
Abstract
Recent studies suggest that carbon monoxide (CO), which is formed by the enzyme heme oxygenase (HO) during the conversion of heme to biliverdin, shares some of the chemical and biological properties of nitric oxide (NO) and may play roles similar to those of NO. Heme oxygenase activity in the kidney has been reported for many years, and there are some reports on the expression of mRNA for two HO isozymes (HO-1 and HO-2) and cellular localization of HO-1 protein. However, cellular localization of HO-2 protein in the kidney under normal conditions has not been reported. In the present study we examined the expression and distribution of HO-2 mRNA and HO-2 protein in rat kidney using RNA protection assay and light and electron immunocytochemistry. RNA protection assay confirmed constitutive expression of HO-2 transcript in rat kidney. HO-2 immunoreactivity was selectively found in epithelial cells of the thick ascending limb and distal convoluted tubule, connecting tubule cells, and principal cells of the collecting duct. These results suggest that HO-2 is synthesized in the kidney and that HO-2 in the epithelial cells of renal tubules may serve as a source for CO generation under normal conditions.
Keywords
N
Materials and Methods
Animals
Adult Wistar rats bred in our laboratory and kept under standardized animal room and food conditions were used.
Preparation of RNA Probe
Total cytoplasmic RNA was extracted from rat kidney and brain using the CLONsep total RNA isolation kit (Clontech; Palo Alto, CA). RNA concentration was determined by absorbance at 260 nm. Rat HO-2 cDNA was generated from rat kidney by using RT-PCR with the two oligonucleotide primers P1 (5′-TGAAAACTGGGAGGAGCAGG-3′), homologous to rat HO-2 cDNA nucleotides +543 to +562, and P2 (5′-AACAGGTAGAACTGGGTCCC-3′), complementary to rat HO-2 cDNA nucleotides +755 to +736 (Rotenberg and Maines 1990). The amplified 198-BP fragment was subcloned to TA Cloning Vector (Invitrogen; San Diego, CA) and sequenced using a T7 DNA polymerase-based kit (Amersham; Sunnyvale, CA).
RNA probes were generated from the restriction enzyme-linearized plasmids containing the 198
RNA Protection Assay
According to a previous method (Ausubel et al. 1995), both anti-sense and sense [32P]-RNA probes were hybridized to 10 μg of total RNA from rat kidney in 20 μl hybridization buffer overnight at 50C. RNase digestion was performed for 30 min at 37C, after which protected RNA hybrids were extracted with phenol/chloroform, precipitated with ethanol, redissolved in a formamide-denaturing buffer, and analyzed by electrophoresis on a 5% polyacrylamide-8 M urea gel.
Antibodies
Rabbit polyclonal antibodies raised against the synthesized N-terminal fragment of rat testis HO-2 were used. The preparation and specificity of the antibodies have been previously described in detail (Yamanaka et al. 1996).
Western Blot Analysis
Fresh kidneys and brains were homogenized in 50 mM Tris-HCl buffer, pH 8.0. Protein concentration was determined with a protein assay kit (Bio-Rad; Richmond, CA). Crude homogenates (50 μg) were loaded, separated on a 12% SDS-PAGE gel, and transferred onto a nitrocellulose membrane. The membrane was immunostained using the HO-2 antibodies and the peroxidase–anti-peroxidase complex (PAP) method.
Fixation
Rats were anesthetized with an
Light Microscopic Immunocytochemistry
One hundred-μm-thick sections of kidney were cut on a vibratome. Endogenous peroxidase activity and nonspecific staining were blocked by incubation with 0.3% hydrogen peroxide and 10% normal goat serum for 30 min. Then the sections were incubated with HO-2 antibodies at a concentration of 0.87 μg/ml in PBS overnight at room temperature. After washing with PBS, sections were immunostained by the PAP method and visualized with 3,3′diaminobenzidine tetrahydrochloride as chromogen. Sections were postfixed in 1.0% osmium tetroxide for 1 hr, dehydrated, and embedded in Epon resin. Semithin sections cut at 3-μm thickness were examined under a light microscope. Preimmune rabbit serum and preabsorbed antibodies, prepared by passing through the peptide-bound column, were used for control immunostainings.
Electron Microscopic Immunocytochemistry
One hundred-μm-thick vibratome sections were treated with Lugol's iodine solution for 10 min, rinsed with 2.5% sodium thiosulfate, then incubated with 10% normal goat serum for 10 min. Sections were incubated with HO-2 antibodies overnight at 4C, washed with PBS, and left in 3-nm gold-labeled goat anti-rabbit IgG (Funakoshi; Tokyo, Japan) for 3 hr. Then sections were developed with the silver enhancement kit (Amersham; Poole, UK) for 5–10 min and osmicated in 1.0% osmium tetroxide for 1 hr. Areas of interest were selected, dehydrated, and embedded in Epon resin. Seventy-nm-thick ultrathin sections were cut, counterstained in 4% uranyl acetate and lead acetate, and examined with an electron microscope H-800 (Hitachi; Tokyo, Japan). Control sections were made by omission of HO-2 antibodies.
Results
The constitutive expression of HO-2 mRNA was examined in rat kidney by RNA protection assay. As shown in Figure 1, a protected band of 198 nucleotides (lower arrowhead) was detected in the RNA samples of rat kidney and brain, a positive control, by using the anti-sense probe for the hybridization. However, no corresponding band was detected in yeast RNA or with sense probe. The relative abundance of HO-2 transcripts in the kidney was smaller than that in the brain.
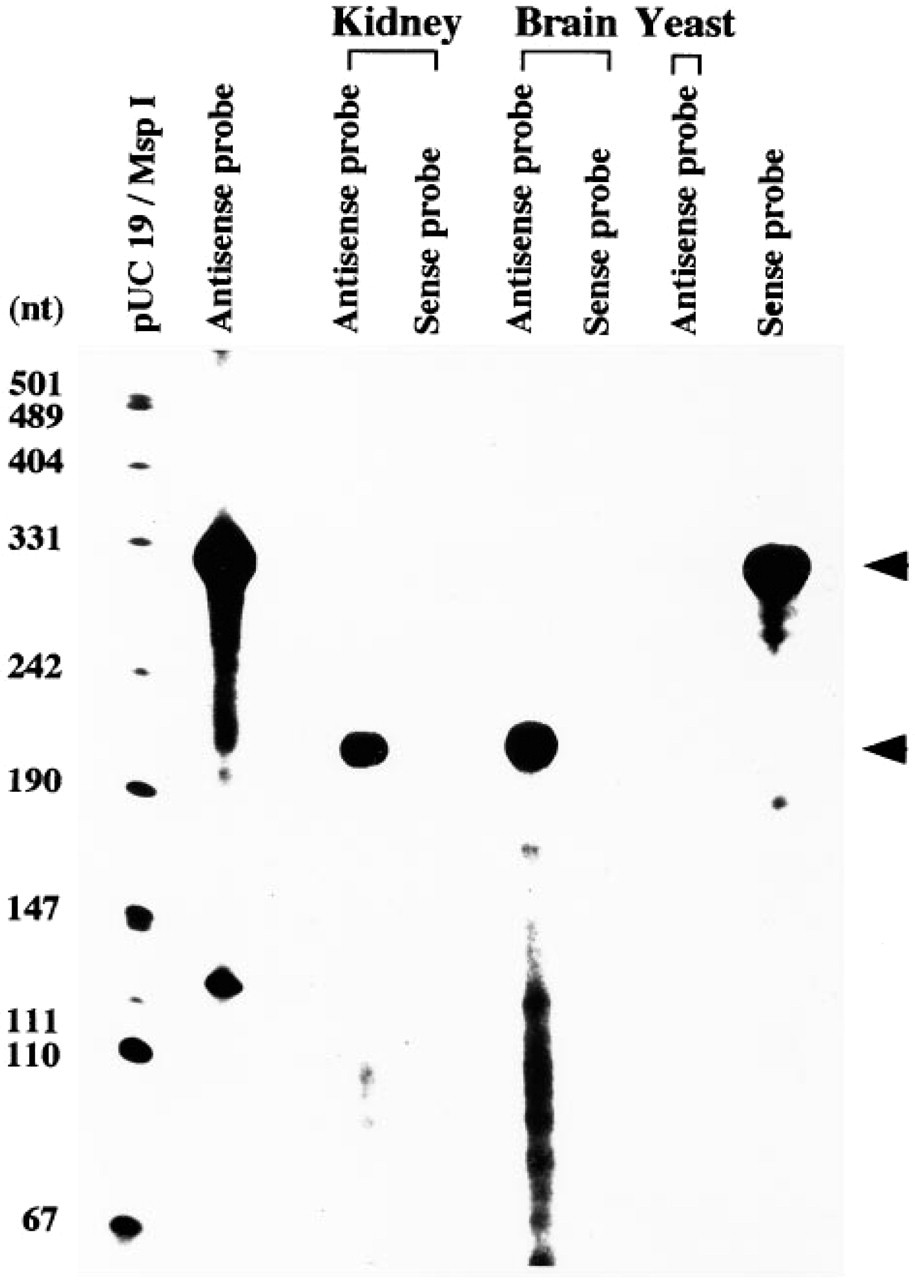
Detection of HO-2 mRNA in rat kidney by RNA protection assay. Upper arrowhead indicates the full length of sense and anti-sense RNA probes. Lower arrowhead shows the protected anti-sense fragments (198 nt) by HO-2 mRNA from the RNase digestion. The pUC19 vector, digested with restriction enzyme Msp I and labeled with [α-32P]-dCTP using the Klenow method, is used as the molecular weight marker.
The constitutive expression of HO-2 protein was examined in rat kidney by Western blot analysis. Immunoblottings from rat kidney and brain homogenates revealed a single clear protein band with a molecular mass of 36 kD in both lanes, with greater abundance in the brain (Figure 2).
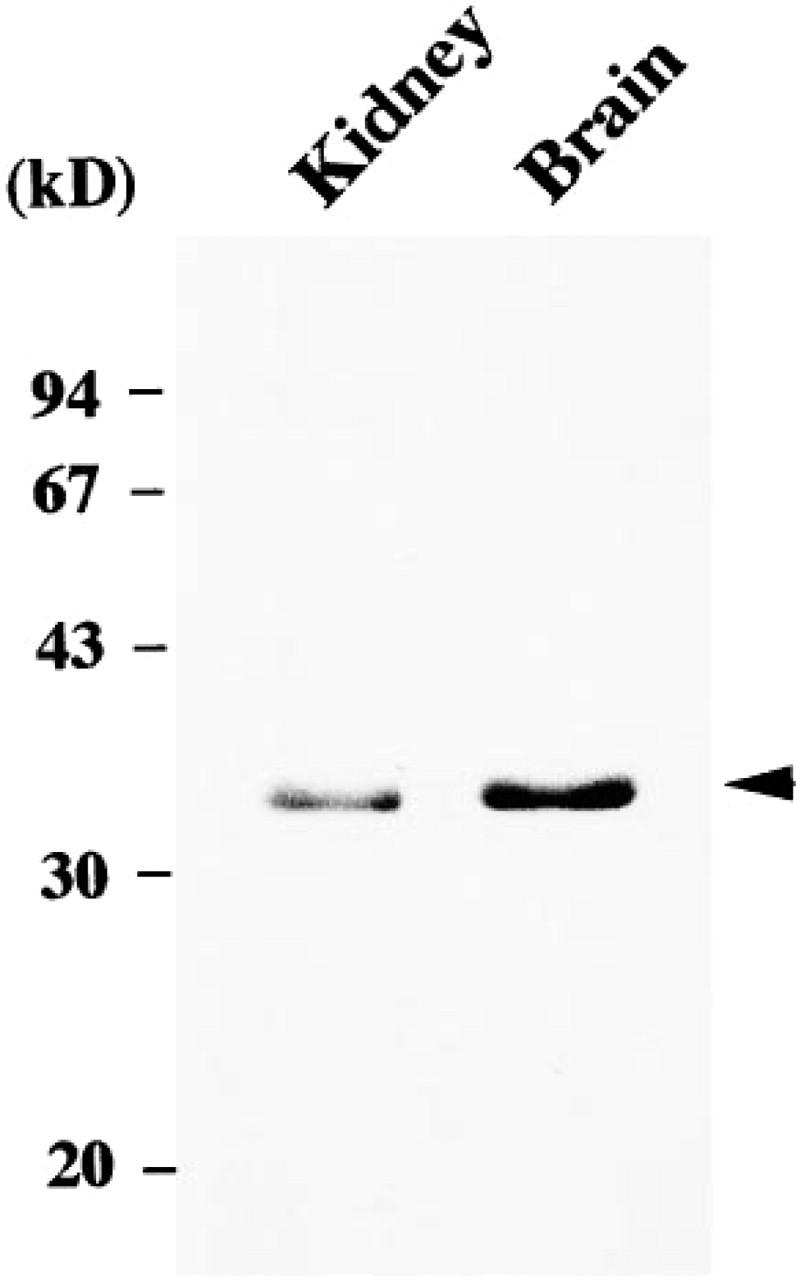
Detection of HO-2 protein in rat kidney by Western blot analysis. Crude homogenates (50 μg/lane) prepared from rat kidney and brain were used for electrophoresis, blotted onto nitrocellulose membranes, and stained with HO-2 antibodies. HO-2 immunoreactivity (arrowhead) was at 36 kD in both rat kidney and brain, with greater abundance in the latter. Molecular masses are indicated in kilodaltons.
Distribution of HO-2 protein in rat kidney under normal conditions was examined by light and electron microscopic immunocytochemistry using HO-2 antibodies. In light microscopic immunocytochemistry, positive staining for HO-2 was obtained in both the renal cortex and the medulla. In the renal cortex, on the basis of cell morphology and cell composition, immunoreactive segments were identified as the distal convoluted tubule, the connecting tubule, and the cortical collecting duct, as follows. The distal convoluted tubule is composed of homogeneous epithelial cells which have extensive basolateral membrane interdigitations (Stanton et al. 1981). Epithelial cells in Figures 3A and 3D, heavily stained in the cytoplasm except for the rim of the basal infoldings, were identified as epithelial cells of the distal convoluted tubules. The cortical collecting duct contains principal cells and intercalated cells. Principal cells are not regularly associated with the basolateral membrane infoldings (Stanton et al. 1981), and intercalated cells represent about 40% of the cells in the cortical collecting duct (Ritter et al. 1995). As shown in Figures 3A–3D, more than half of the epithelial cells in the collecting duct revealed diffuse cytoplasmic staining, whereas the remaining subpopulation of cells were not stained. Therefore, these immunoreactive cells could be identified as principal cells. The connecting tubule contains connecting tubule cells and intercalated cells. The connecting tubule cell is structurally intermediate between the distal cell and the principal cell, and the percentage of intercalated cells in the connecting tubule is also 40%, like that in the cortical collecting duct (Ritter et al. 1995). As shown in Figures 3B and 3C, more than half of the epithelial cells in the connecting tubule were clearly stained in the cytoplasm, except for the rim of basal infoldings, whereas the remaining cells exhibited no staining. The stained cells could be identified as connecting tubule cells. In the outer medulla, many of the epithelial cells of the collecting ducts and all of the epithelial cells of thick ascending limbs were intensely stained (Figure 4A). In the inner medulla, immunoreactivity was found only in the epithelial cells of the collecting ducts (Figure 4B). The immunoreactivity described above was absent in sections treated with preimmune serum or preabsorbed serum. However, some interstitial cells were in the control sections weakly stained (Figure 3E).
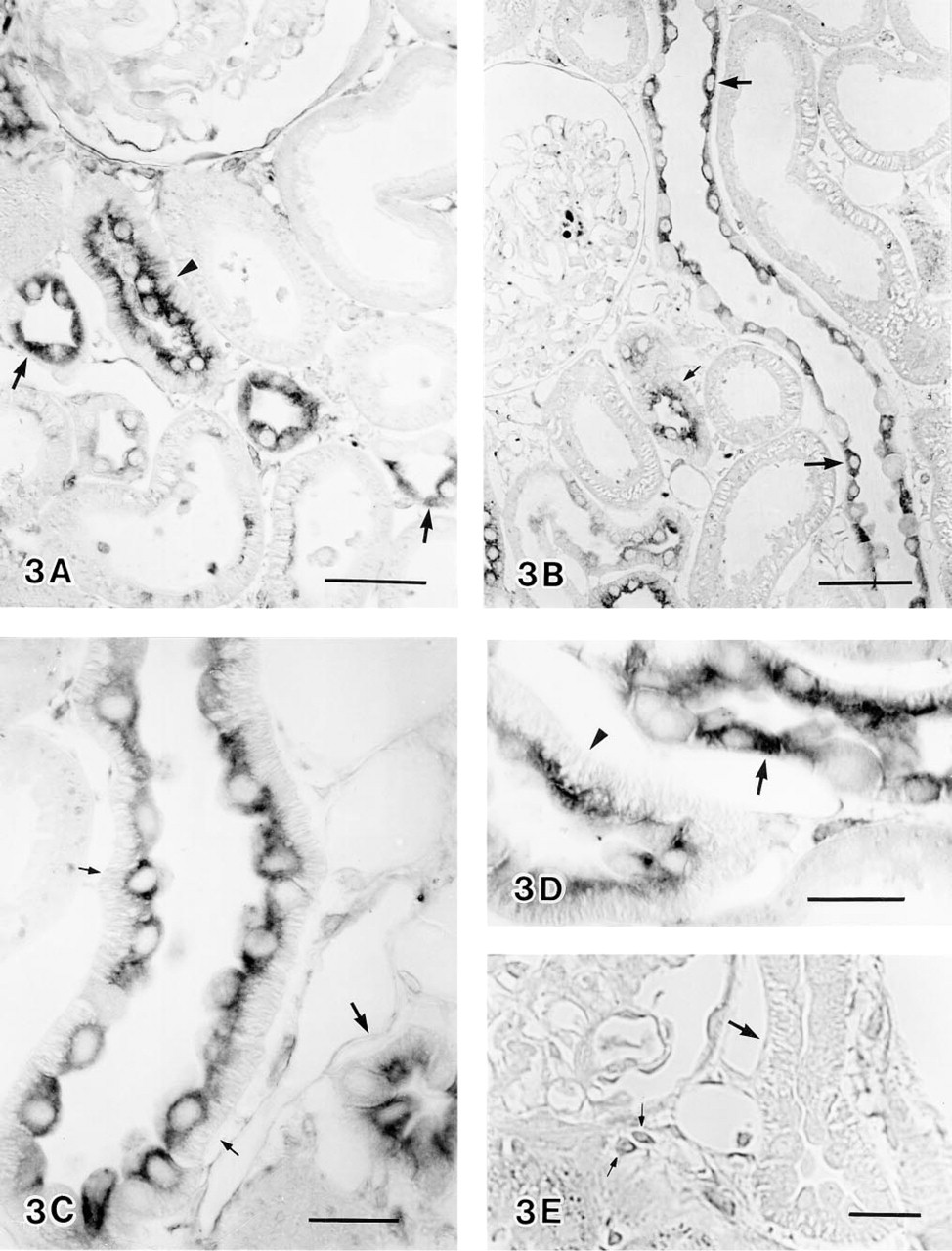
Light micrographs of HO-2 immunoreactivity in the renal cortex. (
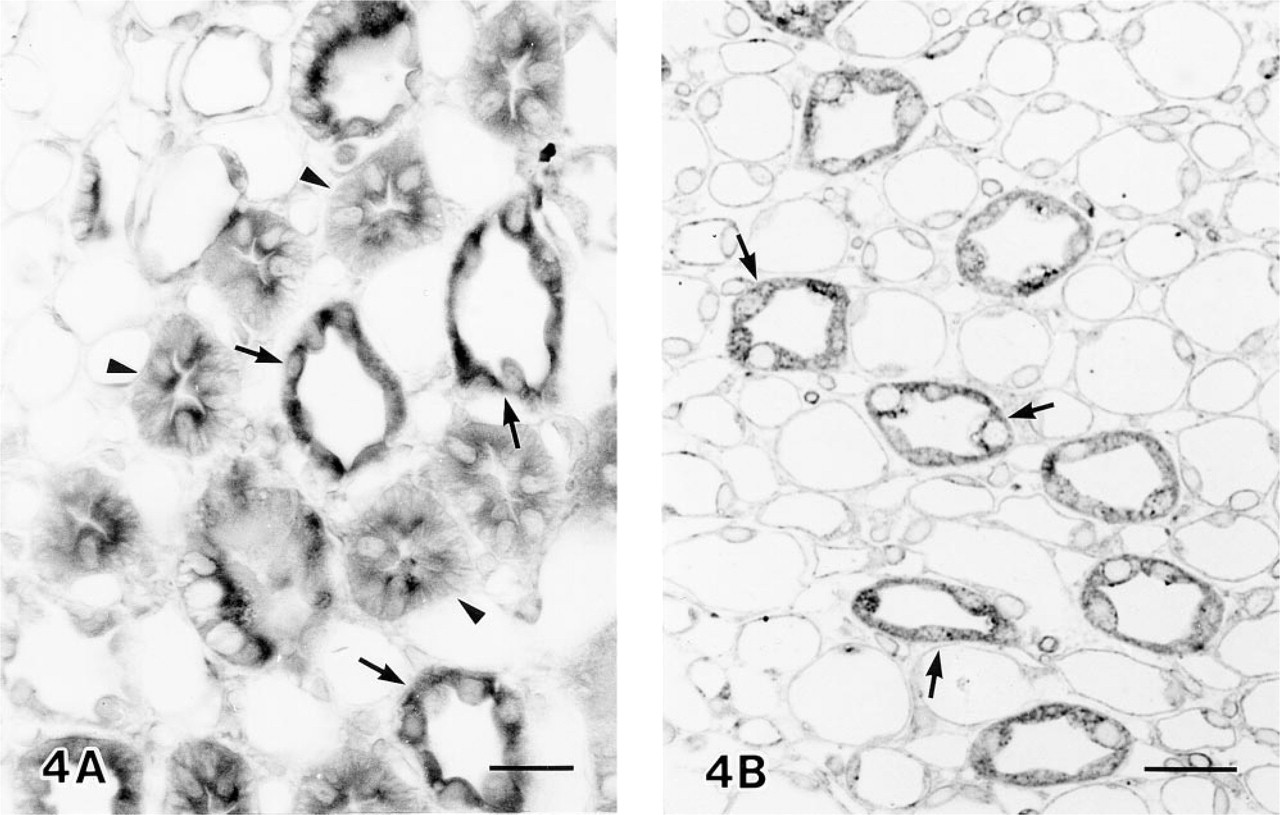
Light micrographs of HO-2 immunoreactivity in the outer (A) and inner (B) medulla. All the epithelial cells of thick ascending limbs (arrowheads) and most of the epithelial cells of the collecting ducts (arrows) were stained. Bars = 20 μm.
Electron microscopic immunocytochemistry further defined HO-2 immunoreactivity in the specific cells of the renal tubules. Figure 5 shows HO-2 immunogold–silver particles which were predominantly located in epithelial cells of the distal convoluted tubule (Figure 5A), the connecting tubule cells (Figure 5B), and the principal cells (Figure 5C). Particles were not found in the intercalated cells of the connecting tubule and the cortical collecting duct. Particles for HO-2 immunoreactivity were observed throughout the cytoplasm and were concentrated in the apical region of the stained cells. Control sections were made by omission of HO-2 antibodies. In these sections, positive staining was absent from the above-mentioned epithelial cells. However some particles were found predominantly on the surfaces of some interstitial cells (Figure 5D). At high magnification (Figures 6A and 6B), most of the silver-enhanced gold particles were associated with the endoplasmic reticulum and the nuclear outer membrane. In contrast, mitochondria, lysosomes, and Golgi complex were not labeled.
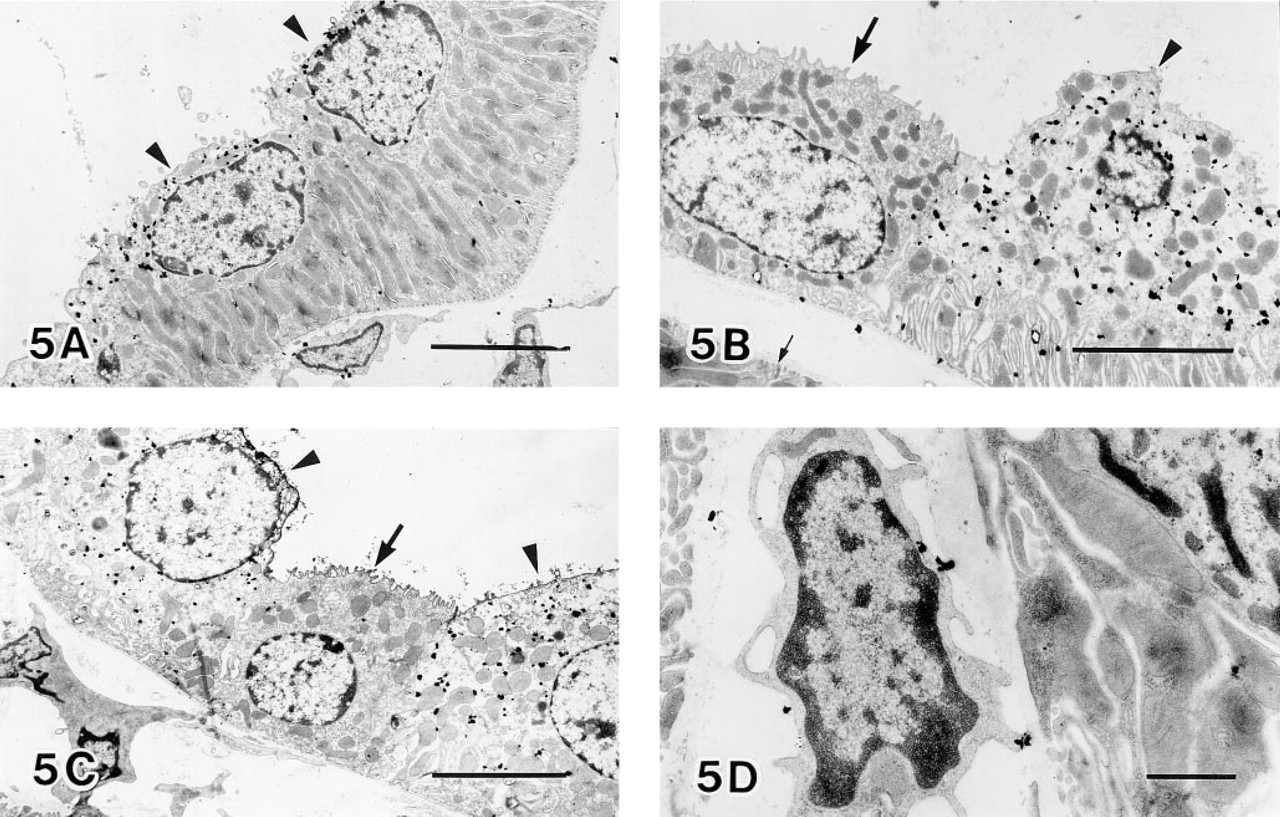
Electron microscopic observation of a distal convoluted tubule (
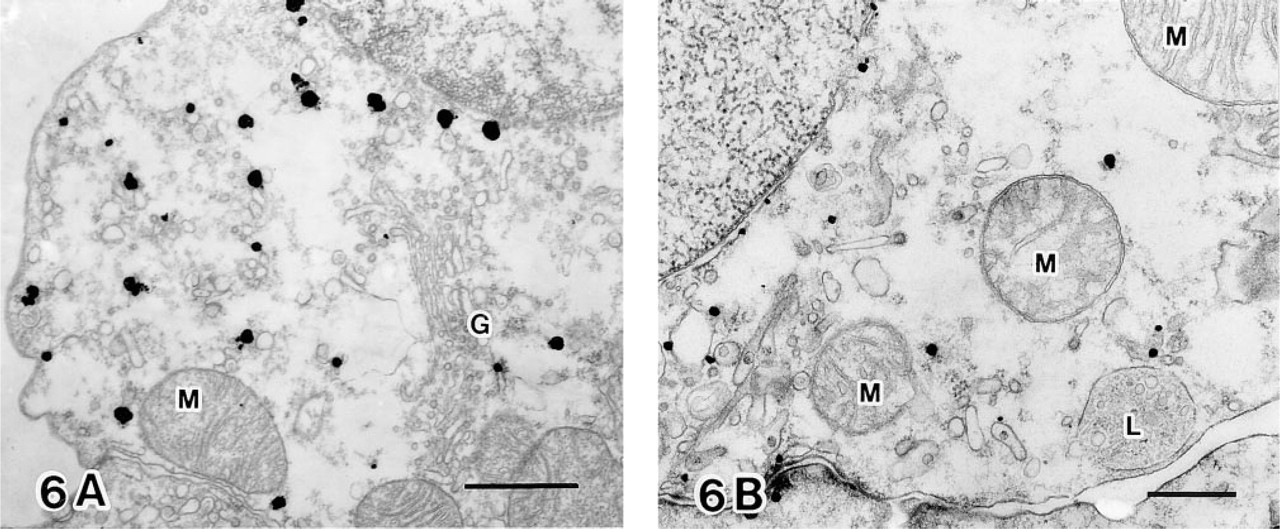
Electron micrographs at high magnification. (
Discussion
The RNA protection assay revealed a fairly high level of HO-2 transcript expression in rat kidney. Constitutive expression of HO-2 mRNA in rat kidney is consistent with the results of Northern blot analyses (Maines et al. 1993; Raju and Maines 1994). In addition to HO-2 mRNA expression, we also confirmed the presence of HO-2 protein in rat kidney by Western blot analysis and revealed the cellular distribution of HO-2 protein by immunocytochemistry.
Degradation of heme takes place largely in tissues of the reticuloendothelial system, such as the spleen, by heme oxygenase-1. The brain is an organ usually considered not to play a significant role in degradation of heme. However, HO-2 mRNA and protein are abundant in the brain (Ewing and Maines 1992; Verma et al. 1993; Yamanaka et al. 1996), and CO produced by the heme oxygenase has been postulated to function as a neurotransmitter, much in the same way as proposed for NO (Marks et al. 1991; Verma et al. 1993). The presence of a considerable amount of HO-2 mRNA and protein in the kidney and its distinct distribution in specific tubule epithelial cells suggest a potentially significant role of HO-2 in the normal kidney.
HO-2 immunoreactivity was found in the thick ascending limb, distal convoluted tubule, connecting tubule, and collecting duct. In the thick ascending limb and the distal convoluted tubule, all epithelial cells were immunoreactive, whereas in the connecting tubule and the collecting duct, immunonegative epithelial cells were found scattered among immunopositive epithelial cells. Those immunonegative cells were considered to be intercalated cells, based on the knowledge that the incidence of these cells is about 40% in the connecting tubule and the cortical collect duct and about 10% in the inner medullary collecting duct (Ritter et al. 1995). Electron microscopic observations of the immunoreactive and the immunonegative cells supported this view. At high magnification, HO-2 im-munogold–silver particles were determined to be associated with the endoplasmic reticulum and nuclear outer membrane. Recently, it was reported that HO-1 is localized to interstitial cells of the rat kidney (Agarwal et al. 1996). In Figures 3D and 3E, some interstitial cells were weakly stained in both experimental and control sections, suggesting that the staining of some interstitial cells is nonspecific. In electron microscopy of control sections (Figure 5D), particles were found predominantly on the surfaces of some interstitial cells, suggesting that the cell surface of those cells may have a tendency to be labeled with anti-rabbit IgG. Therefore, the localizations of HO-1 and HO-2 in rat kidney are different. The function of HO-2 in renal tubule cells remains to be further studied. However, it is proposed that CO, like NO, stimulates SGC and exerts its effects by elevating cGMP level (Maines 1993). In the brain, the distribution of HO-2 protein closely parallels that of SGC, and it is suggested that CO may be the major physiological regulator of SGC (Verma et al. 1993). Recent studies indicated the presence of SGC mRNA and subunits in renal vasculature and in principal cells of the collecting duct (Ujiie et al. 1993; Mundel et al. 1995). Therefore, the present finding that HO-2 was localized in epithelial cells of the thick ascending limb and distal convoluted tubule, connecting tubule cells, and principal cells of the collecting duct suggests that CO produced in those cells can bind to SGC in the same and neighboring cells, elevate the cGMP level, and regulate the function of the cells containing SGC.
To our knowledge, the present study has revealed the precise intracellular localization of HO-2 protein in rat kidney for the first time. Although further work is necessary to clarify the physiological role of HO-2, our present results suggest that HO-2 is synthesized in the kidney and that HO-2 in the epithelial cells of the renal tubules may serve as a source for CO generation under normal conditions.