
Abstract
Because of the presence of a low-permeability cuticle covering the animal, fixation of C. elegans tissue for immunoelectron microscopy has proved very difficult. Here we applied a microwave fixation protocol to improve penetration of fixatives before postembedding immunogold labeling. Using this technique, we were able to successfully localize several components of yolk (YP170) trafficking in both wild-type and transgenic strains expressing a vitellogenin::green fluorescent protein fusion (YP170::GFP). Green fluorescent protein (GFP) and its variants are commonly used as markers to localize proteins in transgenic C. elegans using fluorescence microscopy. We have developed a robust method to localize GFP at the EM level. This procedure is applicable to the characterization of transgenic strains in which GFP is used to mark particular proteins or cell types and will undoubtedly be very useful for high-resolution analysis of marked structures. (J Histochem Cytochem 49:949–956, 2001)
T
We have adapted the microwave fixation method (Jones and Ap Gwynn 1991; Kok and Boon 1990) to C. elegans to demonstrate the subcellular distribution of yolk (YP170) and its receptor in C. elegans tissue of wild-type and transgenic animals expressing VIT::GFP. Yolk is synthesized in the intestine of the adult hermaphrodite nematode (Kimble and Sharrock 1983). Yolk is composed of lipids and lipid-binding proteins called vitellogenins. Once synthesized, yolk is secreted by the intestine into the pseudocoelomic space or body cavity. Yolk must then pass through the gonadal basal lamina, move through sheath pores, and finally be endocytosed by the oocyte, for which it provides essential nutrients that will support the development of the embryo (Hall et al. 1999; Grant and Hirsh 1999). We have previously demonstrated by conventional EM fixation that this yolk protein transits the pseudocoelom as free particles up to 1 μm in diameter, but enters the oocyte through sheath pores as membrane-bound granules (Hall et al. 1999). Yolk uptake by the oocyte is mediated by the RME-2 receptor (Receptor-Mediated Endocytosis) of the low-density lipoprotein receptor superfamily (LDLR) (Grant and Hirsh 1999). This receptor was identified using a screen for mutants defective in transfer of yolk from the body cavity into the oocytes (Grant and Hirsh 1999; Grant et al. 2001). rme-2 mutants are characterized by morphologically normal oocytes that appear devoid of yolk. Instead, most yolk accumulates in the pseudocoelomic space of the parent. rme-2 mutants produce very few viable progeny. Perhaps those embryos that survive take up some yolk by an RME-2-independent mechanism.
Yolk and yolk receptor antibodies have been previously employed using standard immunohistochemistry methods to demonstrate the transfer of yolk between the intestine and oocyte of C. elegans (Hall et al. 1999; Grant and Hirsh 1999). Here we used the same antibodies to illustrate the subcellular distribution of several components of yolk trafficking using postembedding immuno-EM techniques. In addition, we have developed a method to localize GFP at the EM level in VIT:: GFP transgenic animals. This article compares the merits of several monoclonal and polyclonal antibodies against GFP to localize YP170::GFP in thin sections of worms fixed with a microwave fixation protocol.
Materials and Methods
Animals and Antibodies
Wild-type nematodes were cultured according to standard methods (Brenner 1974); all strains were grown at 18C. Transgenic strains expressing green fluorescent protein (GFP) were generated as described previously (Grant and Hirsh 1999). Antibodies raised against GFP were purchased from Clontech (Palo Alto, CA), Research Diagnostics (Flanders, NJ), and Quantum Biotechnologies (Montreal, PQ, Canada). Anti-vitellogenin antibodies (anti-YP170) described by Sharrock et al. (1990) were provided by T. Blumenthal (University of Colorado). Antibodies against the external or internal domain of the yolk receptor protein: anti-RME-2 EXT and INT were generated as described in Grant and Hirsh (1999). Gold (10-nm)-labeled secondary antibodies were purchased from Amersham (Arlington Heights, IL).
Comparison of several GFP antibodies a
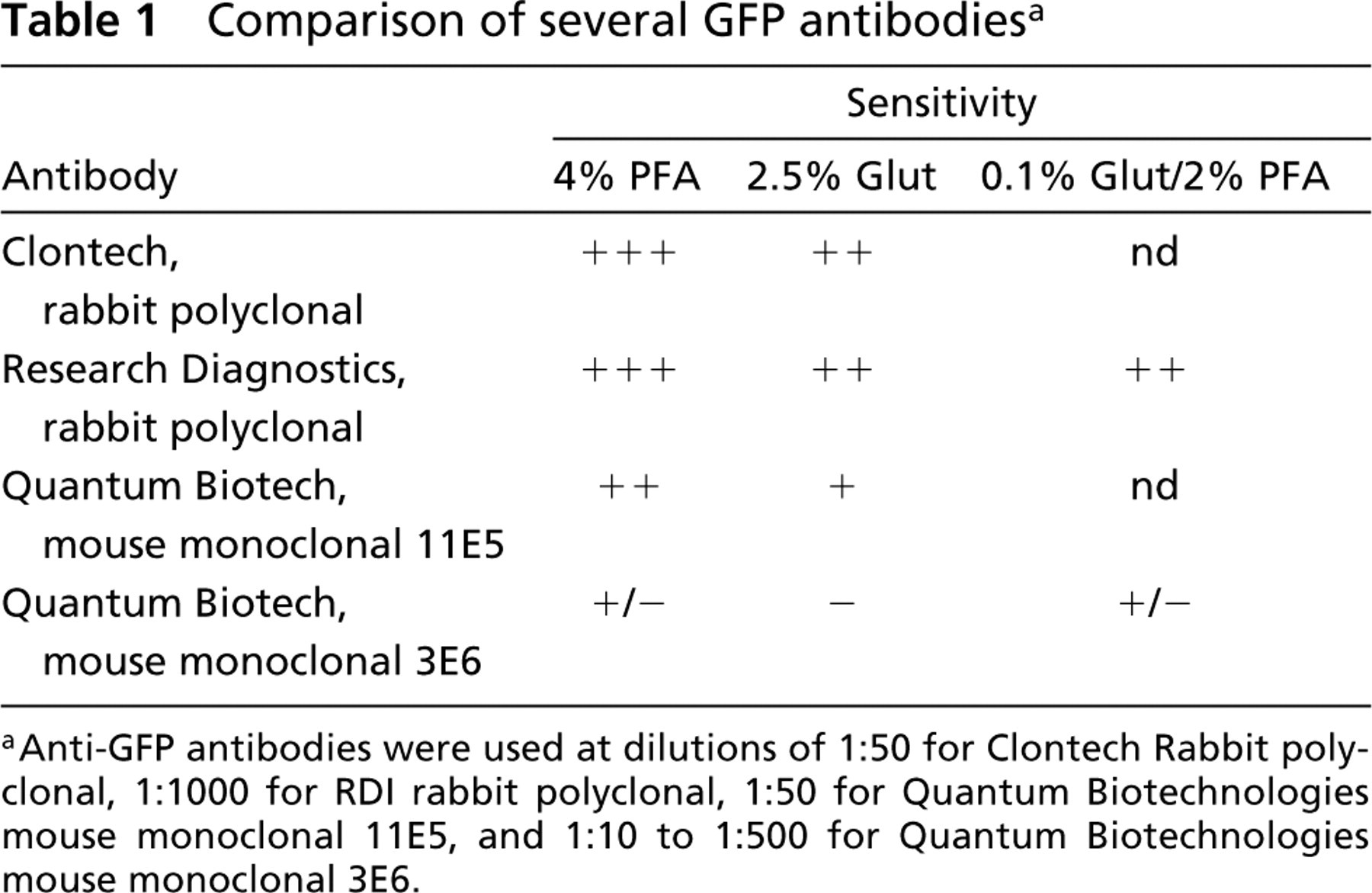
Anti-GFP antibodies were used at dilutions of 1:50 for Clontech Rabbit polyclonal, 1:1000 for RDI rabbit polyclonal, 1:50 for Quantum Biotechnologies mouse monoclonal 11E5, and 1:10 to 1:500 for Quantum Biotechnologies mouse monoclonal 3E6.
Microwave Fixation
Animals were rinsed in M9 buffer and placed into a pre-chilled glass well slide. M9 was replaced by a fixative solution consisting of glutaraldehyde and/or paraformaldehyde (Table 1) (Tousimis Research; Rockville, MD) in 0.12 M sucrose, 0.05 mM MgCl2, 0.1 M HEPES, pH 7.2. The specimens were then positioned at a “hotspot” within a Pelco Microwave Oven model 3450 (Ted Pella; Redding, CA) and irradiated at 50% power for 2 min. During this time, the glass well slide was kept chilled by a slurry of ice slush (∼150 g) and an additional water load of 400 ml was kept within the chamber. The sample volume was approximately 1 ml. Measurements showed that the sample was typically heated from 4C to 10C during this 2-min period. With microwave power off, the sample was allowed to cool for 2 min, during which time the ice slush was replenished. The sample was then irradiated for another 2 min, allowed to chill for 2 min, and irradiated a third time for 2 min. Therefore, total fixation time was only 10 min, with power on for 6 min total. Other microwave regimens were compared (e.g., 4 min on, 2 min off, 4 min on), but longer regimens caused excessive sample heating. After 10 min in fixative, samples were washed several times in 0.12 M sucrose in 0.1 M HEPES, pH 7.2, buffer, and embedded in agarose.
Dehydration and embedding protocol

Embedding in Agarose and LR Gold
Clusters of fixed animals were grouped together in 2.5% SeaPlaque agarose (Sigma; St Louis, MO) near its gelling temperature (30C) and allowed to chill and set (usually overnight) at 4C. Small agarose (2 mm3) blocks were then cut out to carry individual clusters through dehydration and embedding. Specimens were dehydrated through alcohols and into resin according to the schedule in Table 2. After embedment in LR Gold (Polyscience; Warrington, PA) plus 0.5% benzoin methyl ether accelerator (BME; Polyscience), samples were transferred to gelatin capsules in fresh resin, placed in a Pelco cryobox at approximately −20C, and exposed to UV radiation overnight. Most samples hardened within 15 hr; if necessary, the samples were sometimes exposed to an additional 12–24 hr of UV to complete curing.
Immunogold Labeling on Nickel Mesh Grids
Thin sections (60 nm) of LR Gold-embedded worms were collected on formvar-coated nickel mesh grids (Athene thin bar, 200 mesh; Ted Pella). If necessary, nearby sample sections were prescreened by EM to choose regions of special interest for immunostaining. Mesh grids containing sections were washed six times in 0.01 M glycine in 0.1 M phosphate buffer (PB), pH 7.4, at room temperature (RT), blocked with 0.5% gelatin, 0.01 M glycine, 0.5% nonfat dry milk, 1% normal goat serum in 0.1 M PB, pH 7.4, at 37C for 15 min, and washed once in warmed 0.01 M glycine in PB. Each grid was then inverted onto a small droplet (10–30 μl) of dilute primary antibody on Parafilm inside a humidified chamber. Antibody was diluted in 0.5% gelatin in PB and spun in a microfuge before use. After exposure to primary antibody for 1 hr at RT, grids were washed six times in PB at RT (floating on 1 ml of buffer while slowly agitating on a shaker table). Grids were inverted on small droplets of secondary gold-linked antibody (Amersham Auro-Probe; 1:30 dilution in 0.5% gelatin in PB). After exposure to secondary antibody for 1 hr at RT, grids were washed six times in PB, fixed in 2.5% glutaraldehyde in PB for 5 min, washed twice in dH2O, and stained in 2% uranyl acetate, pH 5.1, before examination by EM. Controls included (a) no primary antibody, (b) preimmune serum, if available, (c) positive control, i.e., another primary antibody, or (d) negative control, i.e., anti-GFP antibodies on wild-type sections.
Results and Discussion
Localization of GFP
For postembedding immuno-EM of VIT::GFP transgenic animals, several of the commercially available anti-GFP antibodies work very well, with generally low background (Table 1). In the case of VIT::GFP, some anti-GFP antibodies (Clontech and RDI rabbit polyclonal) remain active on tissues fixed under harsh fixation conditions (2.5% glutaraldehyde) (Table 1), thus allowing good preservation of ultrastructural details (Figure 1A). Similar fixation protocols should be satisfactory for any protein labeled with GFP, except in a few difficult tissues. We have begun to utilize these anti-GFP antibodies to follow the transport of secreted proteins from tissue to tissue (Hall et al. 1999). Here we show images from the vitellogenin::GFP (YP170::GFP) transgenic animals (Figure 1). To label low-abundance proteins, one might modify the technique to introduce an amplification of the signal, e.g., by using an enzymatic label such as HRP in place of the gold label.
When labeled by the postembedding technique, the YP170::GFP protein co-localizes in the same dark-staining organelles as native yolk protein in the intestine (Figures 1A and 1B). Gold label is visible in the pseudocoelom in discrete particles (not membrane-bound) (result not shown for anti-GFP; Figure 2 for anti-YP170) and is taken up into oocytes in small membrane-bound granules (Figures 1C and 2) (cf. Hall et al. 1999). Very similar results are obtained by labeling with anti-yolk antibodies (Figure 2). Unlike normal yolk particles, which are taken up specifically by the germline, the YP170::GFP particles in the pseudocoelom accumulate in oocytes and in scavenger cells called coelomocytes (Figure 3). Accumulation in coelomocytes is apparently stimulated by the GFP moiety, because yolk protein is not found there in wild-type worms (Bossinger and Schierenberg 1992; Hall et al. 1999).
RME-2 Receptor Localization
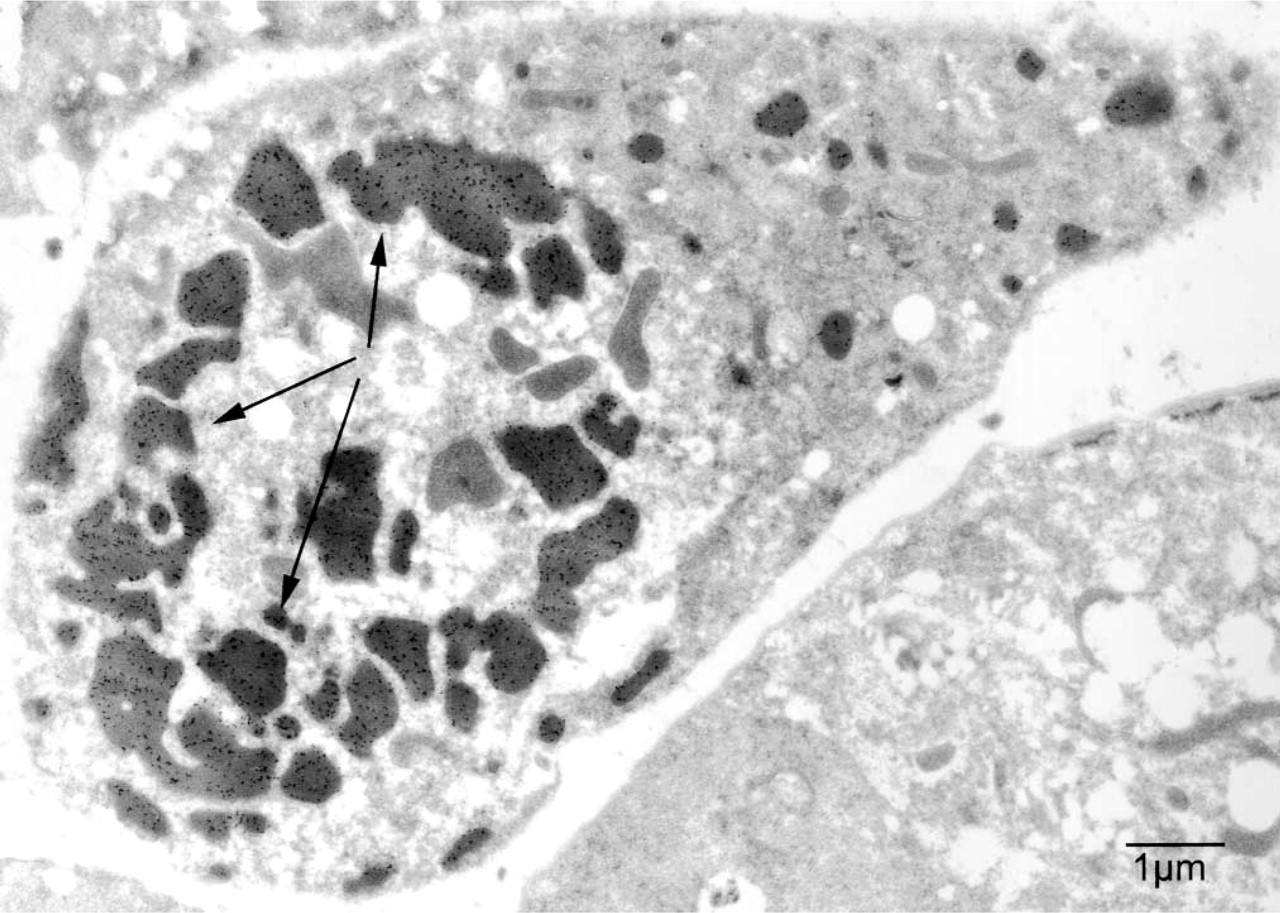
Another example of preserved antigenicity with microwave fixation is demonstrated by the localization of a new component of C. elegans yolk metabolism, the RME-2 receptor (Grant and Hirsh 1999). Using anti-RME-2 antibodies directed towards the extracellular domain of the protein (EXT), we find abundant levels of yolk receptors in the oocytes of adult hermaphrodites. Very little anti-RME-2 signal is detected at the plasma membrane. Instead, RME-2 is associated with internal membrane structures, probably recycling endosomes. Concomitant with yolk accumulation during oocyte maturation, yolk receptors are redistributed from deeper portions of the cytoplasm in early oocytes (not shown) to the cortical cytoplasm near the plasma membrane in the most mature oocytes (Figure 4A) (Grant and Hirsh 1999). After fertilization of the oocytes, the yolk receptor protein becomes internalized again, sinking away from the plasma membrane. Clusters of gold-decorated receptors are found inside the cytoplasm of the early embryo, with very little expression at the level of the plasma membrane (Figure 4B). The same results are obtained using an anti-RME-2 antibody directed towards the intracellular domain of the protein (INT) (data not shown). Our results using microwave fixation and postembedding immunogold labeling are in accord with earlier studies using immunofluorescence techniques (Grant and Hirsh 1999). Fertilized oocytes quickly secrete an eggshell and no longer have access to exogenous yolk particles. Embryonic tissues do not form yolk, but maternal yolk continues to be redistributed within the developing embryo, as monitored by alternative staining methods (Sharrock 1983; Bossinger and Schierenberg 1992). Association between RME-2 protein and yolk granules must be transient, occurring briefly during endocytosis, after which the receptor protein resides in aggregates either near the plasma membrane (at times of maximal uptake) or deeper in the cytoplasm. The fraction of receptor that remains away from yolk granules is strikingly high even at times near the peak of endocytosis, indicating efficient sorting of receptors and ligand soon after uptake. This feature is better scored by immuno-EM than by immunofluorescence.
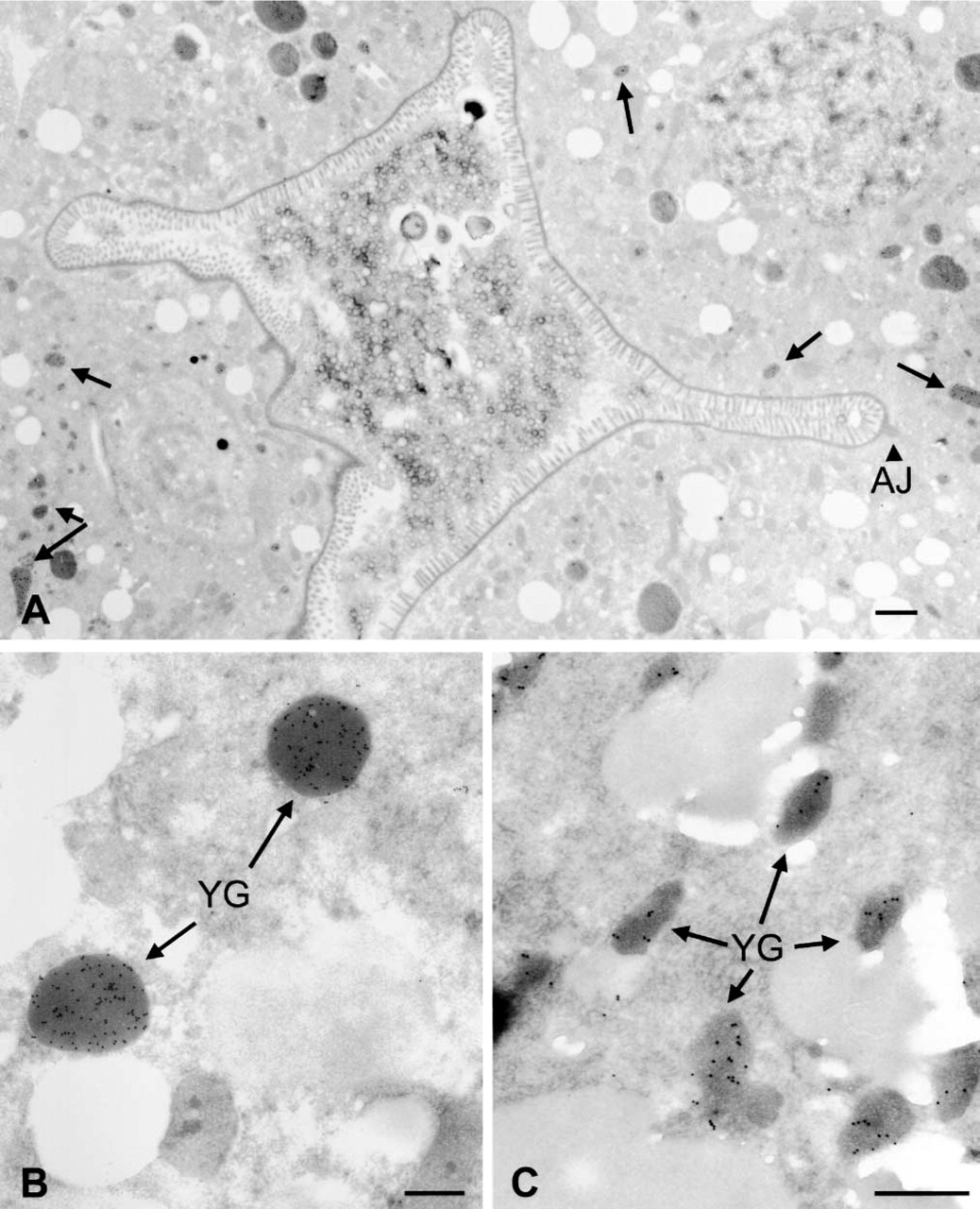
Anti-GFP antibodies label YP170::GFP in transgenic adults. (
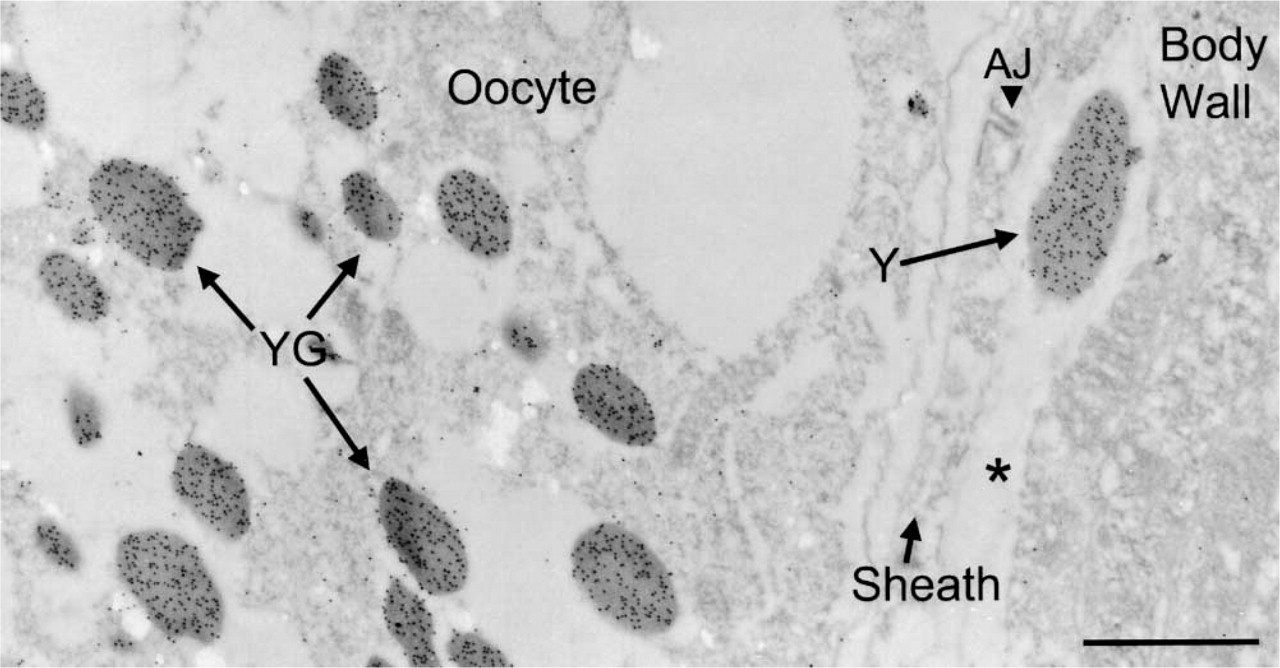
Anti-YP170 antibodies label yolk in wild-type adult. Gold label is seen over yolk granules (YG) in an oocyte and a free yolk particle (Y) in the pseudocoelom (asterisk) of a wild-type worm. An adherens junction (AJ) is visible in the gonad sheath. Microwave oven fixation using 4% paraformaldehyde. Bar = 1 μm.
Microwave Fixation Improves Antigen Preservation
Although tissue preservation after microwave fixation is not as good as after immersion fixation, antigenicity is generally excellent, perhaps because of the very short fixation time employed. At all steps we have endeavored to keep the sample temperature low to reduce damage to the epitope. The most obvious benefit of the microwave technique is that many animals in a sample are treated simultaneously and rapidly, whereas an immersion fixation involves the separate cutting by hand of each animal to provide access through the cuticle. The latter procedure is slow, tedious, and results in very irregular results because some animals are cut open shortly after going into fixation, some are cut open at long times after immersion, and some animals fail to be cut open at all. Because it is desirable to produce pellets containing hundreds of animals for the postembedding procedures, it becomes impossible to produce uniform treatment for all specimens by immersion and hand cutting.
The improvement in infiltration/embedment is most obvious in the preservation of embryos, which are impossible to cut open by hand. Their eggshell provides a second barrier to infiltration of fixative, solvent, or resin. In immersion fixations, the embryos typically produce crushed profiles or holes in the resin block because they do not infiltrate at all. The microwave fixation protocol allows fixatives and subsequent solutions to pass both the mother's cuticle and the embryonic eggshell so that those individual embryos are substantially better preserved (Figure 4B). With this technique, we have successfully labeled cell junctions in early and late embryos (unpublished data).
Our current microwave protocol is still not ideal. Preservation is only marginal for most tissues and is often inadequate for smaller cells and more delicate structures, including neurons and cell membranes. The nematode cuticle still is a barrier to the passage of fixatives and embedments that the microwave treatment alleviates but does not correct entirely. However, the resolution is good enough to recognize most tissues and organelles. Eventually, we expect that fast-frozen tissue may prove to be more suitable for postembedding immunocytochemistry to capture fragile membranes and processes (McDonald 1999). We are now comparing slam-freezing and high-pressure freezing protocols, followed by freeze substitution, as alternatives to microwave fixation. However, microwave fixation is a much cheaper and more accessible method for most laboratories, requiring fairly inexpensive low-tech equipment.
YP170::GFP is taken up by a coelomocyte. Anti-GFP (RDI) labeling demonstrates uptake of GFP-tagged yolk by a coelomocyte. Arrows point to yolk-filled endosomes. The clear space surrounding the coelomocyte is the pseudocoelom. Bar = 1 μm.
Protocols must still be tailored to suit the antigen and tissue(s) of interest. A central issue for immuno-EM is to preserve the epitope of interest while minimizing the loss of ultrastructural details. Every fixation procedure is a compromise and one must determine the best method for each tissue and antigen. Fixative choice is an important part of the equation. Glutaraldehyde, for example, reacts rapidly and irreversibly with proteins, making it the best fixative for ultrastructural preservation. However, for the same reason, its use often results in loss of antigenicity. Formaldehyde, a milder fixative that penetrates tissues more rapidly than glutaraldehyde, is gentler to the epitopes but does not provide very good preservation of ultrastructural details. A combination of formaldehyde and low concentrations of glutaraldehyde is often a good compromise. Individual epitopes are still likely to be affected differently by microwave fixation, dehydration, and embedding. Therefore, one needs to test alternative fixatives and treatments for each new antibody under study, as before. Similarly, some tissues may require different treatment regimens, or different osmolarities, to maximize their preservation. In our experience, it is generally helpful to conduct test fixations on at least three different fixative/buffer combinations when a new antibody for postembedding protocol is explored (Hall 1995). In all cases, one is trying to find the best balance between good structural preservation (harder fixation) and good epitope preservation (weaker fixation). Sensitivity of epitopes to dehydration remains a difficult obstacle for use of many available antibodies; perhaps 30–50% of all tested epitopes have failed to survive alcohol dehydration. Some epitopes may be more readily localized using frozen thin sections in place of plastic embedment (Selkirk et al. 1991). To the extent that we can settle on a few model epitopes for immunocytochemistry, such as the use of GFP as a marker, the same fixation protocol can be followed for many studies of transgenic strains and the same commercial antibodies can be utilized repeatedly.
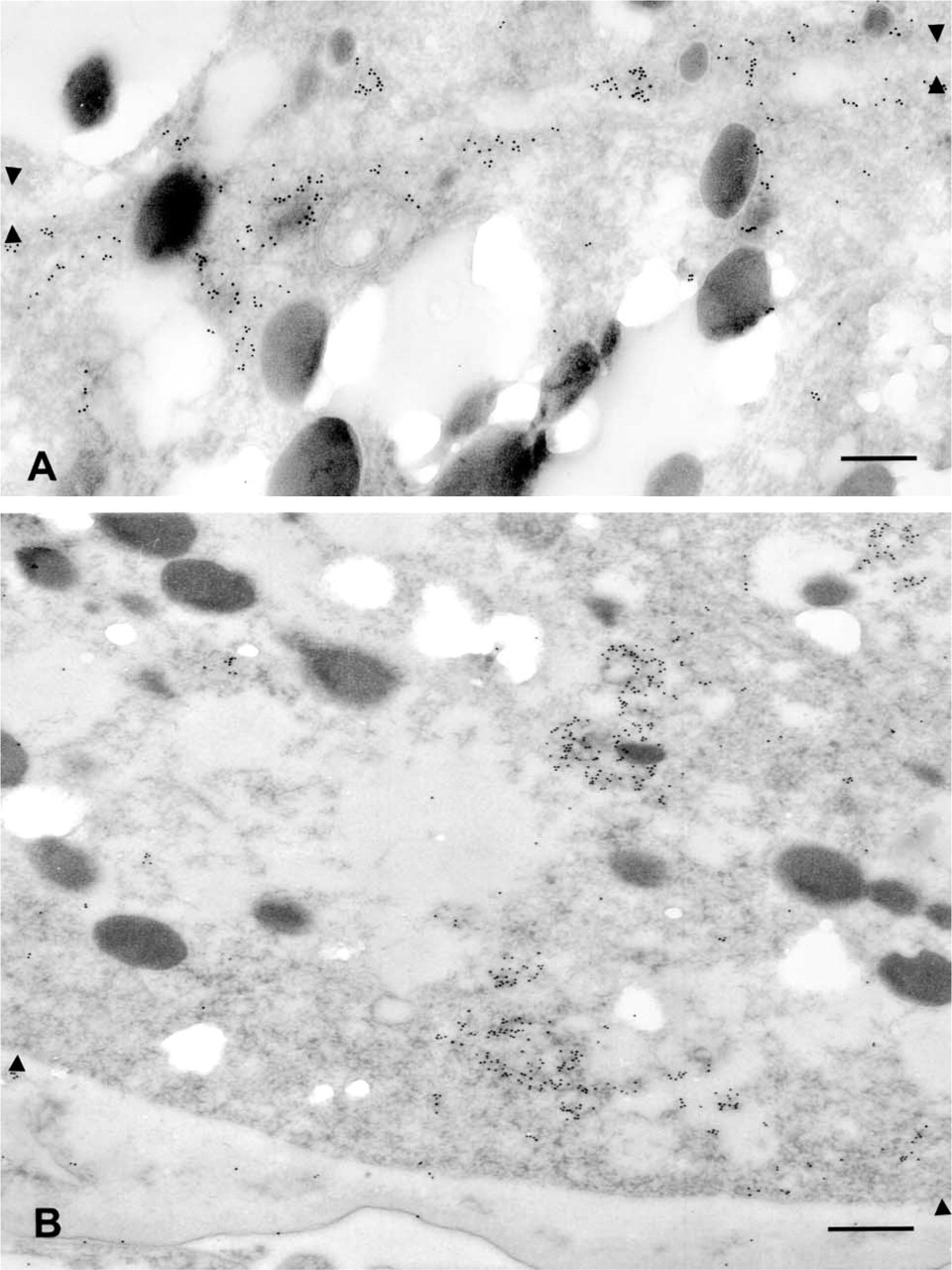
The yolk receptor protein (RME-2) localizes to different cellular compartments in full grown oocytes and in embryos of wild-type adult. (
Footnotes
Acknowledgments
Supported by grant RR12596 (NCRR) to the Center For C. elegans Anatomy (to DHH) and by March of Dimes grant FY99-583 (BG and DH).
We are grateful to Tylon Stephney (printing) and to Yongjing Li and Judith Kimble for excellent discussions about fixation protocols.