
Abstract
α-Amidation is catalyzed by two enzymatic activities, peptidyl-glycine α-hydroxylating mono-oxygenase (PHM) and peptidyl-α-hydroxyglycine α-amidating lyase (PAL), denoted collectively as peptidyl-glycine α-amidating mono-oxygenase (PAM), which also may include transmembrane and cytoplasmic domains. PAM is present in mammalian pancreas, where it appears to be abundant in the perinatal period. Nevertheless, there is no agreement on the cell type(s) that produces PAM or even on its presence in adults. In the present study we found PAM (PHM and cytoplasmic domain) immunoreactivity (IR) in A-, B-, and D-cells of adult mouse pancreas. In contrast to previous reports, PAM IR was found in B-cells of human and rat. Most of the B/D-cells were PAM immunoreactive, although with variable intensity, whereas less than half of A-cells displayed IR. Immunocytochemistry and Western blotting suggested the existence of different PAM molecules. Differences in the cellular distribution of IR for PAM domains were also observed. Whereas PHM-IR was extended throughout the cytoplasm in the three cell types, presumably in the secretory granules, IR for the cytoplasmic domain in A/D-cells was restricted to a juxtanuclear region, perhaps indicating its cleavage in Golgi areas. Although glucagon, insulin, and somatostatin are non-amidated, amidated peptides (glucagon-like peptide 1, adrenomedullin, proadrenomedullin N-terminal 20 peptide) were found in the three cell types.
R
The formation of an amidated peptide from its glycine-extended propeptide requires an enzyme complex denoted collectively as peptidyl-glycine α-amidating mono-oxygenase (PAM) (Eipper et al. 1983). α-Amidation is a two-step process catalyzed by two separate enzyme activities, peptidyl-glycine α-hydroxylating mono-oxygenase (PHM) and peptidyl-α-hydroxyglycine α-amidating lyase (PAL), both encoded by the same gene, which has been cloned and appears to be highly conserved among different species (Eipper et al. 1987). The PHM enzyme is contained in the amino-terminal third of the PAM precursor (Figure 1), followed by the PAL enzyme (Glauder et al. 1990; Stoffers et al. 1991; Milgram et al. 1992). The carboxy-terminal third of the molecule encodes a transmembrane domain and a hydrophilic cytoplasmic domain (Eipper et al. 1987). The diversity found in PAM enzymes is due to tissue-specific PAM mRNA alternative splicing and/or post-translational proteolysis, which yields different bifunctional and monofunctional enzymes as well as soluble and membrane-bound forms (Eipper et al. 1992a; Milgram et al. 1992).

PAM precursor and antisera used in the study.
The amidating enzymes have been identified in diverse tissues of mammalian species, such as heart, neurons, and glial cells, and in several exocrine and endocrine glands (Sakata et al. 1986; Eipper et al. 1988; Ouafik et al. 1989; May et al. 1990; Rhodes et al. 1990; Braas et al. 1992; Schafer et al. 1992; Martinez et al. 1993a).
In relation to the presence of PAM in endocrine tissue, only a few studies refer to the pancreas. Most of them show evidence of PAM existence in pancreatic tissue extracts, and only two studies, carried out in human (Martinez et al. 1993b) and rat (Braas et al. 1992), used immunocytochemical methods to detect the amidating enzymes in situ. PAM activity has been described biochemically in the pancreas of several mammalian species, including rat (Sakata et al. 1986; Ouafik et al. 1987; Scharfmann et al. 1988; Maltese et al. 1989; Zhou and Thorn 1990), sheep (Kapuscinski and Shulkes 1995), and human (Tateishi et al. 1994). In such biochemical studies, a maximum of PAM enzymatic activity is generally reported in neonatal specimens. Moreover, some of the studies point out that in adult pancreas the enzyme PAM is almost undetectable in rat (Maltese et al. 1989). Nevertheless, the two immunocytochemical studies that detected PAM immunoreactivity (IR) in human and rat pancreas were carried out in adult specimens (Braas et al. 1992; Martinez et al. 1993b).
PAM antisera used in the study rendering immunolabeling a
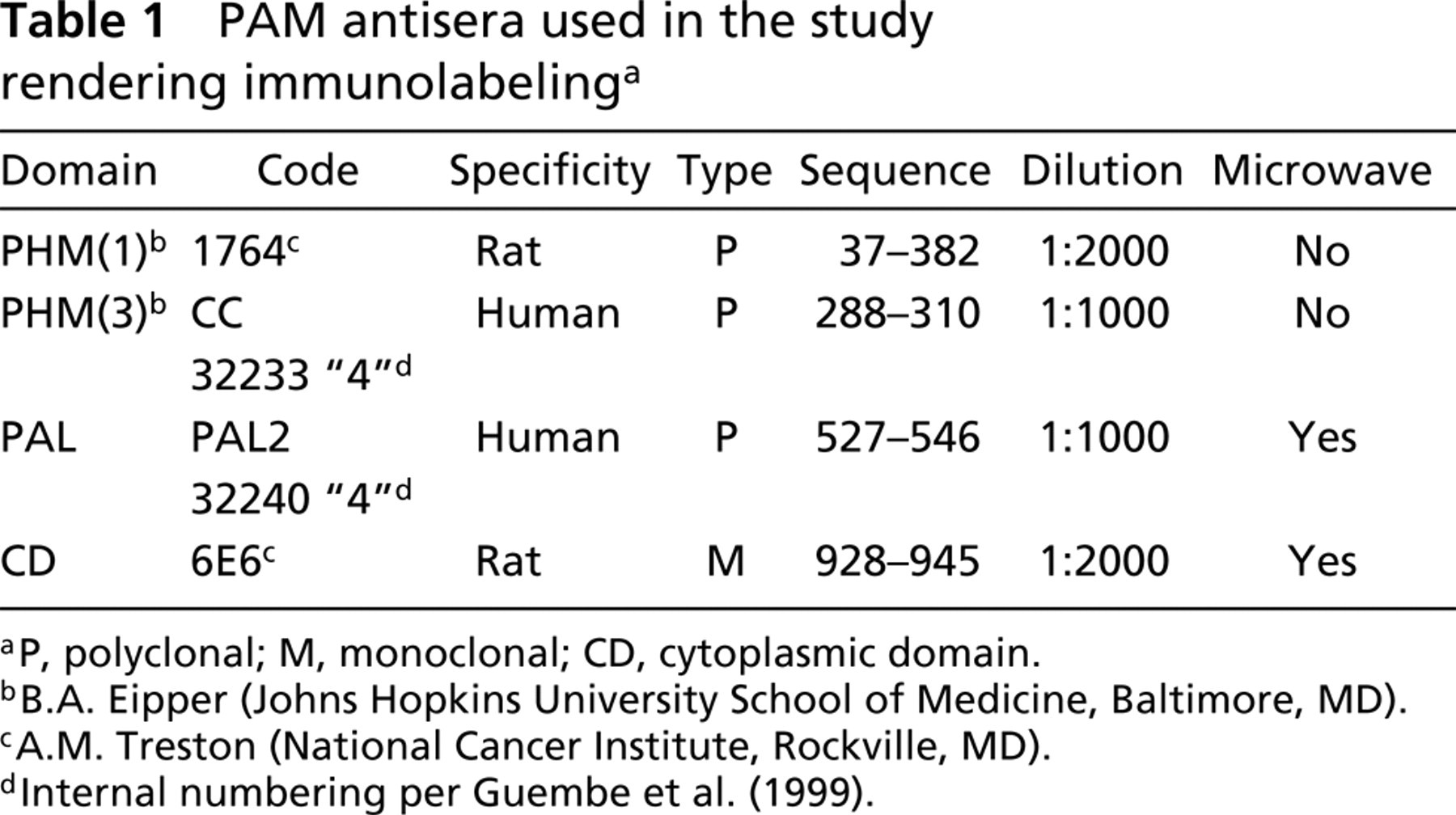
P, polyclonal; M, monoclonal; CD, cytoplasmic domain.
B.A. Eipper (Johns Hopkins University School of Medicine, Baltimore, MD).
A.M. Treston (National Cancer Institute, Rockville, MD).
Internal numbering per Guembe et al. (1999).
On the other hand, there is no agreement concerning the cell types that express the amidating enzyme. Some of the biochemical studies locate PAM activity in endocrine islets and suggest its presence in β-cells (Ouafik et al. 1987; Scharfmann et al. 1988; Maltese et al. 1989). Immunocytochemical studies have described the presence of PAM in pancreatic islets, namely in human A-cells (Martinez et al. 1993b) or in rat “peripheral” (interpreted as A/PP) cells (Braas et al. 1992).
The aim of the present study was the immunocytochemical study of the enzyme PAM in a third mammalian species (mouse), to address the two above-mentioned controversial aspects: the presence of the enzyme PAM in adult mammalian pancreas and the cell type(s) that produces the enzyme.
Antisera against pancreatic and amidated peptides a
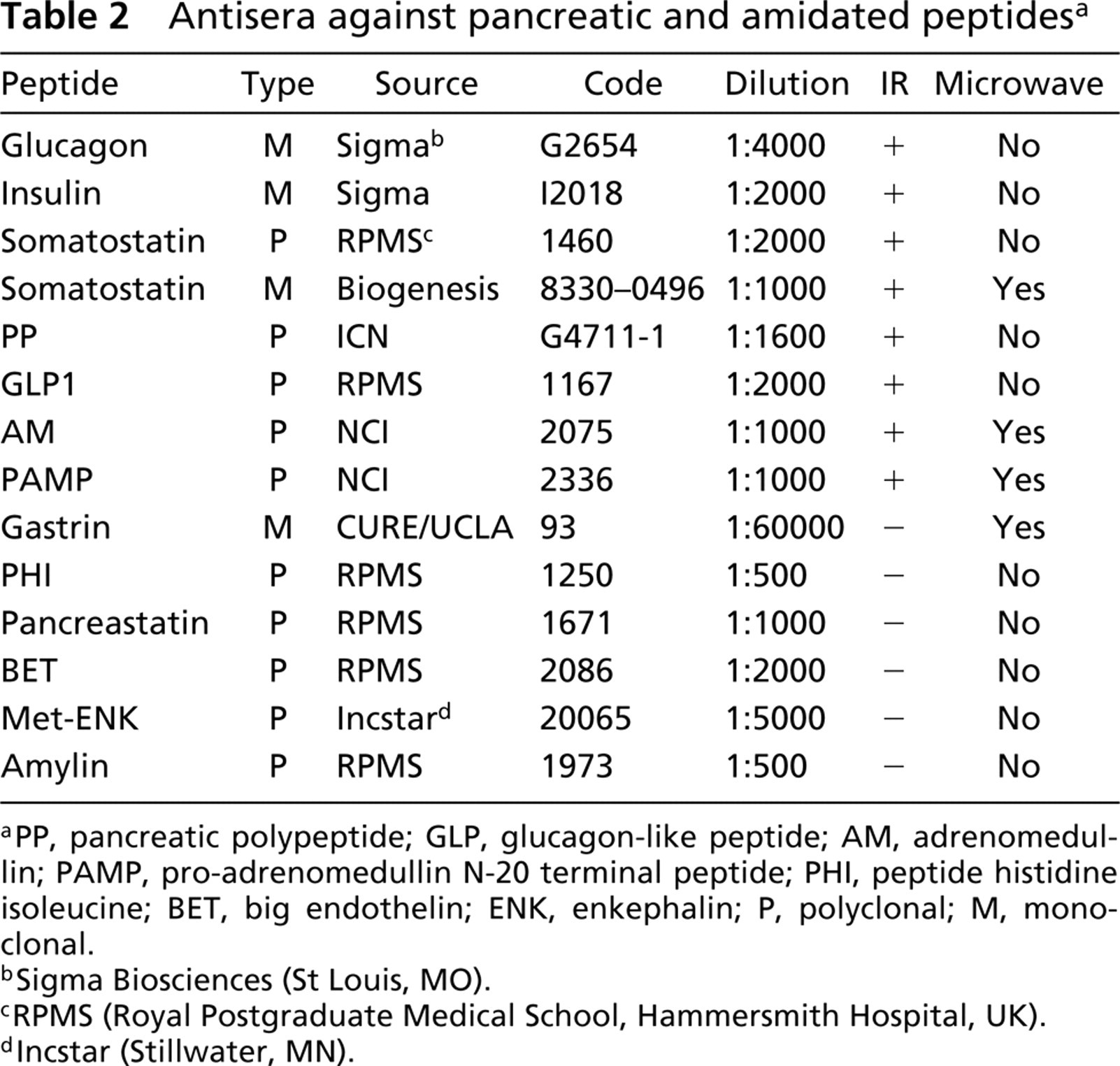
PP, pancreatic polypeptide; GLP, glucagon-like peptide; AM, adrenomedullin; PAMP, pro-adrenomedullin N-20 terminal peptide; PHI, peptide histidine isoleucine; BET, big endothelin; ENK, enkephalin; P, polyclonal; M, monoclonal.
Sigma Biosciences (St Louis, MO).
RPMS (Royal Postgraduate Medical School, Hammersmith Hospital, UK).
Incstar (Stillwater, MN).
Materials and Methods
For immunocytochemical methods, 24 female Swiss mice were sacrificed by cervical dislocation and their pancreata fixed, embedded in paraffin or resin, and cut into sections. Because immunocytochemical staining may vary according to tissue processing, different processing protocols were carried out. Pancreata embedded in paraffin were previously fixed in either Bouin for 24 hr (10 pancreata), Zamboni for 24 hr (two pancreata), or PAF 4% for 4 hr (two pancreata). Before resin embedding, either Zamboni fixative for 24 hr (five pancreata) or PAF 4%–glutaraldehyde 1% fixative for 2 hr (five pancreata) was used. Paraffin- and resin-embedded material was cut into 4-μm and 1-μm sections, respectively. Bouin- and formol-fixed rat and human pancreata were used as controls.
For Western blotting analysis, five female Swiss mice were sacrified by cervical dislocation and their pancreata frozen in liquid nitrogen and stored at −80C.
(
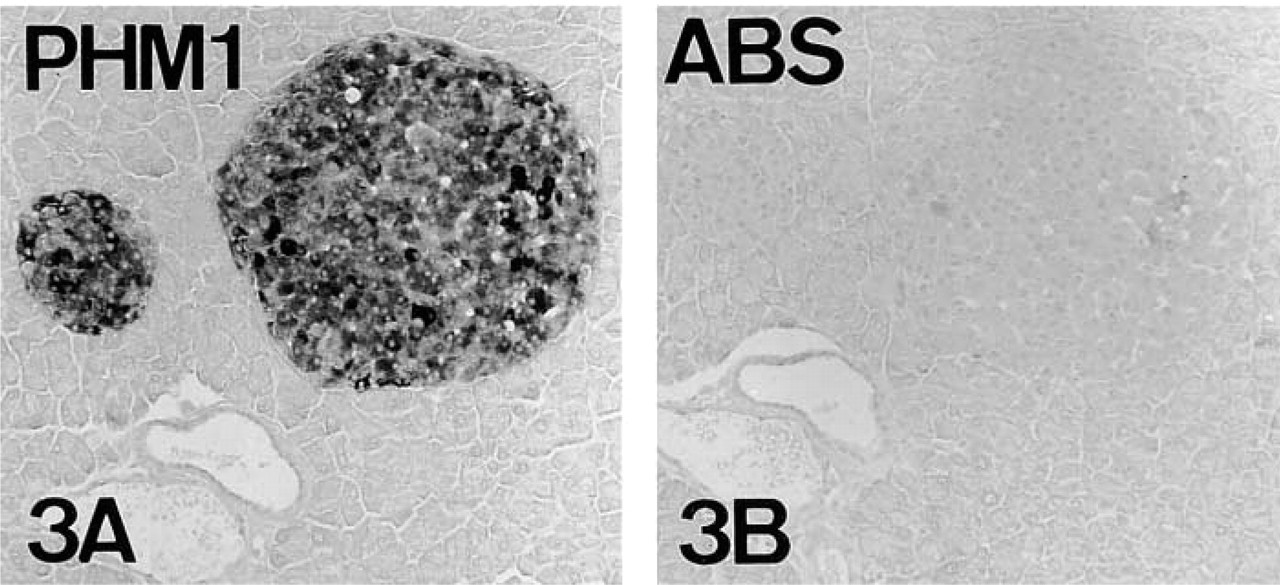
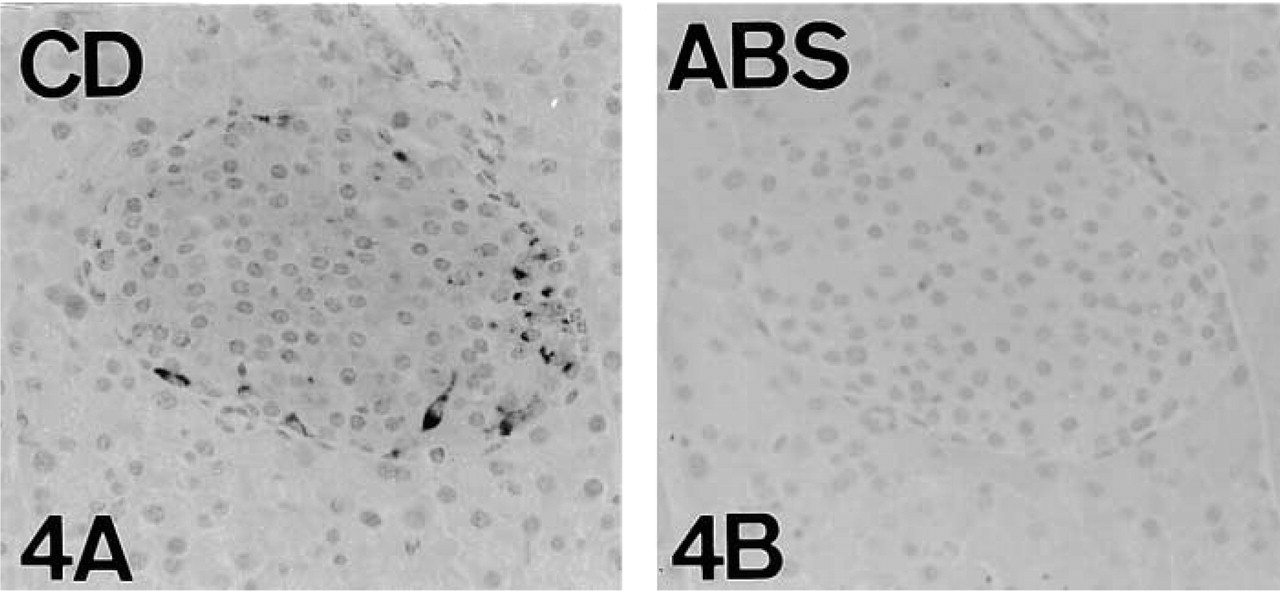
Absorption controls of PAM in serial reversed-face paraffin sections of adult mouse pancreas. PHM (
(
Immunocytochemical staining was performed using the avidin-biotin complex (ABC) (Hsu et al. 1981) and EnVision Plus complex (DAKO; Carpinteria, CA) techniques. Both PAM (Table 1) and various pancreatic and amidated peptides (Table 2) antisera were used. To analyze co-localization of the amidating enzymes and pancreatic hormones, two immunocytochemical methods were employed: the ABC complex technique in serial sections and double immunostaining on the same section using the ABC and alkaline phosphatase-anti-alkaline phosphatase (APAAP; Mason and Sammons 1978) techniques.
ABC Complex Technique
Endogenous peroxidase was blocked by treatment with 3% hydrogen peroxide for 10 min. To avoid nonspecific background, the sections were incubated with normal swine or rabbit serum (1:20; University of Navarra) for 30 min before overnight incubation with the specific primary antibody at 4C. Sections were then incubated for 30 min with biotinylated swine anti-rabbit (K353; Dakopatts, Glostrup, Denmark) or rabbit anti-mouse (K354; Dakopatts) immunoglobulins, followed by incubation with the avidin-biotin–peroxidase complex (K355, Dakopatts) for 30 min. Sections were washed in TBS (Tris buffer 0.05 M, NaCl 0.5 M, pH 7.36) after each incubation. The bound antibodies were visualized with 3–3′ diaminobenzidine tetrahydrochloride (DAB, D-3637; Sigma, St Louis, MO) in sodium acetate/acetic acid 0.1 M pH 5.6, containing 2.5% nickel ammonium sulfate, 0.2% β-
EnVision Complex Technique
Endogenous peroxidase was blocked with the DAKO EnVision system reagent 1. Primary antibody was added and incubated overnight at 4C. Sections were rinsed in TBS and incubated for 30 min with the labeled polymer (HRP, reagent 2). The bound antibodies were visualized as explained for the ABC method.
Double Immunostaining
Sections were incubated overnight with a mixture of the two primary antibodies. The second layer consisted of a mixture of biotinylated swine anti-rabbit and non-biotinylated goat anti-mouse immunoglobulins. After rinsing in TBS, optimally diluted ABC and APAAP complexes were added for 30 min and then re-incubated in non-biotinylated rabbit anti-mouse immunoglobulins and APAAP complexes for 10 more min each. The APAAP bound antibodies were visualized in red with previously hexazoted new fuchsin (Sigma; 638) in 4% HCl 2 M and 1 mg/ml naphthol solution in 0.2 M Tris-HCl (pH 9.2). The ABC bound antibodies were visualized in black as we have previously described, after rinsing the sections in water and sodium acetate/acetic acid 0.1 M, pH 5.6. The sections were finally mounted in PBS:glycerol (1:1).
Microwave Pretreatment
When required for antigen retrieval, sections were incubated for 15 min in citrate buffer 0.01 M, pH 6.0, at full microwave heating power, followed by a second incubation in the same buffer at half power (Tables 1 and 2).
Specificity Controls
Human and rat pancreata were used as positive controls. Absorption controls for PAM and pancreatic peptide-immunoreactive antibodies were performed in mouse. In human and rat, absorption tests for PAM antisera were also carried out. Antisera that rendered immunoreaction were preincubated overnight with their respective synthetic antigens (Table 3). Preabsorbed antibodies were then used for immunocytochemistry.
Western Blotting
Pancreata were homogenized in a buffer containing 10 mM Tris (pH 7.4), 150 mM NaCl, 1% Triton X-100, 1% deoxicholate, 0.1% SDS, and 5 mM EDTA. Tissue final protein concentration was determined (BCA kit; Pierce, Rockford, IL) after ultracentrifugation. Thirty μg protein was heated to 70C for 10 min and loaded into the sample well. Tissue protein extracts were electrophoretically separated on a gradient Nu PAGE 3–8% Tris-acetate gel (Novex; San Diego, CA) and run at 150 V for 1 hr under reducing (5% β-mercaptoethanol) and non-reducing conditions. Transfer blotting was accomplished in the same apparatus equipped with a titanium plate electrode and transferred to a polyvinyldifluoride membrane (Inmovilon PVDF; Millipore, Bedford, MA) at 30 V for 1 hr. The blots were saturated with 5% (w/v) non-fat milk in PBS for 1 hr and incubated overnight in a 1:1000 dilution of rabbit anti-PHM or rabbit anti-CD in PBS. After washing three times in PBS-Tween, they were incubated for 1 hr with peroxidase-conjugated anti-rabbit or anti-mouse antibody at 1:5000 (Amersham Life Science; Arlington Heights, IL) and washed again in PBS-Tween. The immune complexes were detected with Lumi-Light Plus Western Blotting substrate (Roche Diagnostics; Indianapolis, IN) according to the supplier's instructions.
Results
Distribution of PAM IR in Mouse Pancreas
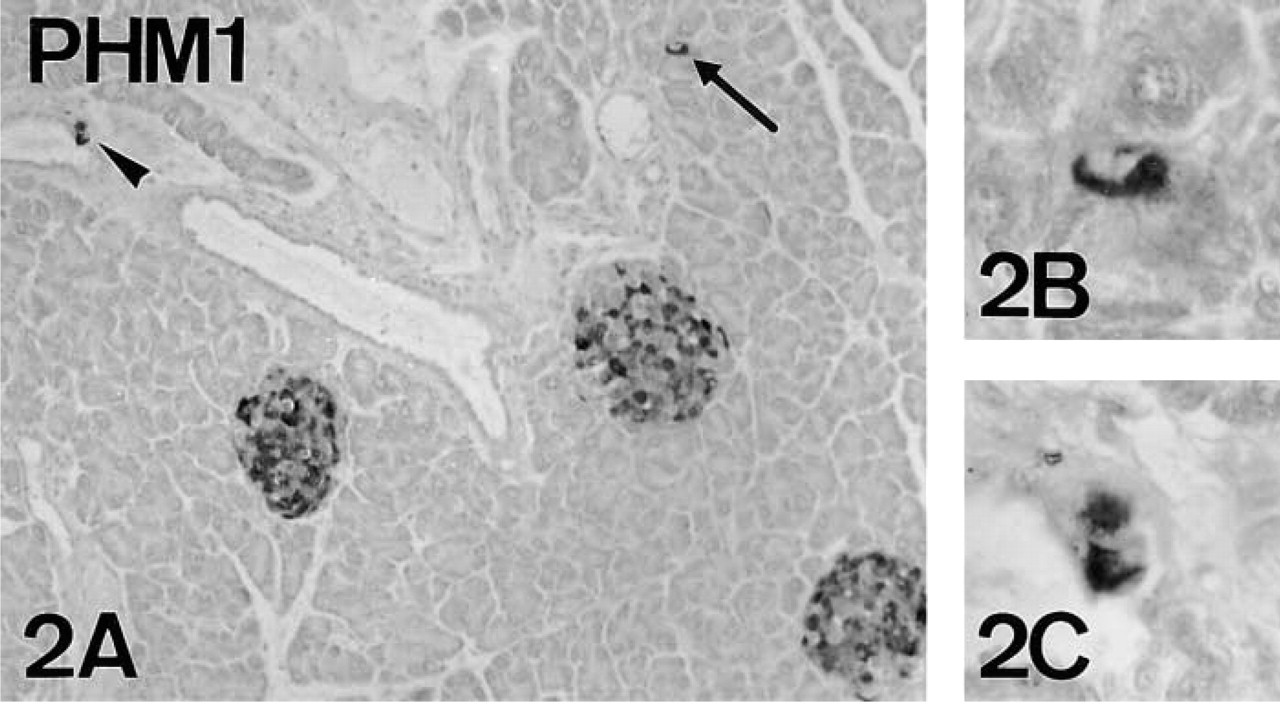
PAM-IR was abundant in the pancreas of adult mice. Immunolabeling was observed mostly in islet cells, but immunostained cells were also found scattered within acini and ducts (Figure 2).
Antisera and peptides used in absorption controls
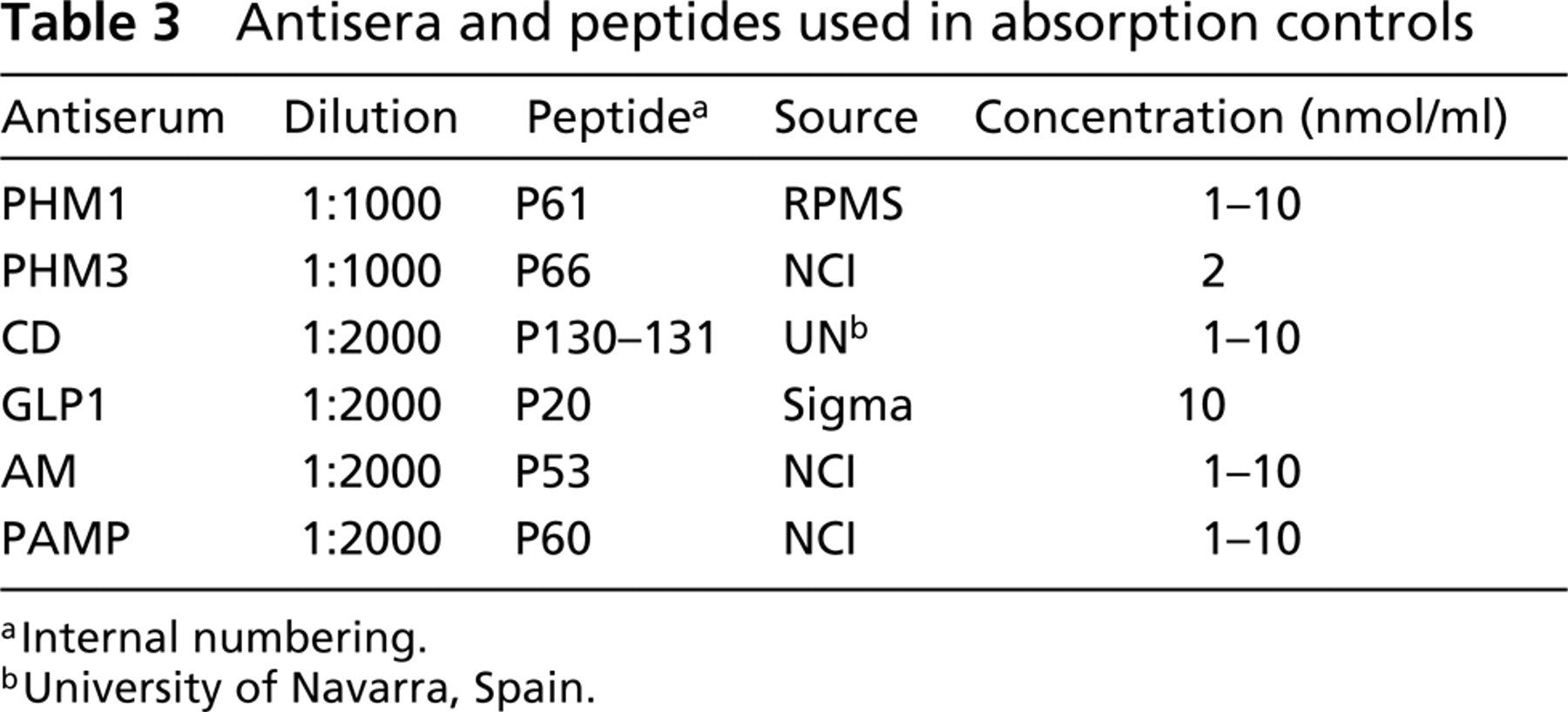
Internal numbering.
University of Navarra, Spain.
(
Human pancreas. PHM1 immunostaining (
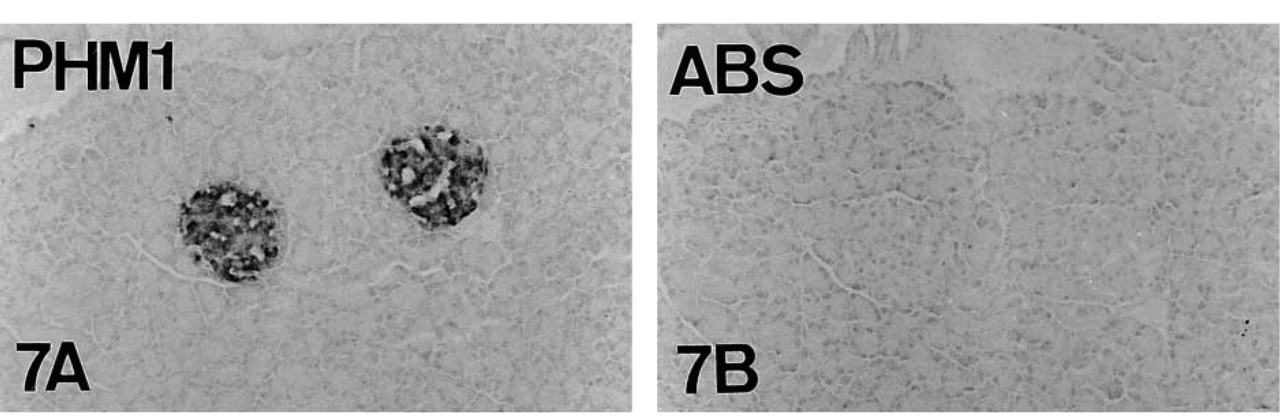
(
Rat pancreas. PHM1-immunostaining (
PHM-IR (
PHM-IR (
Immunostaining was obtained with antisera raised against the PHM enzyme region and the hydrophilic cytoplasmic domain of PAM (Table 1; Figures 3A and 4A). As will be made clear, immunolabeling for the cytoplasmic domain of PAM was variable according to the method of fixation. Preabsorption of the immunoreactive antisera against PHM and the cytoplasmic domain of PAM with their corresponding antigens (Table 3) abolished immunoreactivity (Figures 3 and 4). Although IR was also observed with the antisera against the PAL domain (data not shown), absorption tests did not confirm the specificity of immunoreaction.
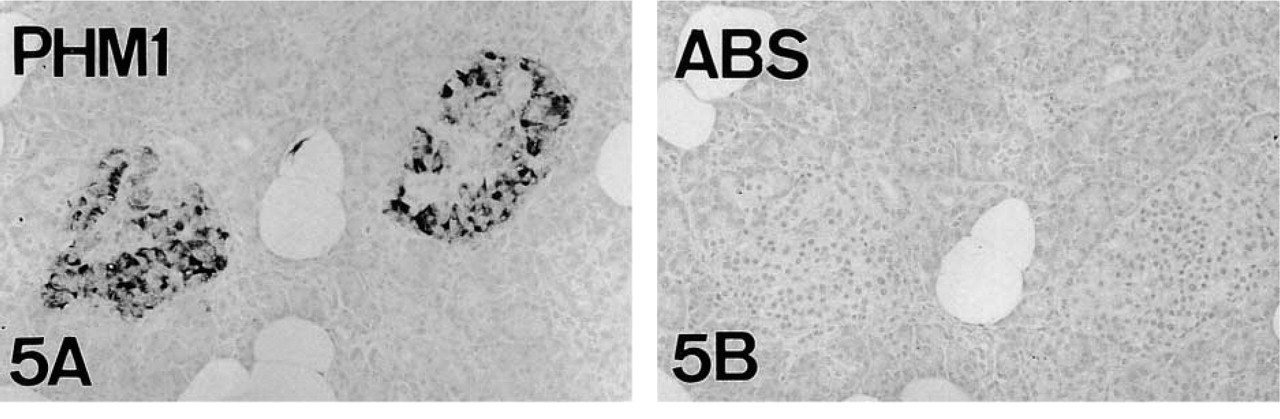
Human and rat pancreata, used as positive controls, also displayed abundant PAM-IR (Figures 5–8). Preabsorption tests confirmed the specificity of staining (Figures 5 and 7).
Identification of the PAM-producing Cell Types
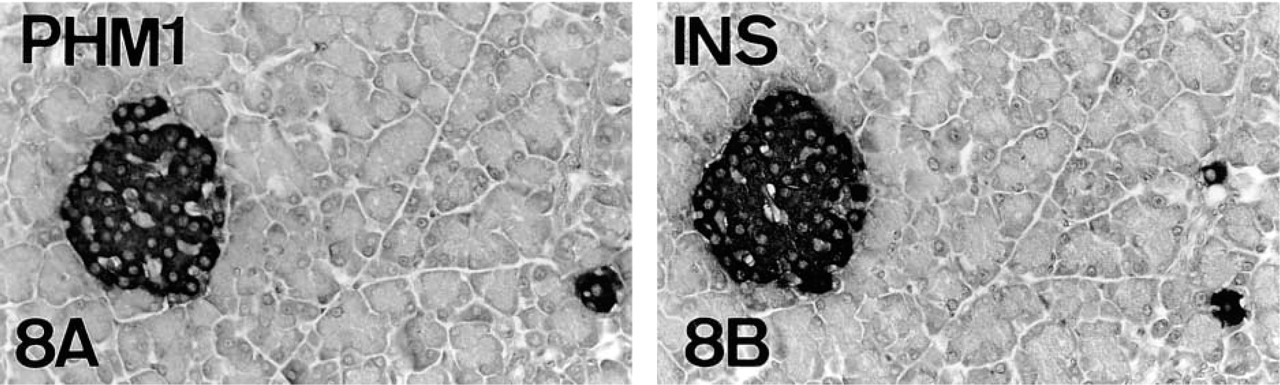
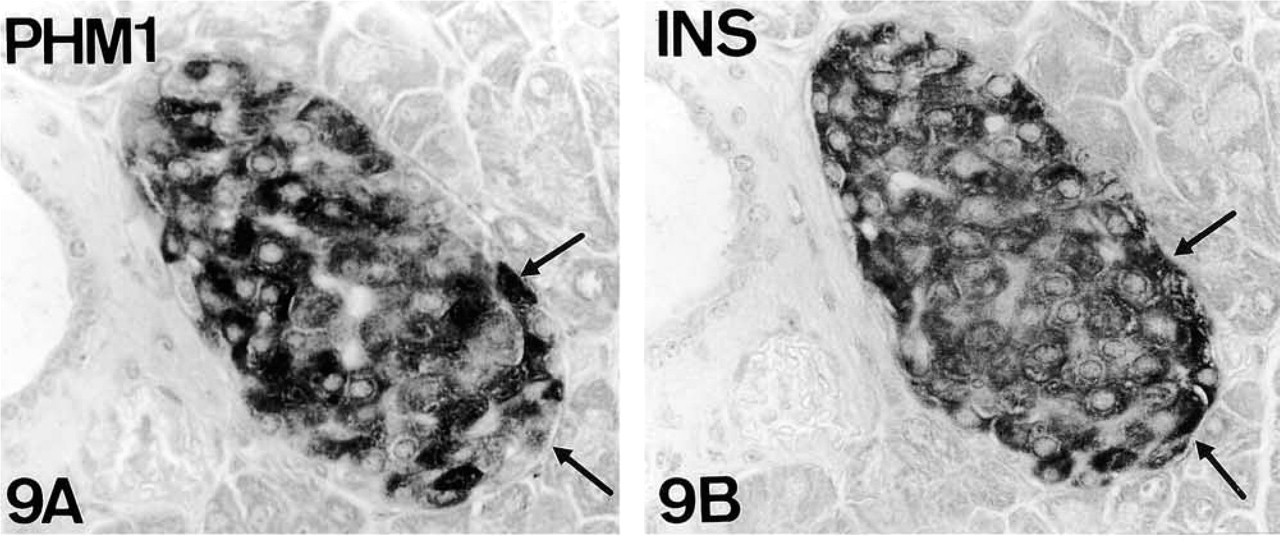
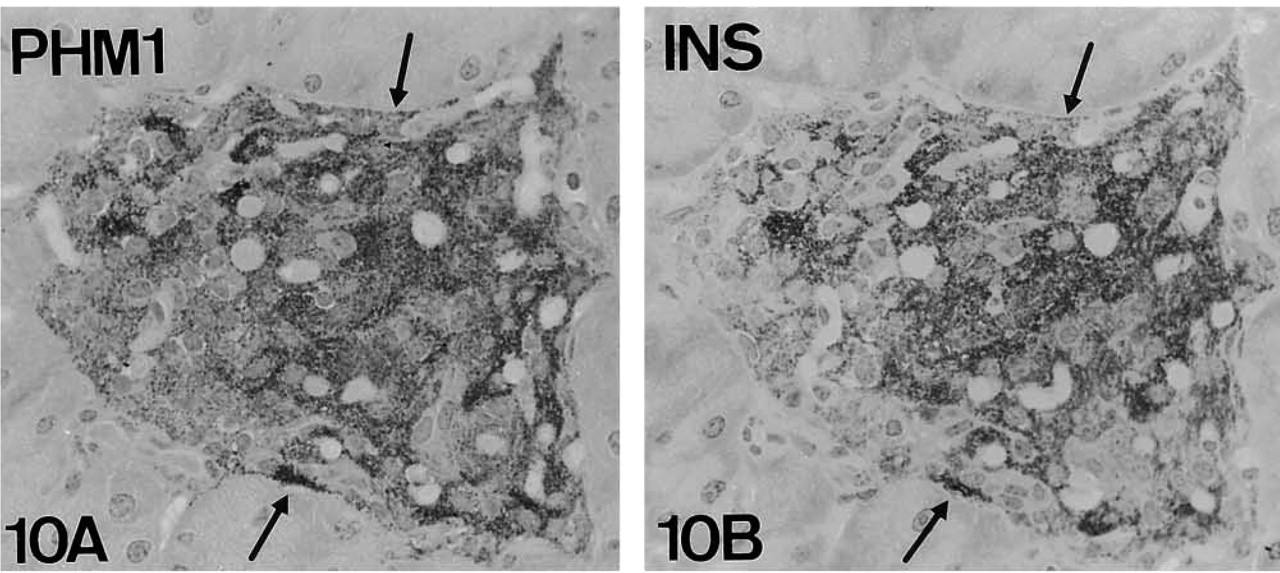
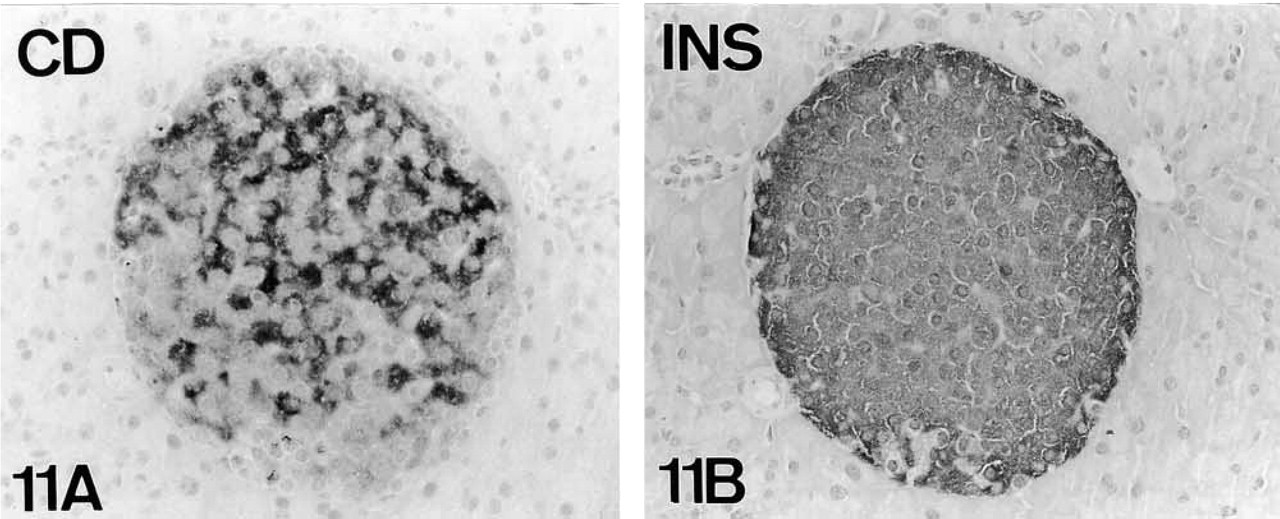
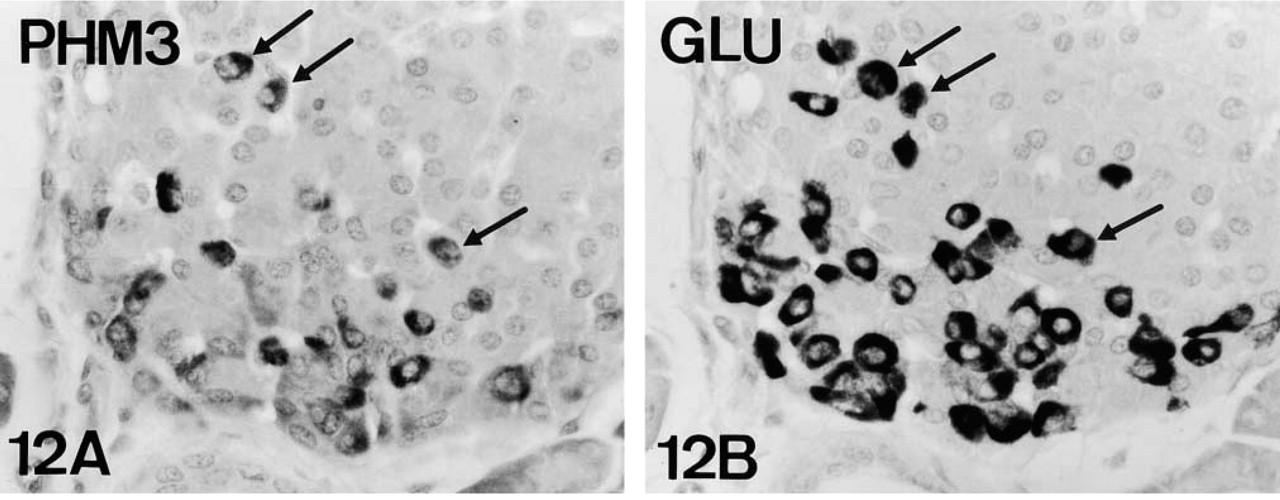
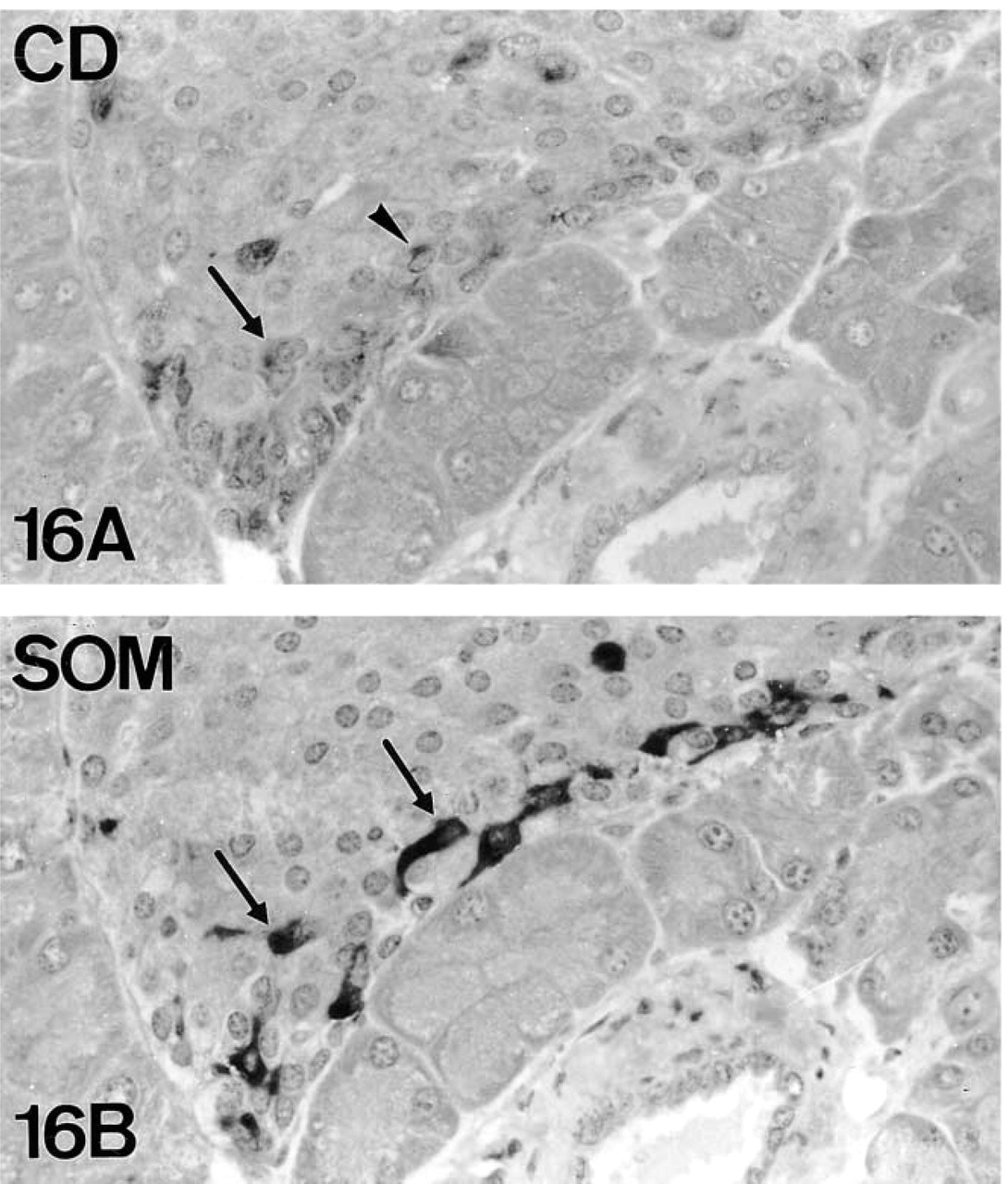
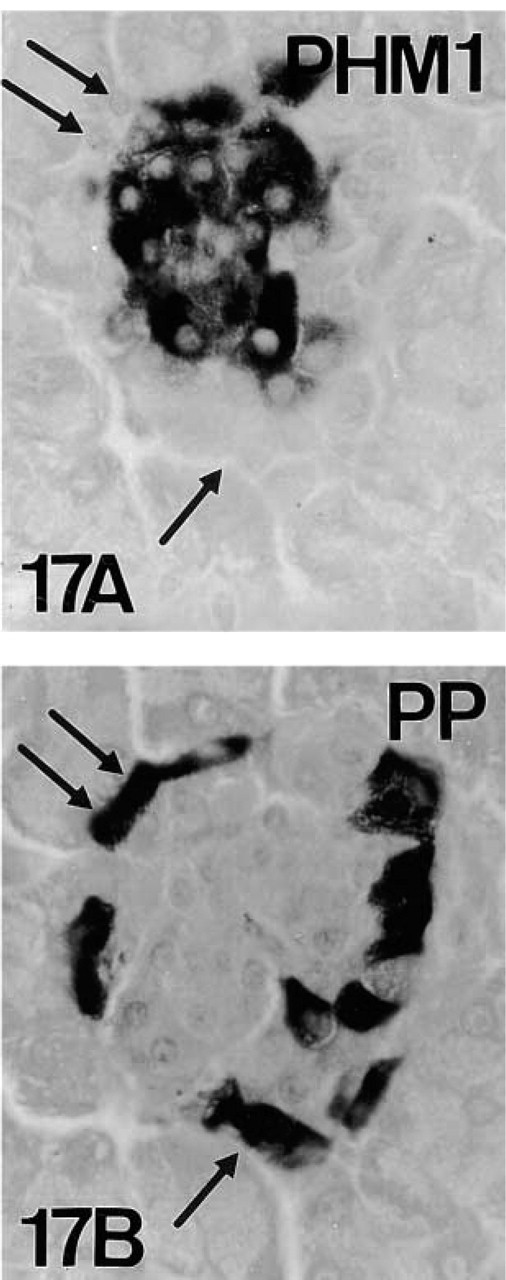
In mouse pancreas, co-localization studies of PAM and the classical pancreatic regulatory peptides showed that three cell types [B-cells (insulin), A-cells (glucagon), and D-cells (somatostatin)] displayed PAM-IR. For D-cells, immunoreaction was found only in a few specimens. Co-localization of the three peptides and PAM-IR was observed in serial sections of both paraffin- and plastic-embedded material (B-cells, Figures 9–11; A-cells, Figures 12–14; D-cells, Figures 11 and 12) and by double immunolabeling techniques (Figures 18–20). On the contrary, no IR for PAM was found in PP cells (Figure 17).
Most, if not all, mouse B-cells were positive for PAM (Figures 9–11 and 14), although with different degrees of intensity (Figure 9). When obtained, PAM-IR was also present in most D-cells (Figure 15). On the contrary, only a subpopulation of A-cells was PAM-immunoreactive (Figure 12).
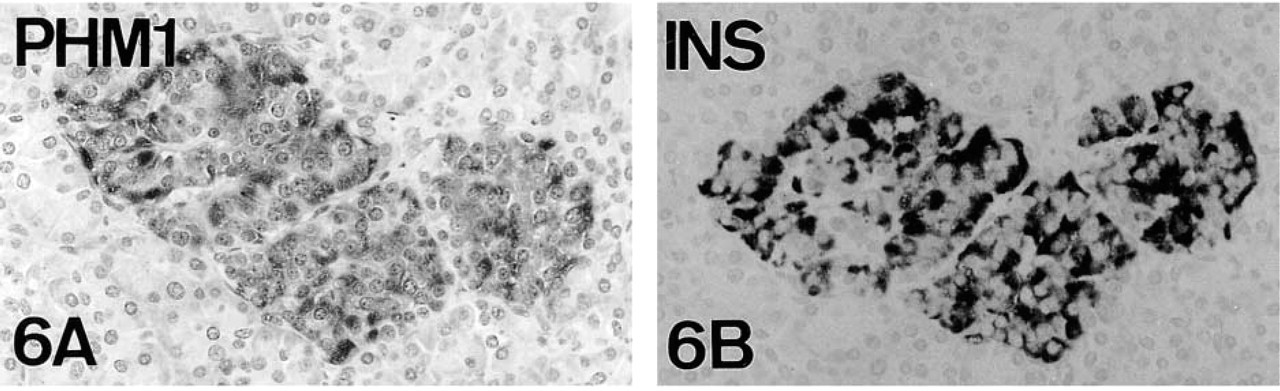
In human and rat pancreata, in addition to the expected PAM IR in A-cells, immunostaining was also demonstrated in B-cells (Figures 6 and 8).
Cellular Distribution and Pattern of PAM Staining
PAM Staining Pattern of Mouse A-, B-, and D-cells. Mouse endocrine cells positive for PAM were not immunoreactive for the same PHM antisera (Table 4). In addition, differences concerning the cellular distribution of CD staining were also observed.
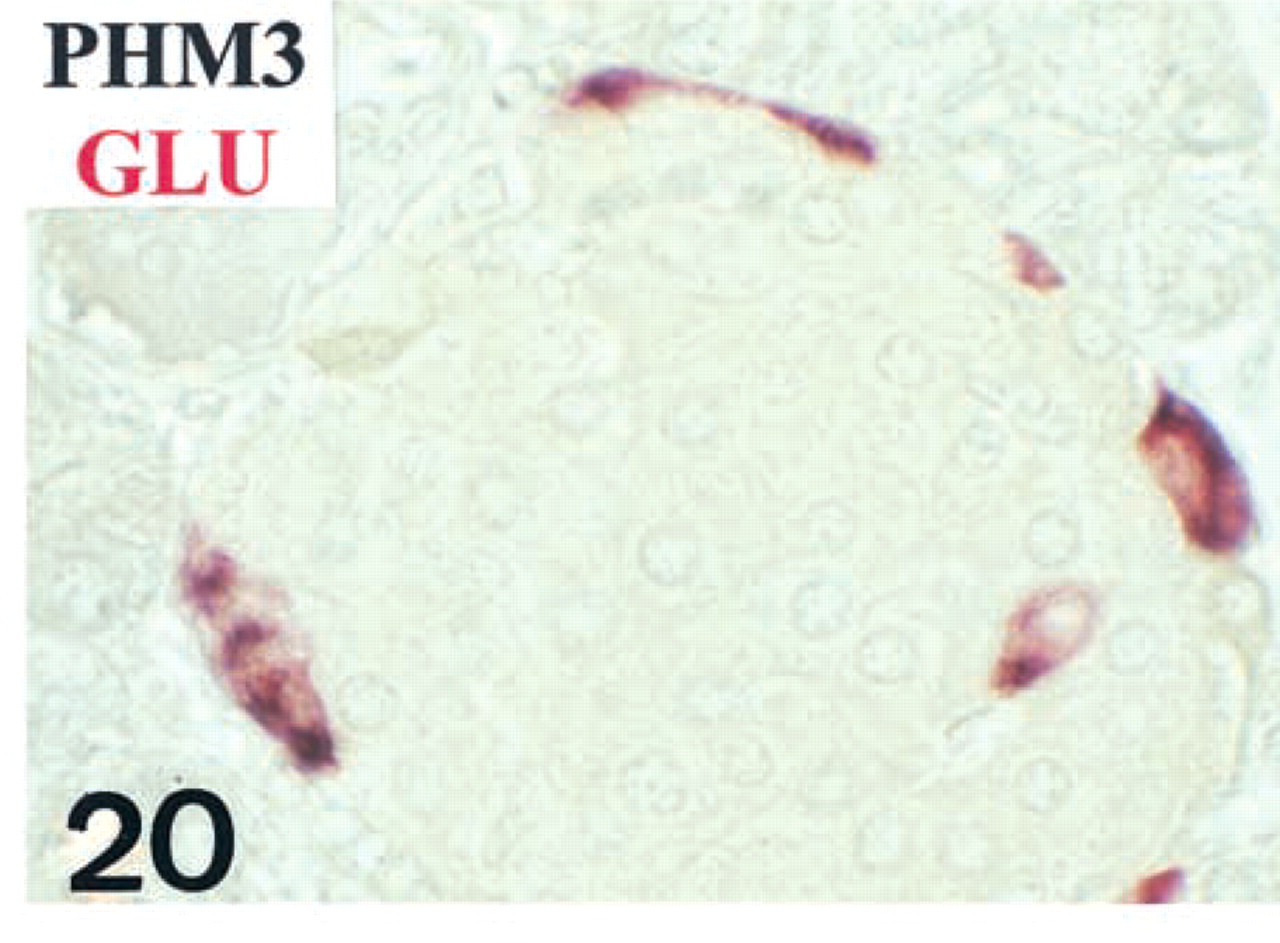
PHM Staining. In the three cell types, PHM immunolabeling extended throughout the cell (Table 5). Unexpectedly, although A-, B-, and D-cells were stained with PHM antisera, the three cell types did not show the same IR pattern. A-cells were labeled with the antiserum PHM3 but not with PHM1, whereas B- and D-cells showed the opposite IR pattern, (PHM1+/PHM3-) (Figures 1 and 20). Such staining patterns did not change with different processing procedures. As discussed below, the existence of different molecular forms of PAM in mouse A- and B-cells might explain the differences in immunoreaction.
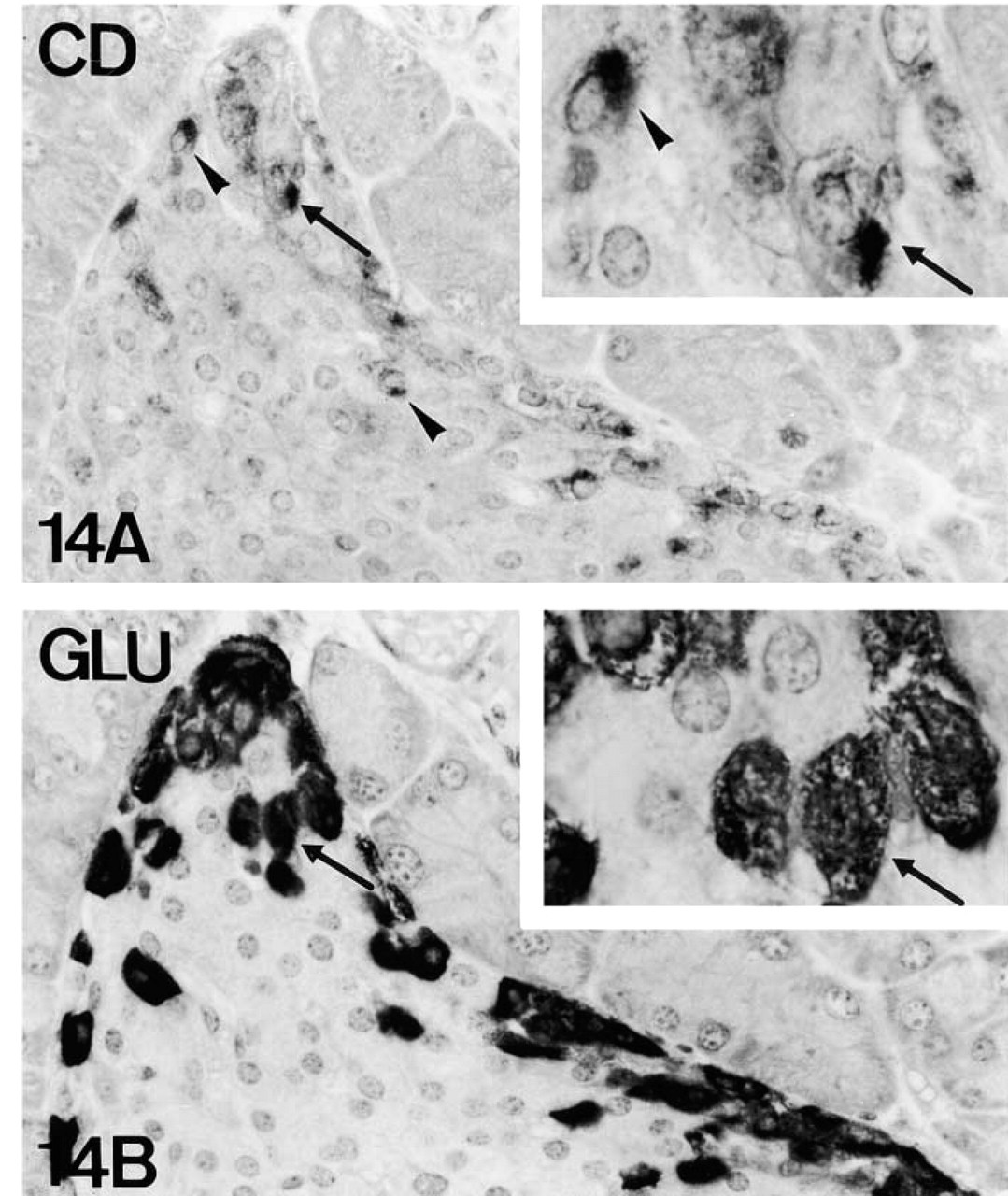
Cytoplasmic Domain Staining. In addition to extended PHM-IR, A- and D-cells showed cytoplasmic areas immunoreactive for the cytoplasmic domain of PAM near the nucleus (Figures 14 and 16). Both positive and negative A- and D-cells were observed for this antiserum, but bearing this localized staining pattern in mind, the lack of IR in some A/D-cells might be due to sectioning artifact. Whereas in A- and D-cells immunolabeling was obtained with all fixatives used, B-cells rendered only weak IR for the cytoplasmic domain of PAM in Zamboni-fixed material. In this case, the staining pattern was not juxtanuclear but extended throughout the cytoplasm of B-cells (Figure 11; Tables 4 and 5).
PHM Western Blotting. Pancreatic extracts were analyzed by Western blotting using both PHM1, PHM3, and cytoplasmic domain antisera (Figure 23). Western blotting analysis revealed immunoreactive bands from about 47 to 96 kD. Most of the mouse bands were immunoreactive for a single PHM antiserum: PHM1 (three bands of 62, 66 and 74 kD) or PHM3 (four bands of 53, 57, 60, and 96 kD). Only a 50-kD band displayed immunoreactivity for the two PHM antisera. Nevertheless, Western blotting does not discriminate whether such immunoreactive bands correspond to the same molecule or to different proteins of the same weight. In relation to CD, a band of approximately 46 kD was also immunoreactive for CD. PHM3 bands of 57 and 96 kD also seem to coincide in weight with a CD-immunoreactive band.
Co-localization of PAM and Amidated Peptides
Because neither glucagon, insulin, or somatostatin are amidated, some amidated peptides (Table 2) were immunocytochemically investigated to determine the possible functional significance of PAM enzymes in mouse A-, B-, and D-cells. The study revealed that, in the three cell types, at least one amidated peptide is present: GLP1 and AM in A-cells, AM and PAMP in B-cells, and AM in D-cells (Figures 21, 22, and 24). These amidated peptides co-localize not only with the corresponding classical pancreatic peptides but also with the amidating enzymes (Figures 21 and 25). In the case of the amidated peptide AM, we found different IR intensities throughout the AM+ population.
PAM-IR in mouse A-cells (arrows). PHM-IR (Figures 12A and 13A) and cytoplasmic domain-IR (Figure 14A) are present in a subpopulation of glucagon-IR A-cells (Figures 12B, 13B and 14B). Note that cytoplasmic domain-IR is restricted to a juxtanuclear area (arrowheads, Figure 14A). Figures 12 and 14, serial reversed-face paraffin sections. Original magnification X500, X1250. Figure 13, serial plastic sections. Original magnification X500.
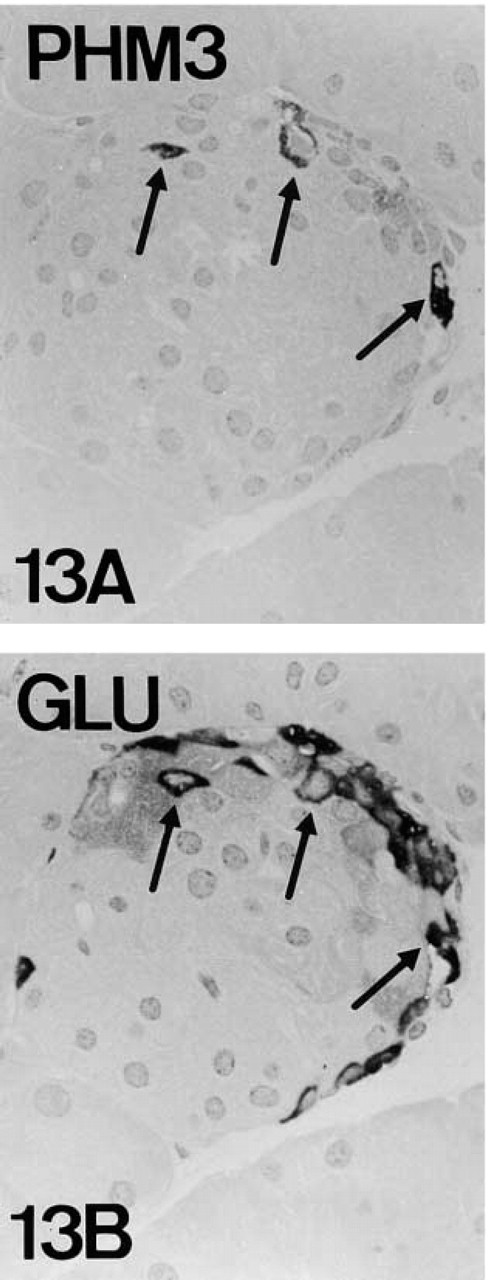
PAM-IR in mouse A-cells (arrows). PHM-IR (Figures 12A and 13A) and cytoplasmic domain-IR (Figure 14A) are present in a subpopulation of glucagon-IR A-cells (Figures 12B, 13B and 14B). Note that cytoplasmic domain-IR is restricted to a juxtanuclear area (arrowheads, Figure 14A). Figures 12 and 14, serial reversed-face paraffin sections. Original magnification X500, X1250. Figure 13, serial plastic sections. Original magnification X500.
PAM-IR in mouse A-cells (arrows). PHM-IR (Figures 12A and 13A) and cytoplasmic domain-IR (Figure 14A) are present in a subpopulation of glucagon-IR A-cells (Figures 12B, 13B and 14B). Note that cytoplasmic domain-IR is restricted to a juxtanuclear area (arrowheads, Figure 14A). Figures 12 and 14, serial reversed-face paraffin sections. Original magnification X500, X1250. Figure 13, serial plastic sections. Original magnification X500.
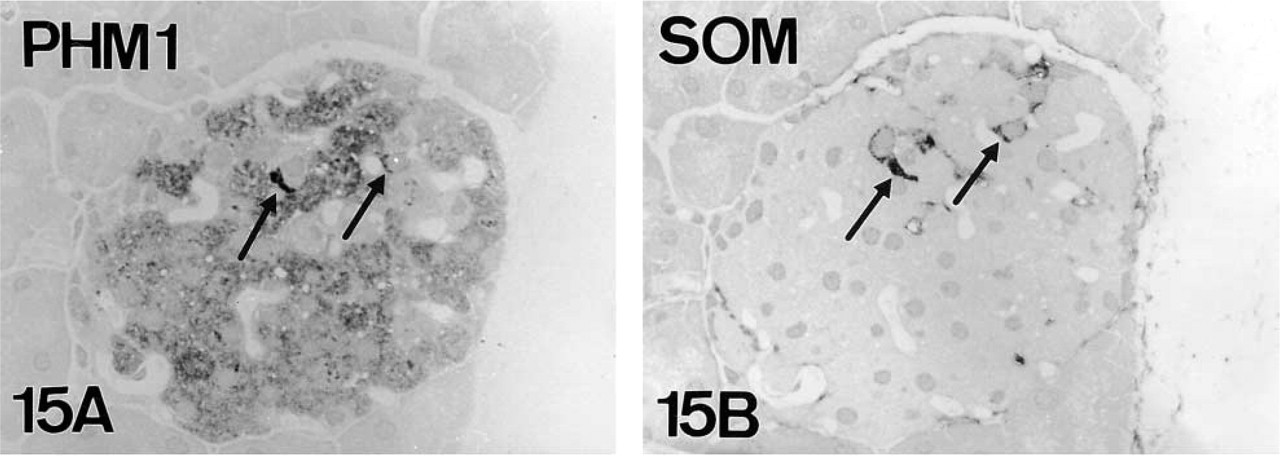
PHM-IR in somatostatin-immunoreactive D-cells (arrows). Serial plastic semithin sections. Original magnification X500.
Cytoplasmic domain-IR (
PHM (
PAM immunoreactivity pattern in mouse islet cells

Both + and − cells exist.
Negative D-cells may be due to section artifact (see text).
Only in Zamboni-fixed tissue.
Only in some specimens.
Discussion
Presence of PAM in Mammalian Adult Pancreas
Studies about the presence of PAM in mammalian pancreas, both neonatal and adult, have been carried out in very few species, with no agreement on its presence in adult specimens. The present study reports the existence of PAM in adults of a third mammalian species (mouse) by both immunocytochemistry and Western blotting. Although the present report focuses mainly on mouse pancreas, we also show the presence of amidating enzymes in more pancreatic mammalian cell types than were previously described in human and rat.
In concordance with previous immunocytochemical studies in human and rat (Braas et al. 1992; Martinez et al. 1993b), our results show that in mouse the enzyme PAM is produced by the pancreatic endocrine tissue of adults. Nevertheless, whereas in previous studies of human and rat few PAM-immunoreactive cells were reported, in the present study PAM immunolabeling was demonstrated in many cells of mouse, human, and rat endocrine pancreas. Such differences are probably due to the different antisera used (see below). The abundance of pancreatic PAM-immunoreactive cells appears to contradict the low levels of PAM mRNA reported in adult rat pancreas (Maltese et al. 1989). However, both findings might reflect a shorter lifespan of PAM mRNA compared with the translated enzyme, which would remain in the secretory granules for a longer period. In mouse, PAM-IR is predominantly observed at the islets of Langerhans, and only occasional scattered immunostained cells within acini and ducts have been found.
Identification of the PAM-producing Pancreatic Cell Types
As indicated, previous immunocytochemical studies localized the enzyme PAM in human A-cells (Martinez et al. 1993b) and in rat “peripheral” (A/PP) islet cells (Braas et al. 1992). We have equally demonstrated the presence of PAM in A-cells of the mouse. However, in partial disagreement with such previous studies, we have identified the enzyme not only in mouse A-cells, but also in B- and D-cells. In fact, concerning the presence of PAM in mammalian B-cells, one of our antisera (PHM1) also rendered immunostaining in B cells of both human and rat pancreata (Figures 6 and 8). This difference in present and previous immunocytochemical results (Braas et al. 1992; Martinez et al. 1993b) is probably because of the different antisera used. In fact, although biochemical studies do not identify clearly the PAM-producing cell type(s), they also suggest that B-cells could be responsible for PAM activity in rat pancreas (Ouafik et al. 1987; Maltese et al. 1989).
As occurs in human pancreas (Martinez et al. 1993b), PP cells of mouse lacked PAM staining. This result was unexpected because PP is the only amidated peptide out of the four classic pancreatic hormones. However, mouse A-, B-, and D-cells do contain other amidated peptides. The absence of IR in PP-producing cells could be explained by amidation taking place in blood by PAM after secretion, as has already been suggested (Martinez and Treston 1996) or by the existence of another PAM molecular form that could not be detected with our antisera. The existence of a different, yet-undescribed amidating enzyme has also been proposed as an explanation for the lack of PAM enzymes in this cell type (Martinez and Treston 1996).
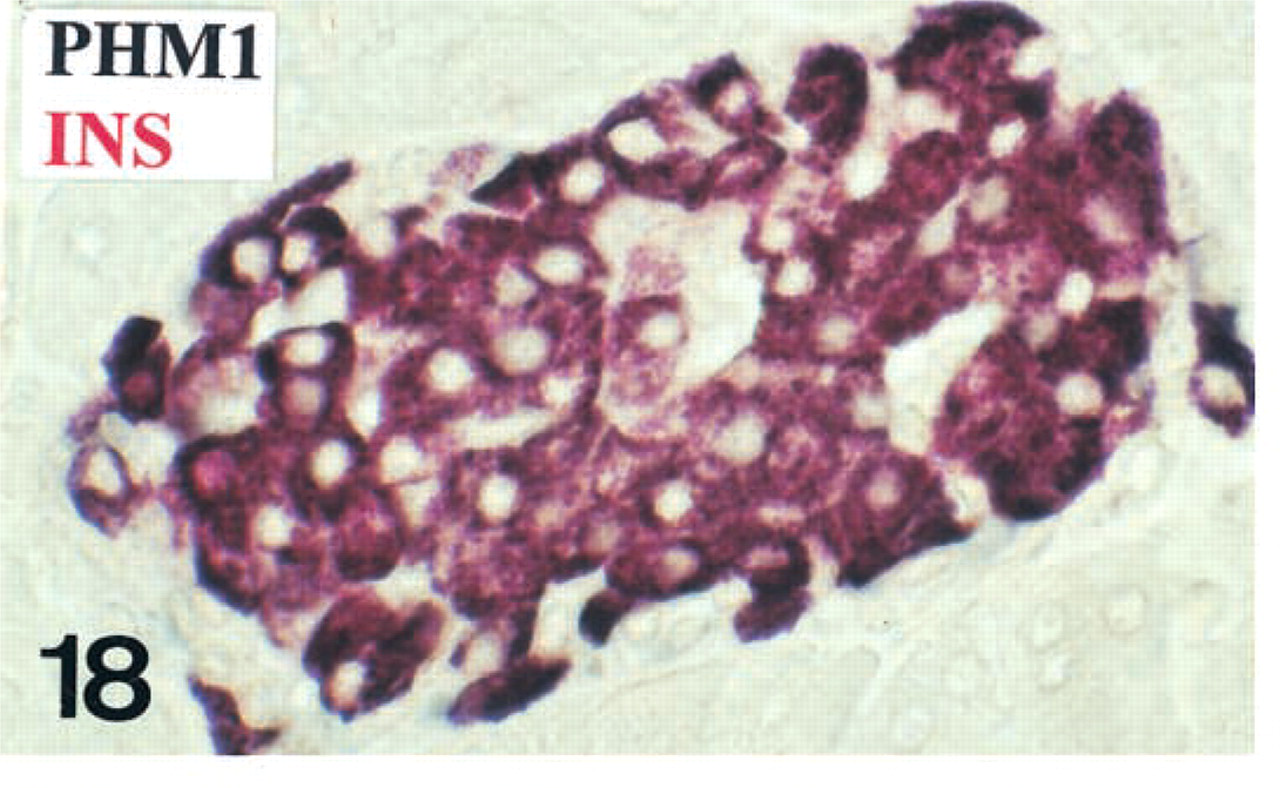
B-cells (purple) displaying both PHM-1 (black) and insulin (red)-IR
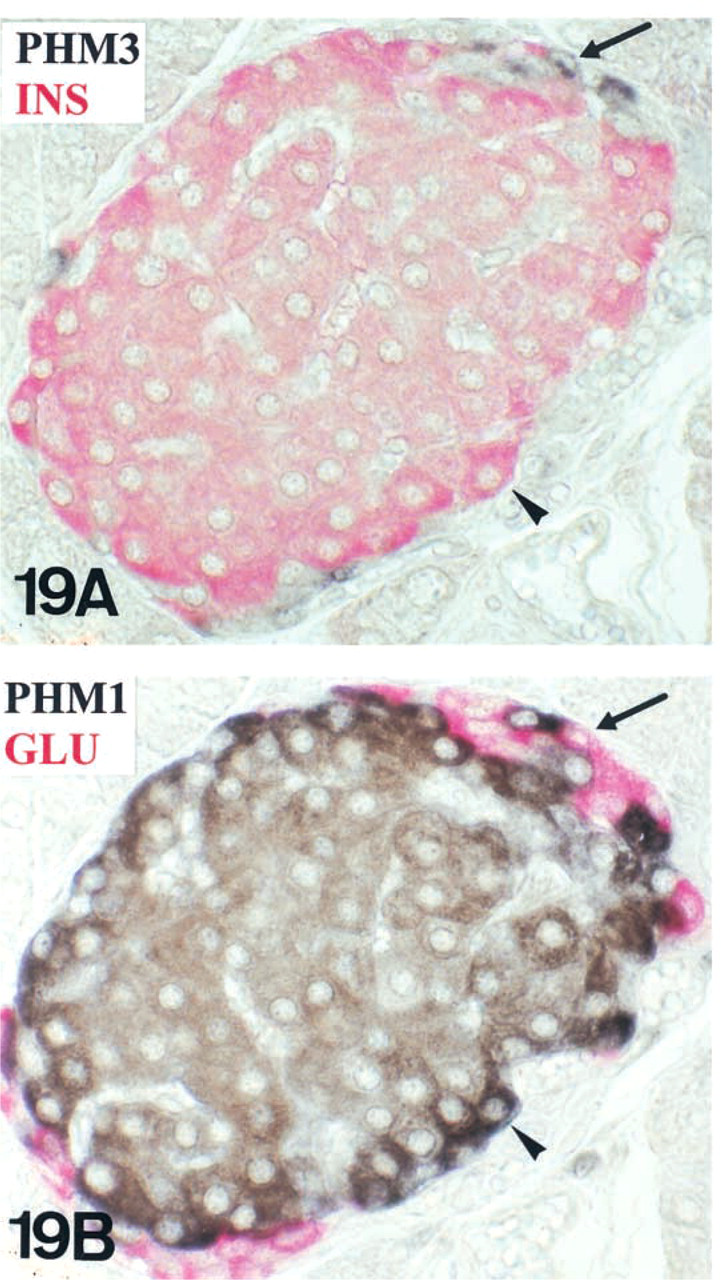
Double IHC was performed in serial reversed-face paraffin sections to demonstrate different IR patterns for PHM antisera of mouse glucagon (arrow) and insulin (arrowhead) cells. Insulin-IR cells (red in
A-cells (purple) display both PHM3 (black) and glucagon (red) IR.
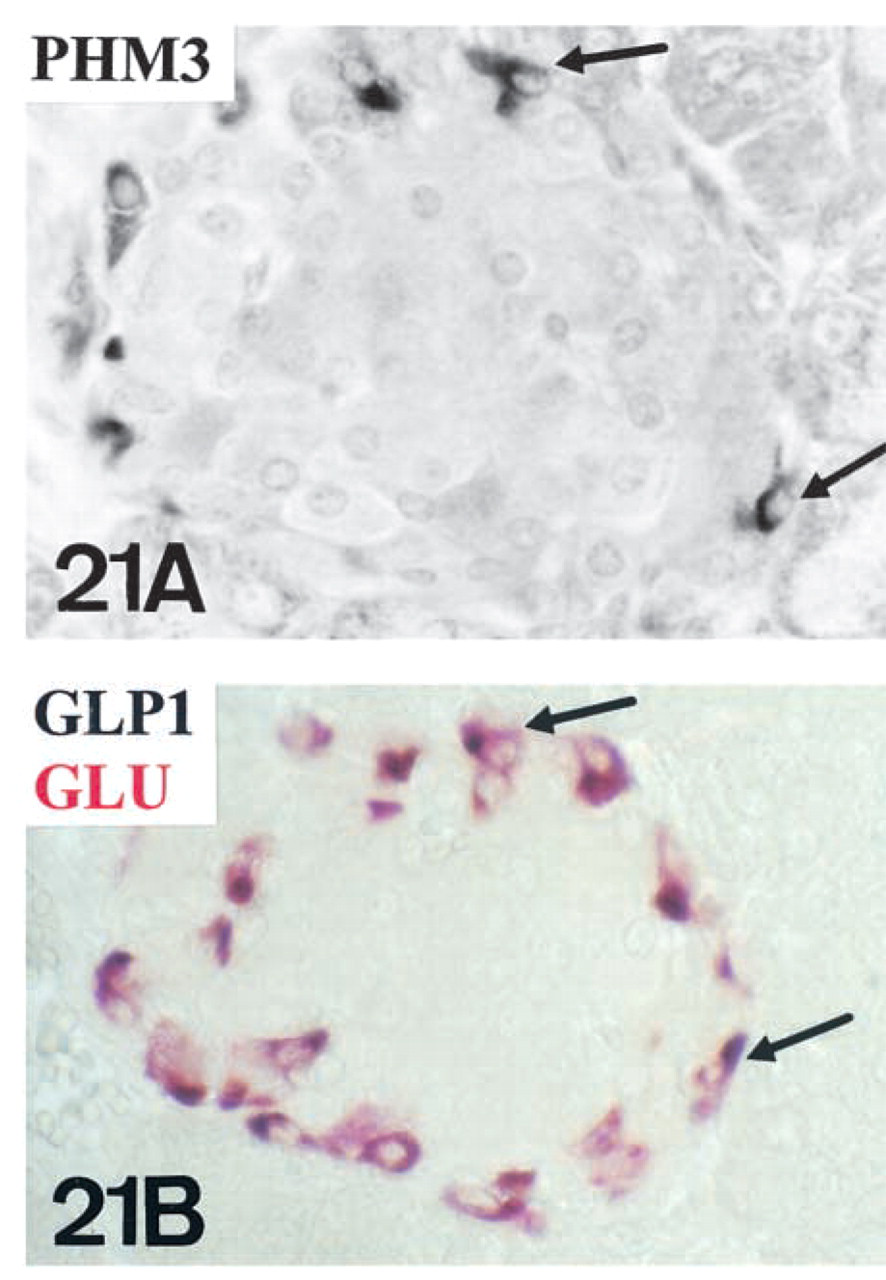
In A-cells (purple in
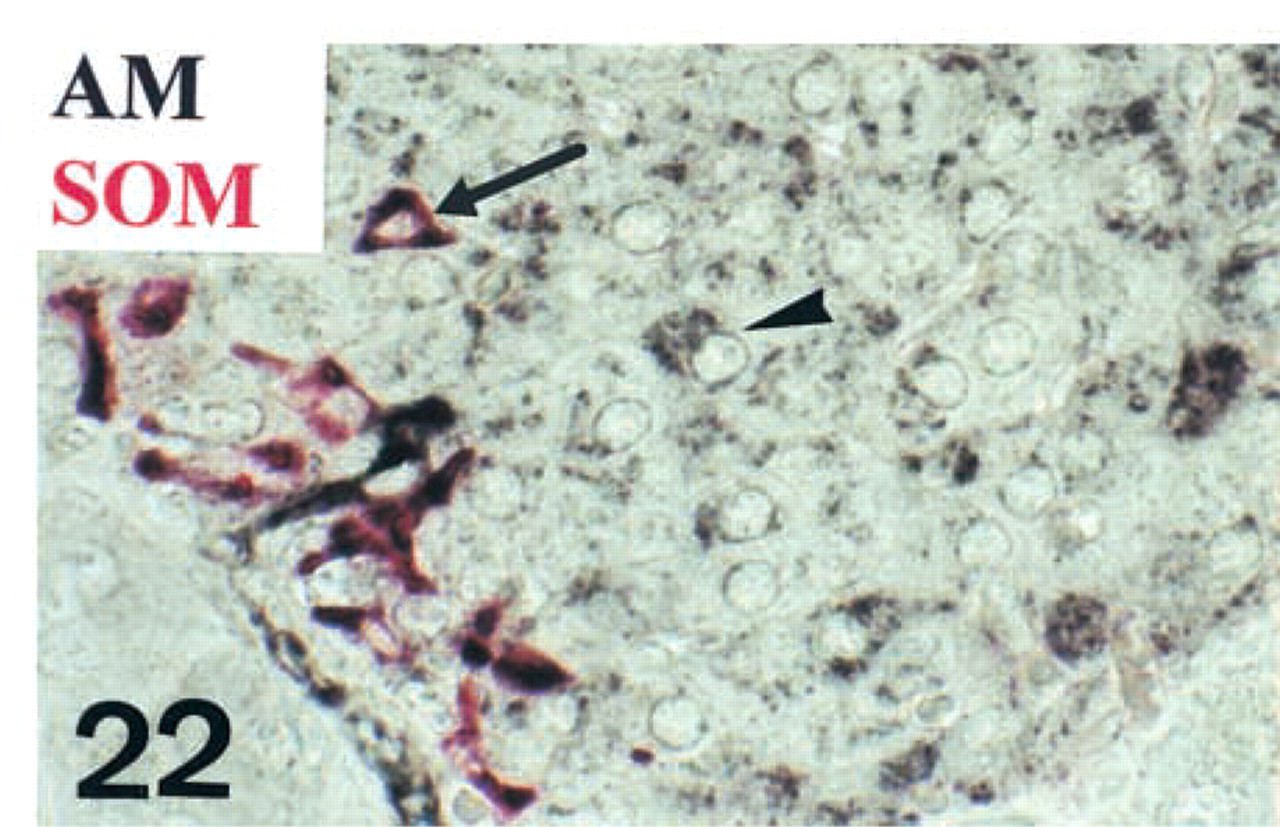
Some D-cells (arrow) immunoreact for both somatostatin (red) and AM (black). Other types of AM-producing cells are also present (arrowhead). Double immunohistochemistry in paraffin. Original magnification X500.
PAM-IR depending on processing methods

Results in paraffin; antiserum against the cytoplasmic domain of PAM did not render IR in plastic sections.
Subpopulations of PAM-producing Mouse Pancreatic Endocrine Cells
The enzyme PAM has been identified in less than half of mouse A-cells. Conversely, most, if not all, B- and probably D-cells appear to be immunoreactive for PAM antisera, although with different degrees of intensity. A similar description of PAM-producing subpopulations was reported in pituitary by Steel et al. (1994).
The existence of a PAM-immunoreactive A-cell subpopulation does not necessarily imply that the rest of the A-cells cannot synthesize the enzyme. PAM might be produced in certain functional moments, responding to nutritional or other physiological situations. This might also account for the absence of PAM in D-cells of certain specimens. In the case of B-cells, there appears to be basal production of PAM in the entire population, but the differences observed in the intensity of staining might also be related to higher or lower enzymatic production phases. Therefore, in mouse pancreatic endocrine cells, functional PAM-producing stages may be suggested, which would give rise to dynamic, changing subpopulations. Such PAM-producing stages may be related to the production of amidated peptides, as is suggested by the different degree of intensities found in IR for the amidated peptide adrenomedullin (Figures 22 and 25).
Diversity of PAM Enzymes in Mouse Endocrine Pancreatic Cell Types
Immunocytochemistry and Western blotting analysis have suggested the existence of different PAM molecules in mouse endocrine pancreas. Immunocytochemistry detected specific IR for PHM and for the cytoplasmic domain of PAM. However, PAL immunoreactivity is uncertain because preabsorbed PAL antisera still stained the cells. Therefore, further biochemical assays are needed to ascertain the presence of monofunctional (only PHM) or bifunctional (PHM/PAL) amidating enzymes in mouse pancreas. Nevertheless, Western blotting analysis showed the existence of an immunoreactive band around 96 kD (Figure 23), a size large enough to correspond to a bifunctional amidating enzyme estimated to be at least 75 kD (Eipper et al. 1992a, b). The smaller forms should correspond to monofunctional PHM proteins. In the case of the 74-kD PHM1-IR band, it could also account for a bifunctional enzyme, because molecular weights are only estimates.
Immunocytochemistry and Western blotting also showed the existence of CD-IR in mouse pancreas, suggesting the existence of membrane-bound molecules. According to molecular weights, CD-IR bands could correspond either to monofunctional or bifunctional enzymes. In addition, soluble molecules (lacking CD) also appear to exist.
The existence of different molecular forms of PAM enzymes in mouse pancreatic islets could involve different catalytic properties of each PAM molecular form, e.g., the specificity for different substrates, as has been previously described in Lymnaea (Spijker et al. 1999).
Differences in PHM Immunostaining Pattern of Pancreatic Endocrine Cell Types
PHM-IR was observed in mouse A-, B-, and D-cells with all the technical procedures. However, the IR to different PHM antisera found (PHM3+/PHM1- in A-cells; PHM1+/PHM3- in B- and D-cells; Table 4) suggests the existence of different PAM molecular forms in insular cells, which would be recognized by different antisera.
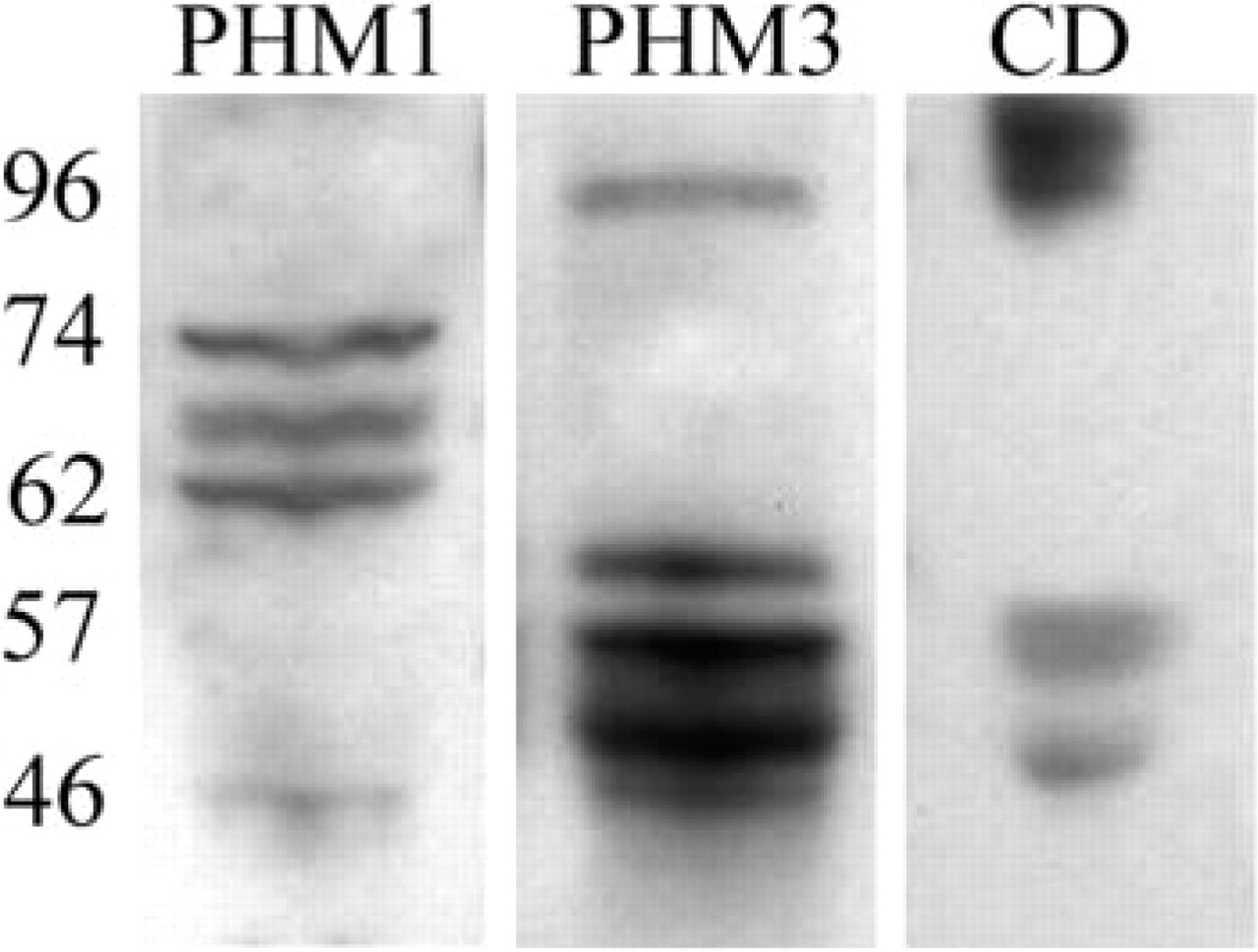
PHM1, PHM3, and CD Western blotting in mouse pancreas.
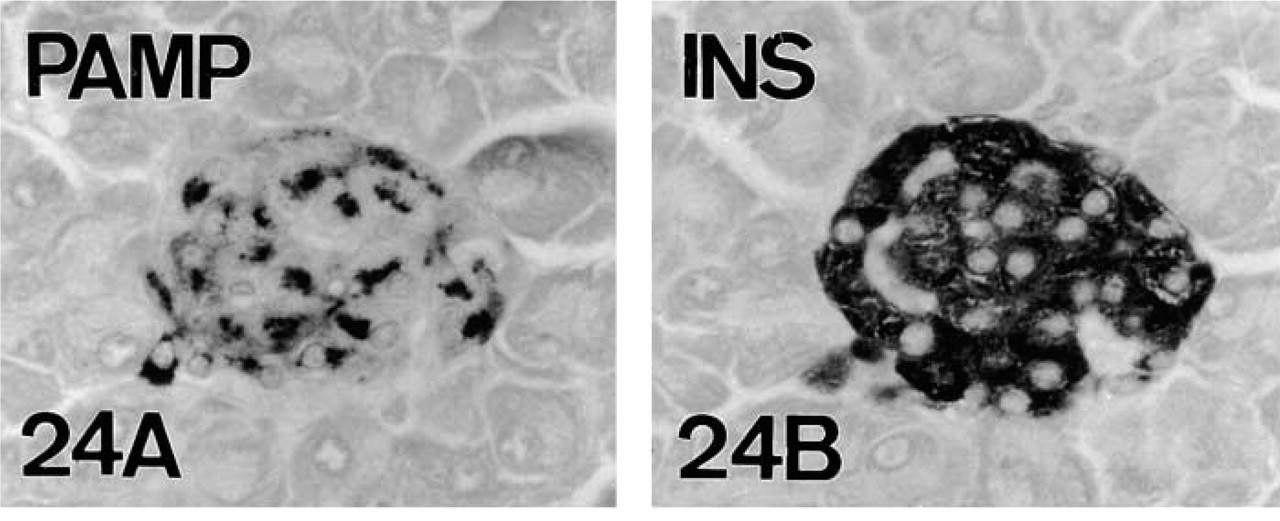
PAMP-IR (
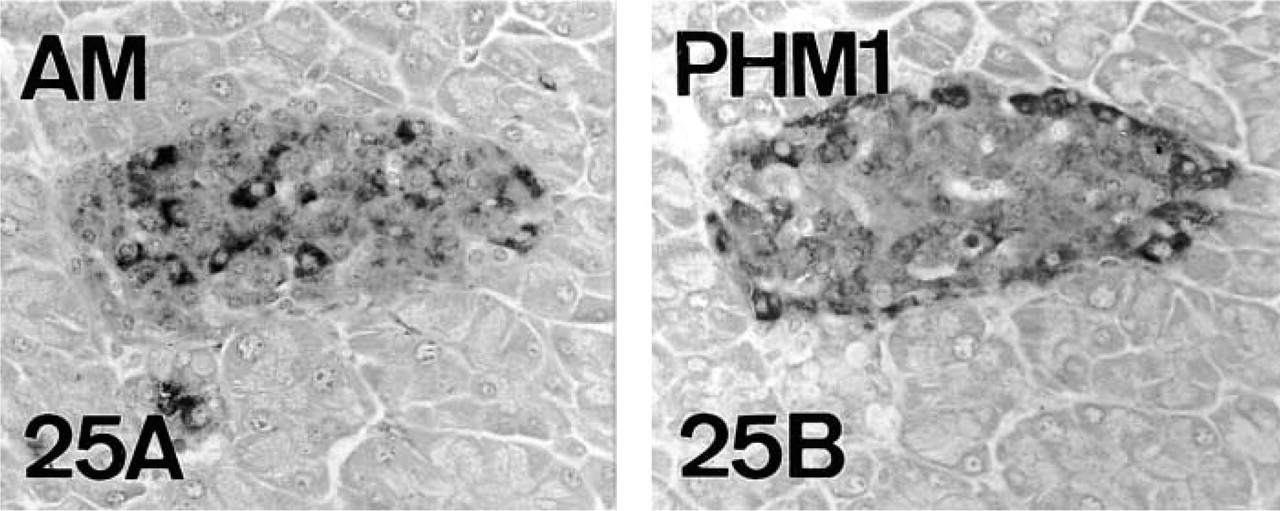
AM+ cells (
Both PHM antisera were expected to recognize the same cells by immunocytochemistry, given that the sequence against which the PHM3 antibody was raised (PAM 288–310) is enclosed in the sequence against which PHM1 was produced (PAM 37–382). However, PHM1 recognized B-cells, whereas PHM3 recognized A-cells (Table 4). This could be explained if PAM molecules present in A- and B-cells were somehow different, so that each form could only be recognized by one of the antisera but not by the other. Such molecular differences between mouse A- and B-cell's PAM molecules could be due either to different post-translational processing (e.g., proteolysis, glycosylation, phosphorylation), giving rise to conformational changes of the protein, or to alternative splicing, as previously reported in pancreatic carcinoma cells (Tateishi et al. 1994) and in other tissues (Stoffers et al. 1989, 1991; Zhang et al. 1997).
Furthermore, Western blots showed the presence of PAM-IR bands with different molecular weights depending on the antiserum used. These differences might also be explained by alternative splicing of the PAM gene product or by post-translational processing that would cause PAM proteins to run differently in the electrophoresis.
In relation to the cytoplasmic domain of PAM, immunolabeling was different in A- and D-cells (obtained with all fixatives) compared to B-cells (weaker and only in Zamboni-fixed tissue). Although differences in the amounts of enzymes containing the cytoplasmic domain may be considered, the reason for these differences is unknown. In addition, a different cellular localization of the IR for the cytoplasmic domain of PAM was obtained (restricted in A/D-cells and extended in B-cells, as discussed below).
PAM in A- and D-cells
In both cell types, PHM-IR was extended throughout the cytoplasm (presumably in the secretory granules), whereas the staining for the cytoplasmic domain of PAM was localized in a restricted juxtanuclear area. Such a morphological pattern suggests that the immunostained zones correspond to biosynthetic, presumably Golgi, areas. In view of the different immunolocalization patterns of PHM and the cytoplasmic domain regions, we suggest that proteolytic cleavage of cytoplasmic and transmembrane domains may occur in Golgi stacks, yielding soluble PAM enzymes in mouse A- and D-cells. A similar immunolabeling pattern in Golgi areas of PAM integral proteins (including the cytoplasmic domain of PAM) has been shown in neuroendocrine transfected AtT-20 cells (Milgram et al. 1993, 1997; Milgram and Mains 1994). These authors also suggested that proteolytic cleavage of integral domains takes place, yielding soluble PAM enzymes.
PAM in B-cells
Because in this case immunolabeling of both cytoplasmic and PHM domains was extended throughout the cytoplasm, differences in the biosynthetic pathway of PAM in mouse B- and A/D-cells appear to exist. Given that the cytoplasmic domain staining in B-cells is weaker than that of PHM, both soluble (PHM) and membrane-bound (PHM/cytoplasmic domain) PAM molecular forms could exist in the secretory granules. This would occur if proteolytic cleavage of cytoplasmic and transmembrane domains affected only part of the enzymatic pool. In fact, Western blotting showed the existence of a PHM1-IR band with a similar size (46 kD) to a cytoplasmic domain immunoreactive one, together with other PHM1-IR bands, presumably soluble, lacking a cytoplasmic domain.
Functional Significance of PAM in Mouse Endocrine Islets
Little is known about the significance of PAM in the endocrine pancreas. Insulin, glucagon and somatostatin are non-amidated peptides, and therefore the presence of the enzyme should be related to the presence of other peptides.
PAM in A-cells. According to our results, the production of GLP-1 and AM could account for the presence of amidating enzymes in mouse A-cells. We have found a total co-localization of GLP-1 with glucagon, as previously described in rat (Fridolf et al. 1991). Consequently, GLP-1 and PHM4 co-exist only in a subpopulation of A-cells. As indicated, they probably correspond not to static but to changing PAM-producing groups of cells.
PAM in B- and D-cells. In mouse B-cells, both AM and PAMP might be substrates for PAM enzymatic activity. Most B (insulin/PAM+)-cells are immunoreactive for AM and PAMP, although displaying different intensities for AM. Our results also show the presence of AM in D-cells.
The presence of AM has already been reported in pancreata of both mammalian (Washimine et al. 1995; Martinez et al. 1996), including a short reference in mouse (Cameron and Fleming 1998), and in non-mammalian (Lopez et al. 1999) species. Its presence in all vertebrate groups appears to indicate that AM plays an important role in the regulation of pancreatic function, including inhibition of insulin, response to septic shock, and amylase release (Martinez et al. 1996; Elsasser et al. 1999; Tsuchida et al. 1999).
Previous studies in rat pancreas reported strong AM-IR in PP cells and a low intensity of AM staining in the rest of the pancreatic endocrine cell types (Martinez et al. 1996, 1998). In the present study, although we also found different immunostaining intensities in the islets of Langerhans, A-, B-, and D-cells that stained strongly for AM and also for the amidating enzyme PAM were observed.
In summary, PAM-IR has been localized to adult mouse endocrine pancreatic A-, B-, and D-cells, colocalizing with amidated peptides such as GLP-1, AM, and PAMP. Subpopulations of PAM-producing cells have been found. Different immunostaining and Western blotting patterns suggest the existence of diverse PAM molecular forms in mouse pancreatic cells.
Footnotes
Acknowledgments
Supported by the PIUNA (University of Navarra).
We wish to thank Prof F. Cuttitta (National Cancer Institute; Rockville, MD) and Prof B.A. Eipper (Johns Hopkins University; Baltimore, MD) for the generous gift of some of the antisera and peptides used in this work.