
Abstract
The extracellular matrix proteolytic machinery is known to play a major role in trophoblast invasion, a process that shares similar features with the pathology of tumor invasion. In this study we investigated the expression of the recently described membrane-type matrix metalloproteinase-1 (MT-MMP-1; MMP-14) in early human placenta and decidual membrane to determine whether it might play a role in invasion. With in situ hybridization, the cytotrophoblasts of trophoblastic columns and the infiltrating intermediate trophoblasts in the decidual membrane were found to be the main producers of MT-MMP-1 mRNA. Gene expression was also seen in the villous double-layered trophoblastic epithelium and in the decidual cells of the decidual membrane. In endothelial and fibroblastic cells, however, the hybridization signal was either very weak or nonexistent. Immunohistochemical analysis and immunoelectron microscopy correlated well with the in situ hybridization findings. The most significant exception to this consisted of pericytes of spiral arteries, which appeared to lack MT-MMP-1 mRNA but showed intensive intracytoplasmic staining for the antigen. Our results show that MT-MMP-1 mRNA production is highly characteristic of intermediate trophoblasts, and MT-MMP-1 may have general importance in the tissue organization of early human placenta. We propose that MT-MMP-1 could be one of the key enzymes in the process of trophoblast invasion, acting alone or as a cell-surface activator of other proteinases.
B
Membrane-type matrix metalloproteinase-1 (MT-MMP-1) is a membrane-associated metalloproteinase discovered by PCT homology screening of placental cDNA library. It was isolated as a unique 3.4-kilobase (
In epithelial malignancies such as human colon, breast, head, and neck carcinomas, MT-MMP-1 mRNA synthesis by in situ hybridization was localized in stromal fibroblasts but not in cancer cells, which, however, showed the location of the antigen mostly intracytoplasmically and sometimes also on the carcinoma cell membrane (Okada et al. 1995). This is similar to the results described earlier for MMP-2 (Autio–Harmainen et al. 1992,1993; Pyke et al. 1993; Naylor et al. 1994).
Human placenta exhibits dramatic invasive properties in early pregnancy, when it is anchored to the uterine wall. Trophoblast invasion is precisely regulated to be confined spatially to the uterus and temporally to early pregnancy. Therefore, early human placenta can be regarded as a physiological counterpart to invasive malignant tumors. In this study we used in situ hybridization, immunohistochemistry, and immunoelectron microscopy to demonstrate that unorganized cytotrophoblasts of trophoblastic columns and invasive intermediate trophoblasts are the main producers of MT-MMP-1 mRNA and that villous trophoblastic epithelium is also capable of synthesizing MT-MMP-1 mRNA.
Materials and Methods
Materials
Five placentas were obtained from legal abortions at 8–10 weeks of gestation, performed for socioeconomic reasons. For in situ hybridization and immunohistochemistry, the placenta tissue was frozen in liquid isopentane. Altogether, 17 different samples from the five placenta were studied by in situ hybridization.
cRNA Probes and In Situ Hybridization
A 420-BP fragment extending from nt 218 to nt 638 of the human MT-MMP-1 cDNA sequence described originally by Sato et al. (1994) was amplified by polymerase chain reaction (PCR) from human placenta cDNA and ligated into BluescriptSK+ vector (Stratagene; La Jolla, CA) to generate clone pBL420. On Northern blot analysis of human tissues this probe recognized a single transcript of around 4.0
In situ hybridization was performed as previously described by Autio–Harmainen et al. (1992) and corresponding sense probes were used as controls to determine specific hybridization. All the solutions were treated with 0.1% diethylpyrocarbonate (DEPC) (Fluka; Buchs, Switzerland).
For in situ hybridization, 8-μm-thick cyrosections were placed on SuperFrost Plus (Menzel–Gläser, Braunschweig, Germany) slides and fixed with 4% paraformaldehyde. After dehydration the sections were incubated in 0.2 M HCl (20 min) at room temperature (RT), followed by a 5-min wash DEPC-H2O). After treatments with proteinase K (Boehringer–Mannheim; Mannheim, Germany) (1 μg/ml, 30 min, 37C) and 0.2% glycine in PBS and washings twice in 1 × PBS for 30 sec, the slides were immersed in 4% paraformaldehyde in PBS for 20 min. The sections were then washed in 1 × PBS and acetylated twice in 0.25%-0.5% acetic anhydride in 0.1 M triethanolamine, twice for 5 min. After a 5-min wash in PBS and dehydration, the sections were allowed to air-dry for 1–2 hr at RT before prehybridization with a hybridization mixture not including the labeled probe for about 2 hr at 50C. The hybridization mixture contained the labeled probe, 10 mM dithiotreitol (DTT), yeast tRNA (1 mg/ml) deionized formamide (50% of the volume), dextran sulfate (10% of the volume), 1 × SALTS [10 × SALTS; 3 M NaCl, 0.1 M Tris-HCl, 0.1 M NaPO4, 0.05 M EDTA, 0.02% Ficoll (w/v), 0.02% polyvinylpyrrolidone (PVP) (w/ v), and bovine serum albumin (BSA) (0.2 mg/ml)]. The labeled riboprobes were diluted at a concentration of 3 × 106 cpm per 40-ml aliquot per slide. Before placing on sections, the labeled probe was denatured by boiling for 1 min and placed on ice. The hybridization was carried out at 50C overnight. The slides were then processed through the following stringency washes containing 50% formamide, 1 × SALTS, 10 mM DTT twice for 1 hr at 50C, followed by a wash in 0.5 M NaCl in 10 mM Tris-HCl, 1 mM EDTA (TE), pH 7.5, at 37C for 15 min, and treatment with RNase A (Boehringer–Mannheim) (40 mg/ml) in 0.5M NaCl in TE buffer for 30 min at 37C. After that, the sections were washed in 0.5 M NaCl in TE for 15 min at 37C and in 2 × SSC twice for 15 min at 45C and 1 × SSC twice for 15 min at 45C. After dehydration through alcohol series, the slides were air-dried and dipped in Kodak NTB-2 (Eastman Kodak; Rochester, NY) photographic emulsion diluted 1:1 with 1% glycerol in water. After exposure for 10–14 days, the slides were developed in Kodak D19 (Eastman Kodak) developer and counterstained with hematoxylin and eosin.
Immunohistochemistry
For immunohistochemical staining, 4-μm cryosections from two placentas and paraffin sections from one placenta were stained using avidin and biotinylated horseradish peroxidase complex (Dako; Dakopatts, Glostrup, Denmark). Nonspecific binding was blocked with fetal calf serum (FCS) 1:5 in PBS. Sections were incubated with monoclonal antibody against MT-MMP-1 (Cao et al. 1995) (Fuji Chemical Industries; Takaoka, Japan) (1 mg/ml in PBS+NaN3; diluted 1:25 and 1:50) overnight at 4C and after that were incubated with biotinylated anti-mouse IgG secondary antibody (Dako) for 30 min, avidin–peroxidase complex for 30 min, and the substrate solution (3,3-diaminobenzidine tetrahydrochloride in H2O2 in Tris buffer), pH 7.4 (Sigma; Poole, UK) for 10 min. For control stainings, PBS was used instead of the primary antibody. Intermediate trophoblasts (Kurman et al. 1984) were identified by using anti-cytokeratin Mak 6 (Triton Diagnostics; Alameda, CA) and human placental lactogen (hPL) (Dako).
Immunoelectron Microscopy
Samples from the placental tissue were fixed with 8% para-formaldehyde in 0.1 M phosphate buffer, pH 7.4, for 2 hr. They were cryoprotected by immersion in 2.3 M sucrose for 30 min and then frozen in liquid nitrogen.
Semithin cryosections were stained with toluidine blue and cytokeratin immunohistochemistry for light microscopic evaluation.
The sections were first incubated in 10% FCS with 0.02 M glycine in PBS (PBS–glycine) on ice for 10 min. They were then incubated with the monoclonal antibody to MT-MMP-1 diluted optimally with 5% FCS in PBS–glycine for 45 min and washed in PBS–glycine (four changes in 15 min). After that, the sections were exposed with the antibody to rabbit anti-mouse IgG (Dako) diluted 1:200 with 5% FCS for 30 min and washed in PBS–glycine (six changes in 15 min) before exposure to the protein A–gold complex diluted 1:20 in 5% FCS in PBS–glycine for 20 min. The sections were washed in PBS–glycine (six changes in 15 min) and postfixed in 2.5% glutaraldehyde for 10 min, followed by brief washings in PBS–glycine and distilled water (five changes in 10 min). Finally, the sections were embedded in 2% methylcellulose with 0.3% uranyl acetate on ice for 10 min, dried, and examined in a Philips 410 LS transmission electron microscope by using an acceleration voltage of 60 kV. In the control sections, the antibody was omitted and replaced with PBS.
Results
In Situ Hybridization and Immunohistochemical Findings
Cytotrophoblastic Columns
In situ hybridization revealed a very strong signal for MT-MMP-1 mRNA in the unorganized cytotrophoblastic cells of trophoblastic columns. All the cells of the columns showed similar hybridization signal except for the multinucleated syncytial giant cells, in which there was considerably less label, often at the background level (Figures 1A and 1B). Immunohistochemically, the mononuclear cytotrophoblasts of columns were all stained for the antigen. The staining reaction was mostly located on the cell membrane, but some cytotrophoblasts also showed intensive intracytoplasmic staining (Figure 1C). Multinucleated giant cells in the columns were negative, except for positive staining in the microvillous projections of the cell membrane (Figure 1C, inset).
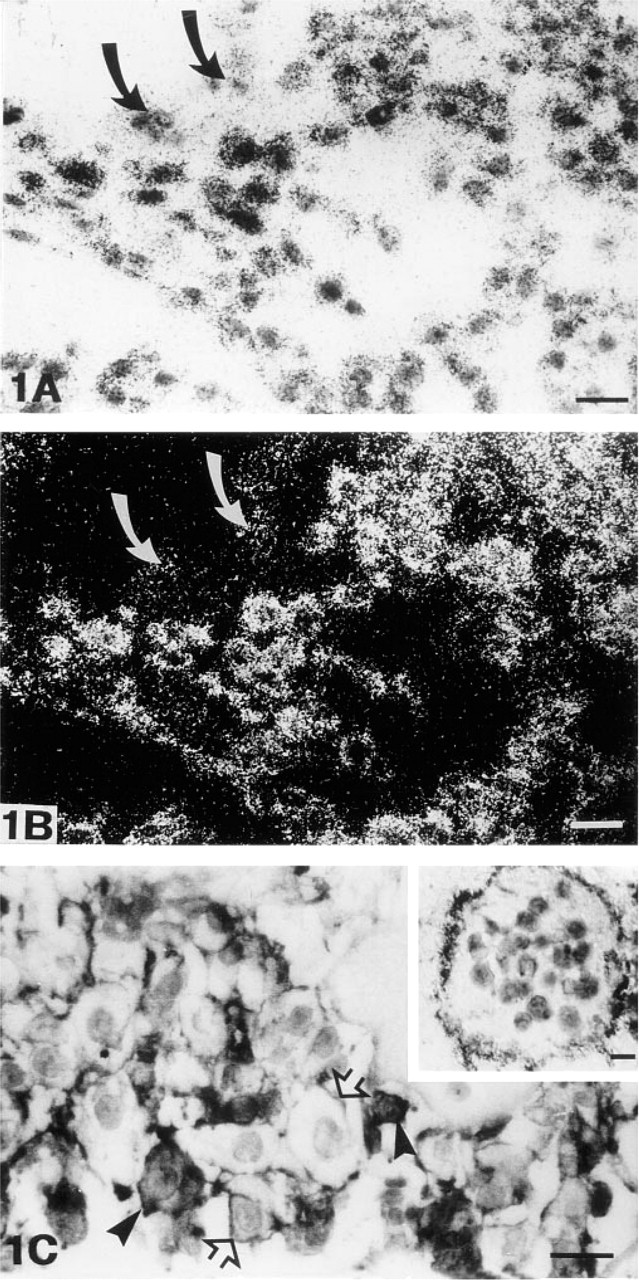
(
Intermediate Trophoblasts
The intermediate trophoblasts of the trophoblastic shell and those infiltrating into the superficial decidual membrane were identified on the basis of positivity for cytokeratin (Figures 2A and 2B) and for human placental lactogen (Figure 2C). Intermediate trophoblasts very intensively expressed MT-MMP-1 mRNA (Figure 3A and 3B). These cells were especially numerous in the decidual membrane around spiral arteries. In such areas, the spiral artery wall and the adjacent decidual tissue often contained small fibrinoid necroses (Figure 3C). Intermediate trophoblasts exhibited an intensive, mostly intracytoplasmic staining reaction in immunohistochemical analysis (Figure 3D).
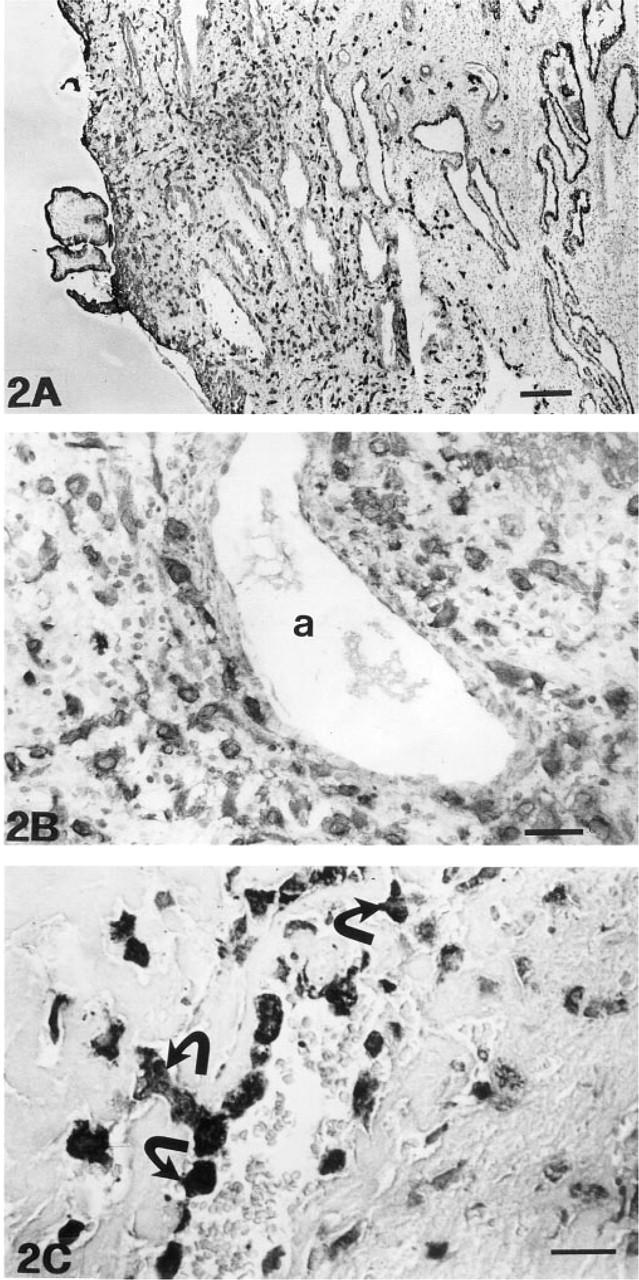
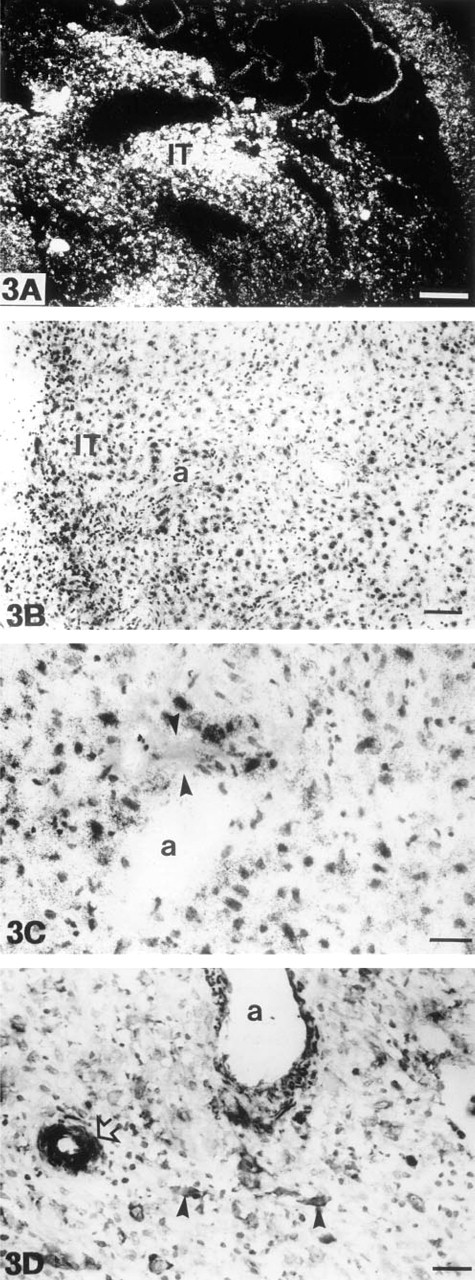
Placental Villi
A clear, although much fainter MT-MMP-1 mRNA expression than in cytotrophoblasts and intermediate trophoblasts was present in the cells of double-layered trophoblastic epithelium of villi (Figures 4A and 4B). Both the syncytial and the cytotrophoblastic cell layers expressed MT-MMP-1 mRNA, but the signal in the syncytial layer was often more pronounced. Expression in villous endothelial and fibroblastic cells was either very week or nonexistent.
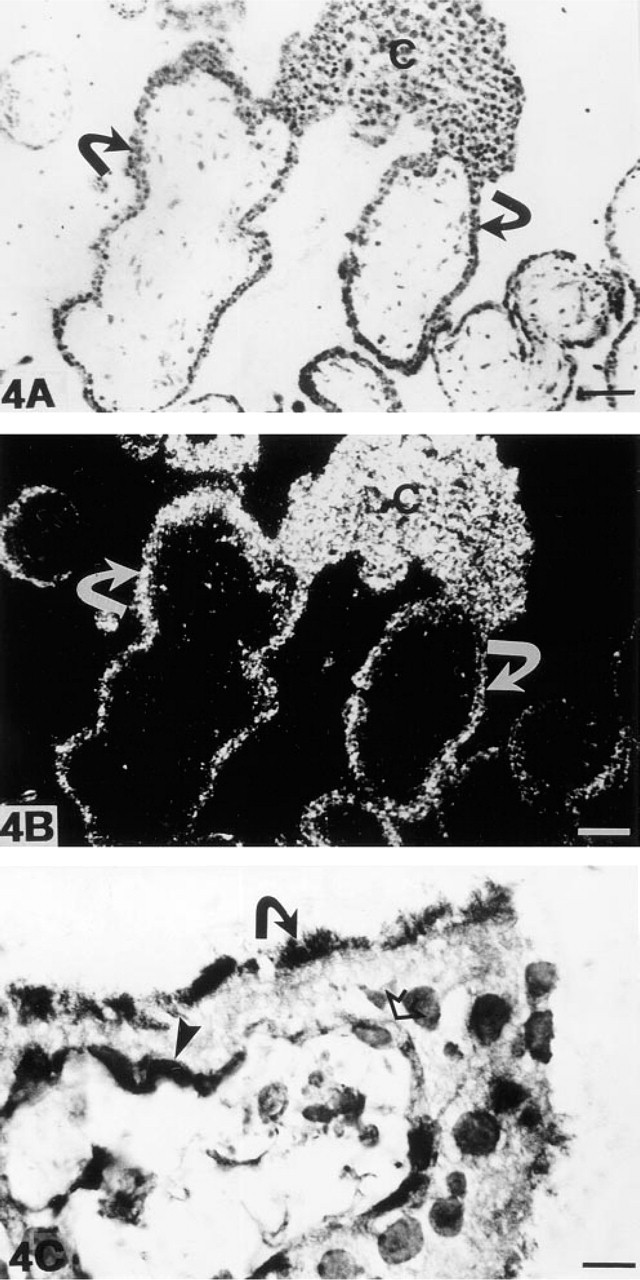
In immunohistochemical analysis, the cells of the cytotrophoblastic layer displayed strong labeling, which was located either intracytoplasmically or on the cell membrane. The syncytial cell layer showed a staining reaction mainly on the apical border, at which microvillous projections were stained (Figure 4C). The endothelial cells of villous capillaries and stromal fibroblasts showed a faint intracytoplasmic staining reaction.
Decidual Membrane
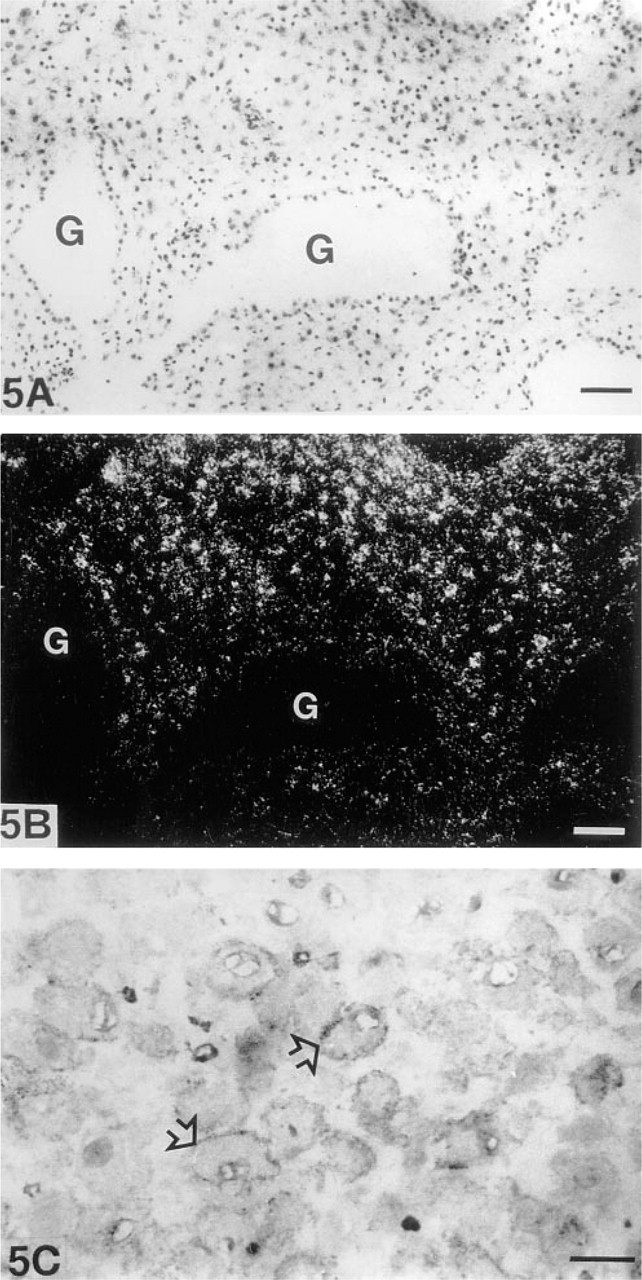
The large decidual cells of the decidual membrane clearly expressed MT-MMP-1 mRNA (Figures 5A and 5B). They showed granular positive depositions concentrating close to the cell membrane in immunohistochemical analysis (Figure 5C). The epithelial cells of endometrial glands were negative for both MT-MMP-1 mRNA and the antigen. The endothelial cells and pericytes of spiral arteries showed either very weak or nonexistent MT-MMP-1 mRNA expression, but fibrocytic endometrial stromal cells apposing spiral arteries exhibited a strong signal for the MT-MMP-1 mRNA (Figure 3B). An intriguing immunohistochemical finding was the intensive intracytoplasmic staining reaction of the pericytes of spiral arteries (Figure 3D).
Immunoelectron Microscopic Findings
Immunoelectron microscopic results generally correlated with the immunohistochemistry, showing immunolabeling in the cytoplasm, in microvillous projections or close to the cell membrane in syncytial and cytotrophoblastic cells of villi, including multinucleated giant cells, and in large decidual cells of decidual membrane (Figures 6A–6E). Similar accentuation of labeling towards the cell membrane, as was evident by immunohistochemistry, cold not be detected by immunoelectron microscopy. Clear intracytoplasmic labeling was also seen in the endothelial and fibroblastic stromal cells of villi.
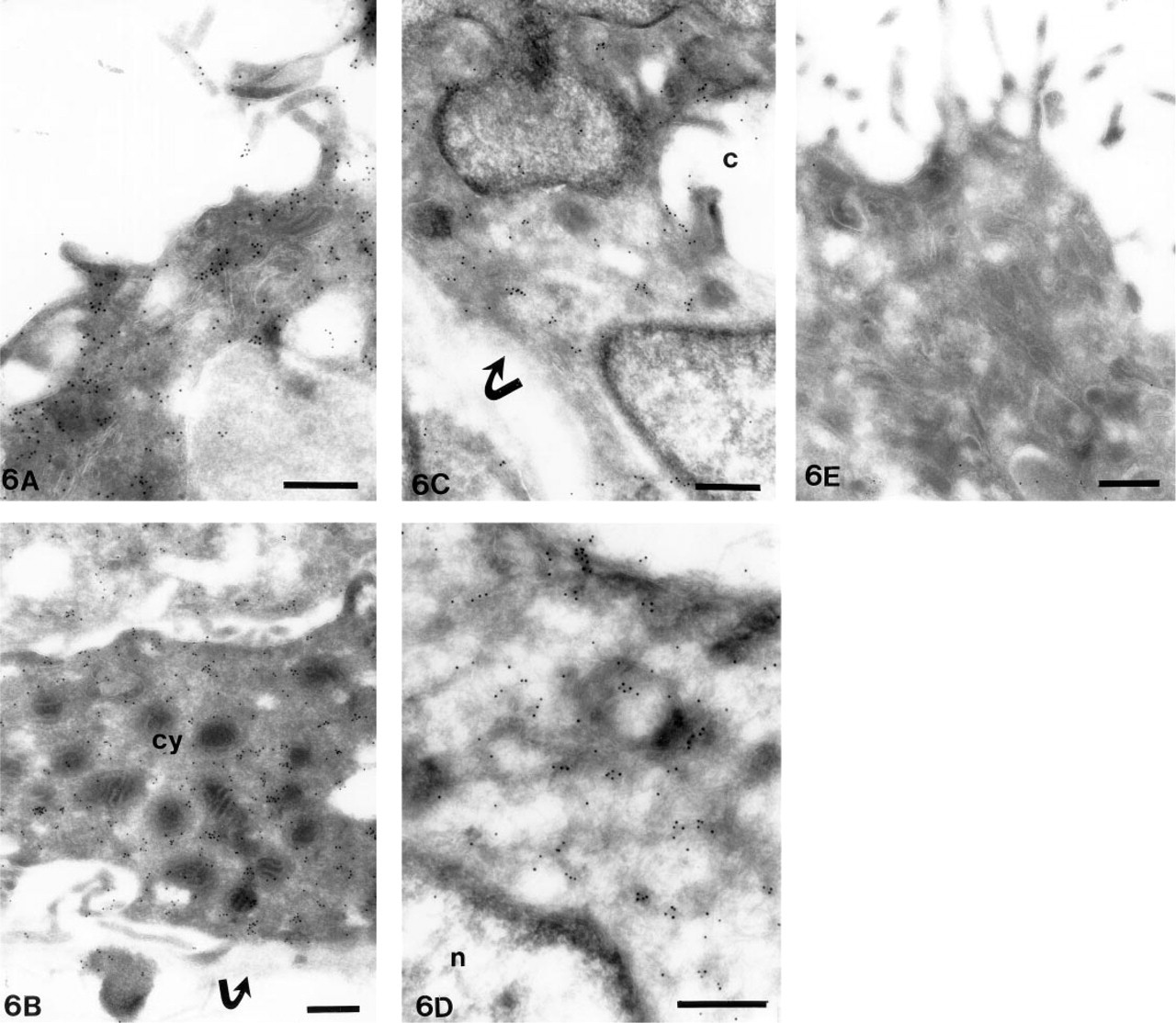
In immunoelectron microscopy, labeling for MT-MMP-1 is seen in the microvillous projections and in cytoplasm of syncytial (
Discussion
Evidence obtained by Northern analysis indicated MT-MMP-1 mRNA expression by cells of human placental tissue. However, there is no previous knowledge about which cells are responsible for this production. Here we describe the MT-MMP-1 mRNA expression in cells of 8–10-gestational week human placentas and correlate the findings with the previously described expression patterns of other ECM proteins involved in matrix turnover in developing placenta. These include laminin-1, Types IV and XIII collagen, and MMP-2 and MMP-9 (Autio–Harmainen et al. 1991, 1992; Juvonen et al. 1993; Hurskainen et al. 1996).
In the developing placenta, the unorganized cytotrophoblastic cells of trophoblastic columns form the cell reservoir for new villi and anchor the ovum to the uterine wall. The anchoring process involves invasion of cells to the decidual membrane, endometrium, and spiral arteries in early pregnancy. It was previously shown that cytotrophoblasts of columns were potent producers of various ECM proteins, such as MMP-2 and MMP-9, TIMP-1, −2, and −3, laminin, and Types IV and XIII collagen (Autio–Harmainen et al. 1992; Juvonen et al. 1993; Polette et al. 1994; Hurskainen et al. 1996), and we demonstrate here that these cells also produce significant amounts of MT-MMP-1 mRNA.
Studies by Alexander and Werb (1992) and Librach et al. (1991) have emphasized the importance of MMPs, more specifically MMP-9, and TIMPs for trophoblast invasion. Invasive trophoblasts, termed intermediate trophoblasts by Kurman et al. (1984), form a distinct cell population among trophoblasts, sharing morphological features of Langerhans-type cytotrophoblasts and decidual cells. They form the trophoblastic shell on the surface of the decidual membrane and they also possess invasive capacity. According to our results, production of MT-MMP-1 is highly characteristic of these cells and may indicate a crucial role for MT-MMP-1 in trophoblast invasion, because a corresponding accentuation in the in situ hybridization studies for MMP-2 and MMP-9 was unable to be detected (Autio–Harmainen et al. 1992; Hurskainen et al. 1996). Our results therefore suggest that MT-MMP-1 may be one of the key enzymes among MMPs that regulates the controlled invasion of trophoblasts to the uterine wall.
MT-MMPs form a unique group of proteinases in the MMP superfamily (Sato et al. 1994; Takino et al. 1995; Will and Hinzmann 1995; Puente et al. 1996). Since 1985, several studies have been published demonstrating the presence of gelatinase-type proteinase bound to the cell membrane Zucker et al. (1985) described enriched collagenolytic and gelatinolytic activity in pancreatic adenocarcinoma plasma membranes. In additional studies, they demonstrated, using gelatin zymography, that well-characterized pancreatic cancer plasma membrane comprised a distinct molecular weight (92 kD, 70 kD, and 62-kD) metalloproteinase species (Zucker et al. 1990). Later, specific cell-surface binding sites for MMP-2 were described on breast cancer cells (Emonard et al. 1992), although no membrane protein that functions as a metallopteteinase receptor has yet been identified. Finally, Sato and coworkers (1994) described MT-MMP-1. Our findings suggest that MT-MMP-1 in the cells of placenta, as described in this study, may function as an important regulator for the activity of MMPs, particularly MMP-2.
We have previously shown that MMP-2 is localized either intracytoplasmically or on the cell membranes of cytotrophoblasts and ovarian carcinoma cells (Autio–Harmainen et al. 1992, 1993). Interestingly, villous trophoblastic epithelial cells were shown to lack MMP-2 mRNA but were intensely stained for the antigen. Such a discrepant finding was explained by the possible presence of a receptor-like mechanism on the epithelial cell surface to bind the immunohistochemically detectable MMP-2 (Autio–Harmainen et al. 1992). We found here a clear signal for MT-MMP-1 in syncytial and a fainter signal in cytotrophoblastic villous epithelium by in situ hybridization. Immunohistochemical stainings and immunoelectron microscopy revealed that the MT-MMP-1 protein was localized on cell membranes of cytotrophoblastic epithelial cells and in the microvillous projections of syncytial epithelial cells. In view of the demonstrated ability of MT-MMP-1 to activate MMP-2 on the cell surface (Sato et al. 1994), the previous explanation of the presence of the receptor-like mechanism on villous trophophoblasts appears to be valid.
Nomura et al. (1995) demonstrated immunohistochemically the expression of MT-MMP-1 in human gastric carcinoma tissue, in which MT-MMP-1 was co-localized in the carcinoma cells in almost all MMP-2-positive cases. They also showed that fibroblastic cells and vascular endothelial cells stain positively for MT-MMP-1. Okada et al. (1995) localized MT-MMP-1 transcripts in fibroblastic cells of tumor stroma but not in cancer cells which, however, they found to be positive in immunohistochemical staining for MT-MMP-1. In the present study, we found the most abundant signal in the trophoblastic cells, followed by a clearly lower signal in the endothelial cells and the lowest or often nonexistent signal in fibroblasts. A common finding with respect to carcinomas appears to be that MT-MMP-1 mRNA is abundant solely in fibroblastic cells, although immunohistochemistry detects an additional presence in endothelial and cancer cells. This is quite comparable with the findings described for MMP-2 in carcinomas (Autio–Harmainen et al. 1993; Pyke et al. 1993; Höyhtyä et al. 1994). Our present study further shows that in developing embryonal tissue, such as placenta, there is a wider range of cells, such as trophoblasts and endothelial cells in addition to fibroblasts, that are capable of synthesizing MT-MMP-1 mRNA.
The presence of a complex proteolytic enzyme profile in first-trimester trophoblastic cells is consistent with the fact that first-trimester cells are capable of penetrating a variety of maternal cell layers and the associated BMs to gain access to spiral arteries. The large decidual cells of the decidual membrane contained a relatively week although clear signal for MT-MMP-1 mRNA. Because invasive intermediate trophoblasts were shown to express considerably higher levels of MT-MMP-1 mRNA than decidual cells, it can be concluded that MT-MMP-1 needed for invasion is produced mainly by the invasive cells themselves. A novel finding was also the intense intracytoplasmic staining of pericytes, although the mRNA signal in these cells was either very low or completely absent. It is possible that the MT-MMP-1 protein is synthesized elsewhere, either in decidual cells and/or stromal cells around spiral arteries, and then is selectively bound or taken up by pericytic cells.
Our results, taken together with the published biochemical studies on MT-MMP-1, suggest an important role for MT-MMP-1 in tissue organization of early placenta and in the invasion of cytotrophoblastic cells to decidua and spiral arteries. Because it is abundantly produced by invasive intermediate trophoblasts, MT-MMP-1 is proposed to be one of the key enzymes among MMPs in the process of trophoblast invasion during early pregnancy.
Footnotes
Acknowledgements
Supported by the Cancer Society of Finland and the Cancer Society of Northern Finland.
We are grateful to Prof Karl Tryggvason for valuable comments during the preparation of this manuscript and to Dr Kazushi Iwata (Fuji Chemical Industries, Ltd.; Takaoka, Japan) for preparing the monoclonal antibodies. We also thank Ms Annikki Huhtela and Ms Riitta Karvonen for expert technical assistance and Mr Tapio Leinonen for preparing the micrographs.