochemical data have been reported on the distribution of the holoenzyme and its subunits in hepatocytes. Only recently has a consensus emerged that functionally competent α/β-heterodimers are expressed in the basolateral plasma membrane domain only (Simon et al. 1995).
Cholestasis is a term for pathophysiological conditions that are characterized by reduced bile flow. Cholestasis has been shown to be associated with altered expression and/or activity of various transport systems (Bossard et al. 1993; Gartung et al. 1996; Simon et al. 1996) and with relocation of domain-specific plasma membrane proteins (Stieger et al. 1994). Few data, mostly on enzyme activity, have been published on Na+,K+-ATPase in cholestatic liver. They indicate that ethinylestradiol (EE) induces a decrease (Bossard et al. 1993; Simon et al. 1996), whereas bile duct ligation (BDL) does not result in appreciable changes (Toda et al. 1978).
Therefore, this study examined the expression, activity, and distribution of Na+,K+-ATPase in two animal models of cholestasis; EE for intrahepatic and BDL for extrahepatic obstructive cholestasis. We applied quantitative immunoblotting, biochemical and cytochemical determination of enzyme activity, and immunocytochemistry to the same rat livers. The data demonstrate that neither EE nor BDL induces alterations in the subcellular distribution of Na+,K+-ATPase, and that protein mass and enzyme activity are only slightly altered.
Materials and Methods
Reagents
17α-Ethinylestradiol was purchased from Sigma Chemical (St Louis, MO), pentobarbital (Nembutal) from Abbot Laboratories (North Chicago, IL), 1,2-propanediol from Fluka AG (Buchs, Switzerland), and the kit for determination of bile acids from Becton Dickinson Diagnostics (Orangeburg, NY). All other reagents were of the highest purity available from either Merck (Darmstadt, Germany), Fluka, or Sigma.
Animals and Treatments
Cholestasis was induced in male Sprague–Dawley rats (240–275 g; BRL, Füllinsdorf, Switzerland) as described (Rahner et al. 1996) by EE and BDL, respectively. Controls included treatment with solvent (propanediol) alone or sham operation.
Cholestatic Parameters. The degree of cholestasis was monitored as described (Rahner et al. 1996). All experimental animals showed clear signs of cholestasis (data not shown).
Preparation of Livers
Animals were anesthetized and livers flushed free of blood as described (Rahner et al. 1996). After tying the hila, the right caudal and lateral lobes were excised and homogenized immediately for biochemical examination (see below). The remainder of the liver was fixed by perfusion for 5 min at room temperature (RT) with 4% (w/v) paraformaldehyde, 0.1% (v/v) glutaraldehyde, 2% (w/v) sucrose in 0.1 M Pipes (piperazine-N,N'-bis(2-ethanesulfonic acid)), adjusted to pH 7.4. After ligation and excision of the left lobe for immunofluorescence and electron microscopy, perfusion of the median lobe was continued for an additional 5 min with 0.25% glutaraldehyde (v/v) in the same buffer for cytochemistry.
Western Blot Analysis
To concentrate the homogenate, aliquots were centrifuged at 100,000 X g for 1 hr and resuspended in the buffer used for homogenization. After protein determination, equal amounts of protein of this total particulate fraction were separated by SDS-PAGE and further processed by Western blotting (Stieger et al. 1994). The nitrocellulose filters were probed with a monospecific polyclonal antiserum against the α1-sub-unit of Na+,K+-ATPase (Upstate Biotechnology; Lake Placid, NY) at a dilution of 1:1000 and antibody was visualized with iodinated protein A. In preliminary experiments, linear responses to the amount of protein and to various exposure times of the film were established (data not shown). The autoradiographs were densitometrically quantitated with a CAMAG TLC scanner II (CAMAG AG; Muttenz, Switzerland).
Determination of Enzyme Activities in Homogenates
Immediately after removal the right caudal and lateral lobes were homogenized (20% w/v) as described (Stieger et al. 1994). The homogenate was filtered through one layer of cheesecloth and was subsequently used to determine the activity of aminopeptidase N (Haase et al. 1978) and of Na+,K+-ATPase (Berner and Kinne 1976) at 37C in an LKB 2086 (Bromma, Sweden) reaction rate analyzer. For calculation of specific activities protein was measured with a modified Lowry procedure (Bensadoun and Weinstein 1976).
Cytochemistry
Fifty-μm tissue slices were collected in 100 mM Pipes buffer, pH 7.4. Na+,K+-ATPase activity was shown using the cerium technique, as described (Angermüller et al. 1995). Controls were carried out in incubation media without substrate and in preincubation and incubation mediums containing substrate and 2.5 mM ouabain, a specific inhibitor of Na+, K+-ATPase. All sections were postfixed with reduced osmium containing 1% aqueous osmium tetroxide and 1.5% potassium ferrocyanide, dehydrated in graded ethanol, and embedded in Epon 812. Ultrathin sections were examined without counterstaining.
Immunofluorescence
Cryosections of 0.5-1 μm were reacted for indirect immunofluorescence as described (Stieger et al. 1994). The primary antibody (monoclonal antibody IEC 1/48 against the β-subunit) (Marxer et al. 1989) was diluted 1:50 (v/v) and applied for 30 min. Incubation with affinity-purified FITC-labeled goat anti-mouse IgG (Jackson ImmunoResearch Laboratories; West Grove, PA) diluted 1:100 (v/v) lasted for 20 min. Controls included substitution of the primary antibody by PBS.
Immunoelectron Microscopy
Ultrathin cryosections were picked up with 2.3 M sucrose and stored on drops of PBS containing 0.5% (w/v) BSA and 0.1% (w/v) gelatin (incubation buffer). The grids were floated at RT sequentially on drops of incubation buffer containing (a) 0.02 M glycine for 10 min, (b) 5% (v/v) goat serum for 15 min, and (c) the primary antibody IEC 1/48 diluted 1:100 (v/v) for 30 min. After washing, grids were transferred for 20 min onto drops of goat anti-mouse IgG-coated 10-nm colloidal gold (Amersham International; Amersham, UK) diluted 1:40 (v/v) in incubation buffer. Controls were floated on incubation buffer without antibody IEC 1/48. After washing, grids were floated on 2% (w/v) aqueous OsO4 for 15 min, washed three times with distilled water, floated on 0.5% (w/v) uranylacetate, and thin-embedded with 2% (w/v) polyvinyl alcohol (Tokuyasu 1989).
Results
Protein Expression
We first examined the expression of Na+,K+-ATPase at the protein level in control and cholestatic rat liver by Western blot analysis of the total particulate fraction. The data demonstrated that in all control and experimental groups the antiserum recognized a protein of identical electrophoretic mobility (Mr 110,000) corresponding to the α1-subunit (data not shown). To assess whether the amount of protein expressed was altered by cholestasis, we analyzed the immunoblots of all animals densitometrically. The data demonstrated wide differences among animals and indicated a cholestasis-associated decrease by 11% (5223 vs 5856 in controls) after EE treatment and an increase by 15% (5433 vs 4709) in the BDL group.
Enzyme Activity in Homogenates
Enzyme activity decreased to 90% of the control level in the EE group (63.4 vs 70.3 mU/mg protein) and rose to 110% (52.9 vs 48.3) in BDL. These alterations are means of widely differing individual values. They are in line with our data on protein expression. However, the activity of aminopeptidase N, a canalicular plasma membrane enzyme that is not affected by BDL (Stieger et al. 1994), was used as a control and was not altered by any treatment modality (data not shown).
Cytochemistry
Na+,K+-ATPase activity in situ was demonstrated ultrastructurally with the cerium method (Figure 1, left column). Livers of control animals treated with propanediol alone (Figure 1A) or subjected to sham operation displayed enzyme activity at the basolateral plasma membrane of hepatocytes, whereas no reaction product was observed at the canalicular cell pole. No differences were found within the liver acinus. Neither distribution nor intensity of reaction product were altered by treatment with EE (Figure 1B) or BDL (Figure 1C). Rarely, the canalicular cell pole also displayed reactivity after BDL (Figure 1C, arrow).
Immunocytochemistry
The subcellular distribution of Na+,K+-ATPase was examined morphologically by indirect immunostaining at the light and electron microscopic levels. Immunofluorescent localization of Na+,K+-ATPase in semi-thin sections (Figure 1, right column) showed that in control liver (Figure 1D) immunoreactivity was localized at the basolateral domain of hepatocytes and cholangiocytes but was absent in the canalicular plasma membrane. Neither EE (Figure 1E) nor BDL (Figure 1F) induced any appreciable alterations in the distribution pattern or in the intensity of immunofluorescence. Only BDL livers occasionally displayed a few weakly immunopositive canaliculi (Figure 1F, arrow). The ultrastructural localization of Na+,K+-ATPase was demonstrated by immunogold labeling of ultrathin cryosections (Figure 2). In all animal groups the antigen was expressed exclusively in the basolateral plasma membrane (Figure 2, left), whereas the canalicular domain (Figure 2, right) was immunonegative. In addition, the density of gold label showed no appreciable alterations in the different groups. No intracellular membranes or vesicular structures showed immunoreactivity. Very few canaliculi in livers of the BDL group displayed any gold particles.
Discussion
Cholestasis is associated with increased levels of cholephilic compounds in serum. This defect may be caused by paracellular regurgitation of bile constituents through defective tight junctions (Rahner et al. 1996) and by altered expression, distribution, and/or activity of transport systems (Bossard et al. 1993; Gartung et al. 1996; Simon et al. 1996) indirectly linked to Na+,K+-ATPase. Therefore, the present study examined Na+,K+-ATPase in BDL- and EE-induced cholestasis using biochemical, cytochemical, and immunological methods.
Our initial expectation of establishing a correlation between the three techniques at the animal level was not realized because no significant changes in Na+,K+-ATPase could be observed. Among the experimental groups, however, the results of the different methods agreed well. Our data indicate that protein expression and activity are not affected significantly by EE or BDL treatment. This parallels the subcellular protein distribution, which is not altered by either model of cholestasis.
A substantial decrease in EE-induced Na+,K+-ATPase activity has been reported previously (Simon et al. 1996). In contrast, our biochemical data suggest only a slight decrease (-10%) in protein expression and enzyme activity, the latter having been reported in earlier studies (Bossard et al. 1993, and literature cited therein). They are therefore in line with our cytochemical results, which do not show a difference in density of reaction product (Figures 1A-1C). However, we cannot rule out that impairment of Na+,K+-ATPase activity plays a role in EE-induced cholestasis. Discrepancies with earlier biochemical data may be caused by technical differences because in this study the livers were perfused thoroughly before processing for analysis. Earlier reports showed that BDL does not alter Na+,K+-ATPase activity (Toda et al. 1978) or its mRNA level (Gartung et al. 1996). This is in line with our findings that show negliglible increases in protein expression and activity in homogenate.
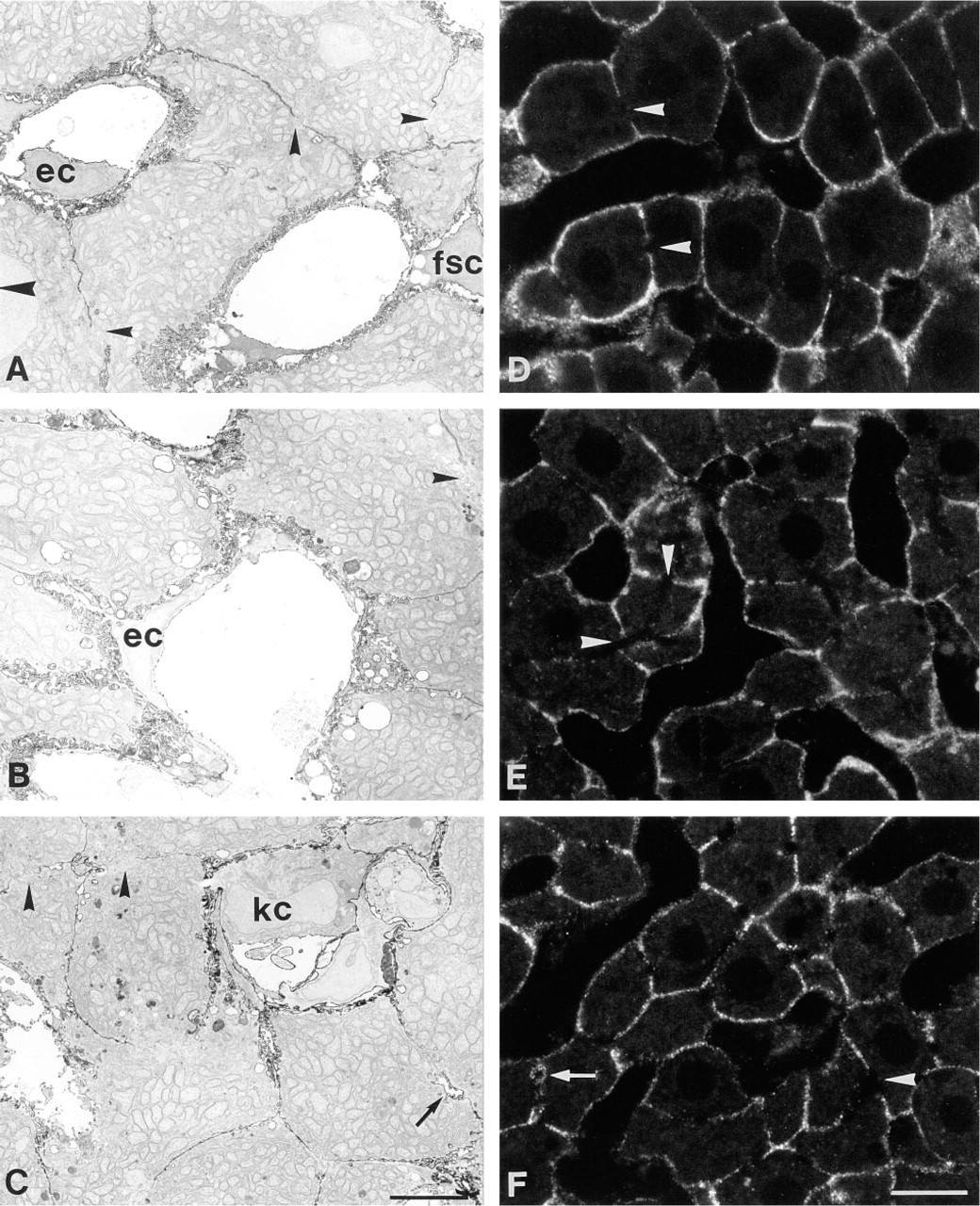
Cytochemical demonstration of Na+,K+-ATPase activity (left) and immunolocalization of protein expression (right) in normal and cholestatic rat liver. The micrographs show representative samples of control animals treated with propanediol (
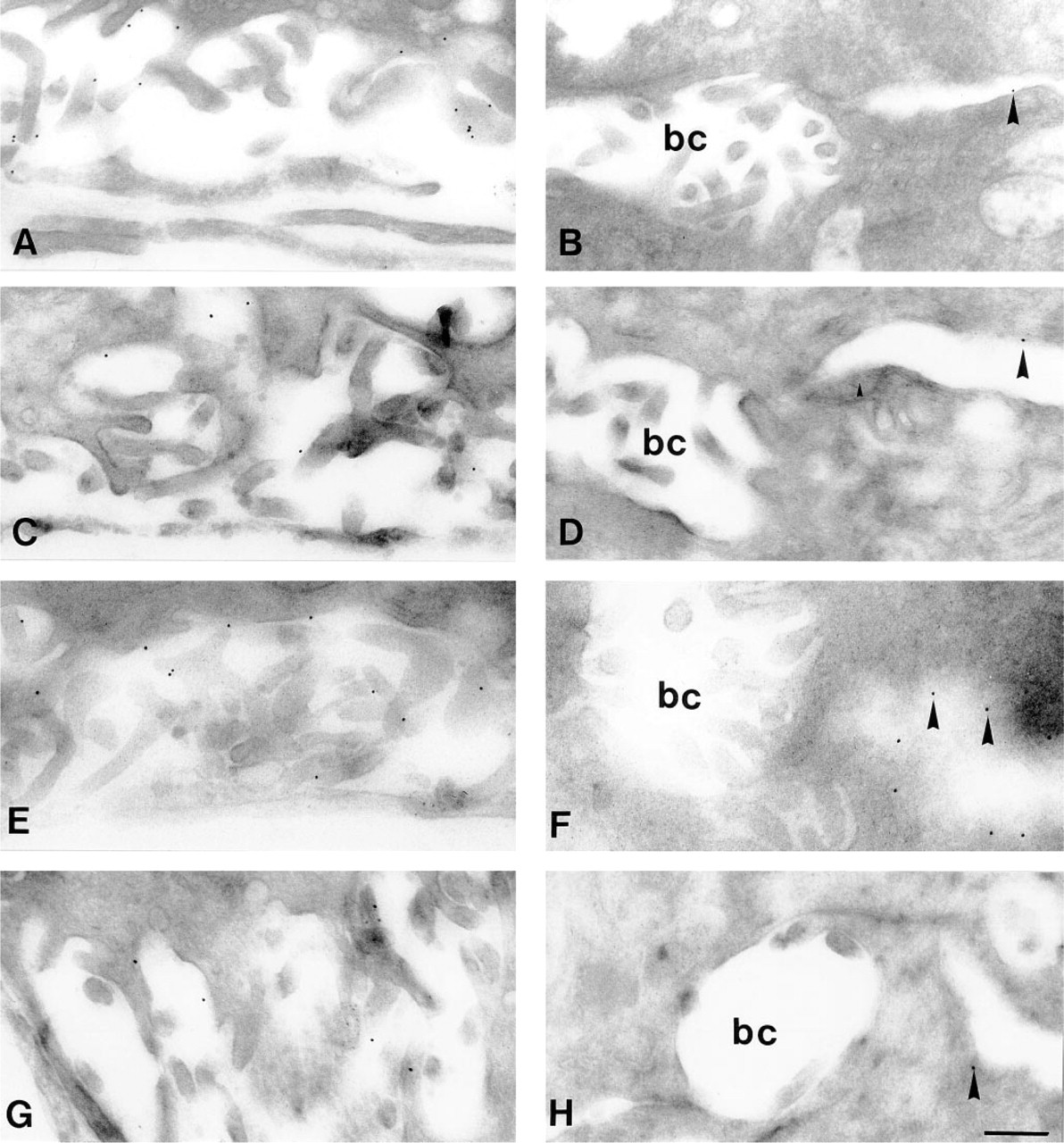
Immunoelectron micrographs of control and cholestatic rat liver. Ultrathin cryosections were incubated sequentially with monoclonal antibody IEC 1/48 and with secondary antibody-coated 10-nm colloidal gold. The sinusoidal membrane domain is displayed in the left column (
Although most studies using cytochemical (Latham and Kashgarian 1979; Angermüller et al. 1995) or biochemical in combination with cell fractionation (Toda et al. 1978; Sellinger et al. 1990) techniques indicate a basolateral localization of the functionally competent enzyme, immunocytochemical data are controversal. Some studies showed that the α-subunit is localized to the basolateral plasma membrane domain (Latham and Kashgarian 1979; Sztul et al. 1987), whereas others suggested both a basolateral and a canalicular localization (Schenk and Leffert 1983; Simon et al. 1995). The β-subunit was identified in the basolateral domain only (Simon et al. 1995). Our data (Figures 1D-1F, and 2) support the emerging consensus that functionally competent α/β-complexes are expressed in the basolateral membrane only.
BDL, but not EE, has been demonstrated to cause redistribution of domain-specific basolateral and canalicular plasma membrane proteins (Stieger et al. 1994). In contrast, our data show that the polarized expression of Na+,K+-ATPase is not altered by either model of cholestasis (Figures 1D-1F). It is conceivable that cytoskeletal elements keep this enzyme in the basolateral domain, as demonstrated in kidney cells (Nelson and Hammerton 1988). Our observation of a few immunopositive and enzymatically active canaliculi in BDL animals indicates that this interaction is partially disturbed by severe cholestasis.
In conclusion, our data demonstrating unaltered subcellular distribution of Na+,K+-ATPase and only minor changes in protein expression, as well as in enzyme activity, suggest that this enzyme is only slightly affected by cholestasis. Therefore, other mechanisms appear to play a more decisive role in the pathogenesis of cholestasis.
Footnotes
Acknowledgements
Supported by grants An 192/1–2 from Deutsche Forschungsgemeinschaft (SA), 34-40486.94 (LL), and 31-33520.92 (BS) from the Swiss National Science Foundation.
We thank J.P. Boeglin and R. Plank for skillful technical assistance, and Dr A. Quaroni for the kind gift of antibody IEC 1/48.