
Abstract
We investigated the expression of the immunoglobulin superfamily cell adhesion molecule, C-CAM, in developing and mature rat placenta. By immunohistochemical staining at the light microscopic level, no C-CAM-expression was seen before Day 9 of gestation, when it appeared in the trophoblasts of ectoplacental cones. On Day 10.5, spongiotrophoblasts and invasive trophoblasts around the maternal vessels of the decidua basalis were stained positively. On Day 12.5, C-CAM was detected in the spongiotrophoblasts of the junctional layer, but labyrinth trophoblasts and secondary giant trophoblasts were not stained. On Day 17.5, C-CAM was found only in the labyrinth and lacunae of the junctional layer. At this stage, both the labyrinth cytotrophoblasts of the maternal blood vessels and the endothelial cells of the embryonic capillaries were strongly stained. Placental tissues from gestational Days 12.5 and 17.5 were analyzed by immunoelectron microscopy to determine the location of C-CAM at the subcellular level. On Day 12.5, positive staining of the spongiotrophoblasts was observed, mainly on surface membranes and microvilli between loosely associated cells. On Day 17.5, staining was found primarily on the microvilli of the maternal luminal surfaces of the labyrinth cytotrophoblasts, and both on the luminal surface and in the cytoplasm of endothelial cells of the embryonic vessels. RT-PCR analysis and Southern blotting of the PCR products revealed expression of mRNA species for both of the major isoforms, C-CAM1 and C-CAM2. Immunoblotting analysis of C-CAM isolated from 12.5-day and 14.5-day placentae showed that it appeared as a broad band with an apparent molecular mass of 110–170 kD. In summary, C-CAM was strongly expressed in a specific spatiotemporal pattern in trophoblasts actively involved in formation of the placental tissue, suggesting an important role in placental development. In the mature placenta, C-CAM expression was confined to the trophoblastic and endothelial cells lining the maternal and embryonic vessels, respectively, suggesting important functions in placental physiology.
IN MANY MAMMALS, implantation involves invasion of the endometrium by trophoblasts to establish the placenta and set up communication with the maternal blood circulation (Welsh and Enders 1987, 1991). Extensive communication among trophoblasts and the endometrium, decidual cells, and maternal vascular cells plays an important role in the organization of placental development (Strickland and Richards 1992; Aplin 1991). This involves intercellular and cell–matrix interactions that are mediated by several classes of cell adhesion molecules (Ruck et al. 1994; Leach et al. 1993; Lessey et al. 1992; Damjanov 1986; Richa et al. 1985) and extracellular matrix proteins (Damsky et al. 1992; Castelluci et al. 1991; Aplin et al. 1988). The majority of the cell adhesion molecules can be grouped into three large protein superfamilies, the immunoglobulin superfamily (IgSF), the cadherins, and the integrins, and one smaller family, the selectins (Öbrink 1991). Whereas the former three are involved in a number of cellular interactions in solid tissues, the selectins function primarily in leukocyte trafficking.
C-CAM is a well-characterized cell adhesion molecule of the IgS family (Öbrink 1991), which is expressed in various epithelial and hematopoietic cells and in the endothelia of small blood vessels (Murphy et al. 1994; Sawa et al. 1994; Odin et al. 1988; Odin and Öbrink 1987). It also appears to have important roles in development, e.g., in the late phases of liver maturation (Odin and Öbrink 1986, 1988) and tooth formation (Lüning et al. 1995; and Terling et al., submitted for publication). Daniels et al. (1996) recently suggested that biliary glycoprotein 1 (Bgp-1), a mouse homologue of rat C-CAM, may be involved in a number of specific embryonic processes. Svalander et al. (1987) demonstrated that C-CAM appears in the trophectoderm of normal 4-day and delayed rat blastocysts but disappears from the embryonic cells at the time of implantation. After implantation, no expression of C-CAM could be found in the embryonic tissues up to 8 days of gestation (Svalander et al. 1990). It was suggested that C-CAM might be involved in the regulation of blastocyst implantation.
C-CAM is a transmembrane, highly glycosylated protein that occurs in several isoforms owing to differential splicing (Edlund et al. 1993; Najjar et al. 1993; Culic et al. 1992; Öbrink 1991). The two major splice variants, C-CAM1 and C-CAM2, comprise four Ig-like domains in their extracellular portions but differ in their cytoplasmic domains (Edlund et al. 1993). The cytoplasmic domain of C-CAM1 consists of 71 amino acids, whereas that of C-CAM2 contains only 10 amino acids. In addition, two allelic variants of C-CAM have been found in rats and mice (Edlund et al. 1993; McCuaig et al. 1993), whereas a similar allelic variation has not yet been reported in humans. The allelic variants, which have been named a and b in the rat (Edlund et al. 1993), differ in 16 amino acids in the most N-terminal of the Ig domains, but the remaining parts of the molecules are identical.
It has recently been found that C-CAM1 isoforms in both rat and mouse appear to be negative regulators of cellular growth and function as tumor suppressors in both prostate and colon carcinogenesis (Hsieh et al. 1995; Kunath et al. 1995). The cytoplasmic domain of C-CAM1 contains phosphorylatable tyrosine signaling motifs, which on phosphorylation can bind tyrosine kinases, e.g., pp60c-src (Brümmer et al. 1995). The functional role of the shorter cytoplasmic domain of the C-CAM2 isoforms is not yet known, but it has been found that it can be phosphorylated on a specific serine residue by protein kinase C (Edlund et al. 1996b), and the calcium-regulated protein calmodulin can bind to this domain as well as to the cytoplasmic domain of C-CAM1 (Edlund et al. 1996a; Edlund and Öbrink 1993).
In addition to cell adhesion and transmembrane signaling it appears that C-CAM can influence membrane transport. Therefore, C-CAM1 has been found to affect transport of bile acids through the plasma membrane (Sippel et al. 1993, 1994).
To learn more about the functions of C-CAM, we decided to determine when and where it appears during postimplantation embryonic development. We now report that C-CAM becomes expressed in proliferating and invading trophoblasts in the developing placenta from Day 9 of gestation. In the mature placenta it is expressed in cytotrophoblasts of the maternal blood lacunae and in embryonic capillary endothelial cells.
Materials and Methods
Animals
Sprague–Dawley rats were obtained from the local breeding colony at the animal facility of Kyorin University, School of Medicine. The day on which a vaginal plug became apparent was regarded as Day 0.5 of gestation (E0.5). Rat placentae of gestational days 6.5 (E6.5), 7.5 (E7.5), 8.5 (E8.5), 10.5 (E10.5), 12.5 (E12.5), 14.5 (E14.5), 15.5 (E15.5), 17.5 (E17.5), and 19.5 (E19.5) were studied. The animals were sacrificed by ether anesthesia and the placentae were removed and fixed with 4% paraformaldehyde in 0.1 M phosphate buffer, pH 7.4 (PB) for light and electron microscopic immunohistochemical analysis. For immunoblotting and RNA analyses, the tissues were immediately frozen in liquid nitrogen and stored at −80C until used.
Antibodies
Rabbit polyclonal antibodies against purified rat liver C-CAM were produced and characterized as previously described (Odin et al. 1987). They were shown to be monospecific for C-CAM by immunoblotting of liver. Rabbit antibodies against human pregnancy-specific glycoproteins (PSG) were obtained from Dako (Glostrup, Denmark). They have been demonstrated to crossreact with PSGs of rat origin (Ogilvie et al. 1989).
Light Microscopic Immunohistochemistry
Rat placental tissues were fixed with 4% paraformaldehyde in 0.1 M PB and embedded in paraffin. Sections (5 μm) were cut, deparaffinized, and blocked with 0.3% H2O2 in methanol to quench endogenous peroxidase activity. Immunostaining was performed using the avidin–biotin–peroxidase complex (ABC) method (Hsu et al. 1981). The sections were incubated with 10% normal goat serum for 30 min and then with anti-C-CAM antiserum [diluted 1:1000 with Tris-buffered saline (TBS); 150 mM NaCl, 50 mM Tris HCl, pH 7.6], or anti-PSG antiserum (diluted 1:200 with TBS) for 2 hr. Nonimmune rabbit serum was used as a control. The sections were washed three times with TBS, incubated with biotinylated anti-rabbit IgG (Dako) (diluted 1:500 in TBS) for 1 hr, washed with TBS, and incubated with avidin-biotin–peroxidase complex (Dako) for 1 hr. All incubations were performed at room temperature (RT). The peroxidase was visualized using 3, 3′-diaminobenzidine tetrahydrochloride (Wako; Tokyo, Japan) as a substrate in the presence of 0.03% H2O2, and the sections were counterstained with 1% hematoxylin.
Electron Microscopic Immunohistochemistry
The tissues were fixed with 4% paraformaldehyde in PB, immersed in a graded series of sucrose in 0.1 M PB up to 30% sucrose, embedded in OCT compound (Tissue Tek; Miles, Elkhart, IN), and frozen in liquid nitrogen. Sections (20 μm thick) were cut on a Histostat microtome (Meiwa; Tokyo, Japan) and mounted on plastic plates. The plates were immunostained as described for light microscopic immunohistochemistry. They were then washed with PB, dehydrated in a graded series of ethanols, immersed in propylene oxide, and embedded in epoxy resin. Sections (1 μm thick) were cut and stained with toluidine blue for light microscopic examination. Selected areas were then thin-sectioned, stained with uranyl acetate and lead nitrate, and examined with a Hitachi H-7000 electron microscope at 75 kV.
Immunoblotting
Rat placentae were stored at −80C until used. To examine whether placental C-CAM is an integral membrane protein, the samples were analyzed by phase separation in Triton X-114. Placental tissues were homogenized in 2% Triton X-114 (Sigma; St Louis, MO), 1 mM phenylmethylsulfonyl fluoride (Sigma) in TBS, in a glass homogenizer. The homogenate was placed on ice for 10 min and then centrifuged at 3000 × g for 5 min. The supernatant was layered on a 0.5-ml sucrose cushion (6% sucrose, 0.06% Triton X-114 in TBS) and the tubes were incubated for 10 min at 30C. The samples were then centrifuged at 18, 000 × g for 5 min at RT, which resulted in separation into a clear upper aqueous phase and an oily lower detergent phase. Each of the two phases was subjected to a second phase separation by centrifugation as described by Bodier (1981). The phases were mixed with equal volumes of SDS-PAGE sample buffer (4% SDS/0.05 M Tris-HCl, pH 6.8/0.01% bromophenol blue/ 10% glycerol), dithiothreitol (0.1 M) was added, and the mixtures were boiled for 10 min. The samples were then centrifuged at 18, 000 × g for 20 min, subjected to SDS-PAGE (Laemmli 1970), and transferred to nitrocellulose filters (Towbin et al. 1979). The filters were blocked with 1% fat-free powdered milk in TBS for 30 min at RT and were then incubated overnight at RT with anti-C-CAM antiserum diluted 1:200 in TBS. Nonimmune rabbit serum, instead of the primary antibody, was used as a control. The filters were then washed three times with 0.05% Tween 20/TBS and incubated with alkaline phosphatase-conjugated anti-rabbit IgG (Dako) diluted 1:2000 in TBS for 1 hr at RT. After washing in 0.05% Tween 20/TBS, the filters were developed with nitroblue tetrazolium (NBT) (Wako) and 5-bromo-4-chloro-3-indolyl phosphate (BCIP) (Sigma) in 5 mM MgCl2, 100 mM Tris-HCl, pH 9.0. Prestained proteins (Bio-Rad; Richmond, CA) were used as molecular weight markers.
RT-PCR and Southern Blot Hybridization
Total RNA was prepared from rat placentae using Isogen solution according to the manufacturer's protocol (Wako). In brief, placental tissues were disrupted in Isogen solution and an equal volume of chloroform was added. After centrifugation for 5 min at 18, 000 × g, the aqueous phase was precipitated with isopropanol. PolyA(+) RNA was purified using oligo-dT-latex (Oligo-dT 30 super; Takara, Japan) beads. Using 1 mg of polyA(+) RNA as template, cDNA was then produced with reverse transcriptase (Promega; Madison, WI) and random priming. The resultant cDNA was amplified by PCR with 40 cycles at 95C for 45 sec, 55C for 45 sec, and 72C for 45 sec, following the manufacturer's protocols of the GeneAmp RNA system (Perkin Elmer; Norwalk, CT). The primer sets used for amplification of C-CAM consisted of two forward primers: F5, Xho-1 (5′-GCC TCG AGA TGG AGC TAG CCT CGG CTC GT- 3′; nt. 1–21) and F3 (5′-GCA AGC TTT TTT GAG CCA GTG ACT CAG CCC TCC-3′; nt. 955–978), and a reverse primer B-6,Xho-1 (5′-GCC TCG AGC AGG ACA GAC AAT GTC AC-3′; nt. 1557–1574). The indicated nucleotide (nt) numbers refer to the C-CAM cDNA nucleotide sequence (Edlund et al. 1993). Rat G3PDH primers were purchased from Clontech (Richmond, CA). Each PCR-amplified product was electrophoresed on a 1.2% agarose gel containing 0.2% ethidium bromide. The PCR products that were amplified with the primer combination F5,Xho-1/B6,Xho-1 were purified by agarose electrophoresis and re-amplified with the primer combination F3/B6,Xho-1. To confirm a specific amplification of C-CAM cDNA, the PCR products were electrophoresed on 1.5% agarose gels, transferred to nylon membranes (Amersham; Poole, UK) using the alkali transfer method (0.4 M NaOH), and hybridized with C-CAM sequence-specific oligonucleotides. The following probes were used: Probe F (5′-TGT GGC TGG AGG TTG AGG GTT TGT GCT CTG TGA GAT CTC G-3′) covering nucleotides 1375–1414; Probe G (5′-GTC GTC AGA AGG ACC CAG ATC CGC CAG TCT TCC TG-3′) covering nucleotides 1347–1361 plus 1415–1434. Probe F recognizes sequences that are contained only in the large mRNA transcripts specific for C-CAM1, whereas probe G recognizes mRNA transcripts specific for both C-CAM1 and C-CAM2 variants under low-stringency conditions (Edlund et al. 1996a). Probes F and G were labeled with 11-digoxigenin (DIG)-dUTP, using the DIG oligonucleotide tailing kit of Boehringer–Mannheim (Mannheim, Germany). The nylon membranes were prehybridized for 3 hr at 65C in Solution A (5 × SSC, 5 × Denhardt's solution, 100 mg/ml denatured salmon sperm DNA, 0.5% SDS). They were then incubated for 16 hr at 55C in fresh Solution A containing 10 pmol/ml of DIG-labeled probes F or G, washed once with 2 × SSC, 0.1% SDS for 10 min at RT, and twice with 1 × SSC, 0.1% SDS for 20 min at 50C. The membranes were then washed with TBS and blocked with 1% fat-free powdered milk for 60 min at RT. They were then incubated with anti-DIG IgG conjugated with alkaline phosphatase (Boehringer–Mannheim) (diluted 1:4000 with TBS) for 2 hr at RT. After washing three times with TBS containing 0.02% Tween 20, the alkaline phosphatase was developed by NBT and BCIP. To control for the specificity of the hybridization, a tenfold excess of non-labeled oligonucleotides (probes F and G) was added to the hybridization solution.
Results
Spatiotemporal Expression of C-CAM in Placenta
The temporal expression pattern of C-CAM in the developing placenta was investigated by immunohistochemistry at the light microscopic level. No C-CAM was detected in the embryonic or extraembryonic cells on gestational Days 6.5 or 7.5. On day E8.5 it appeared in the trophoblasts of the ectoplacental cone (Figure 1A), and on Day E10.5 both spongiotrophoblasts (Figure 1C) and trophoblasts invading the maternal blood vessels (Figures 1D and 2A) of the decidua basalis were positively stained. The spongiotrophoblasts of the junctional zone continued to express C-CAM on Day E12.5 (Figure 1E and 2B) at a high level, but at this stage no staining was seen in the labyrinth or in secondary trophoblasts (Figure 1E). Labyrinth trophoblasts, and maternal blood lacunae of the junctional layer became stained by anti-C-CAM antibodies on Day E15.5 (Figure 1G), whereas most of the trophoblasts of the junctional layer no longer expressed C-CAM at this stage (Figure 1G). On Day E17.5, C-CAM expression was observed only in the labyrinth and in the lacunae of the junctional layer (Figure 1H). At higher magnification it could be seen that both the maternal surfaces of the labyrinth cytotrophoblasts and the endothelial cells of the embryonic capillaries were stained at this stage (Figure 2C). Only the cytotrophoblasts, and not the syncytiotrophoblasts, of the labyrinthine wall were stained. By Day E19.5 the expression pattern of C-CAM was the same as on Day E17.5. No staining was observed with nonimmune rabbit serum (Figures 1B and 1F).
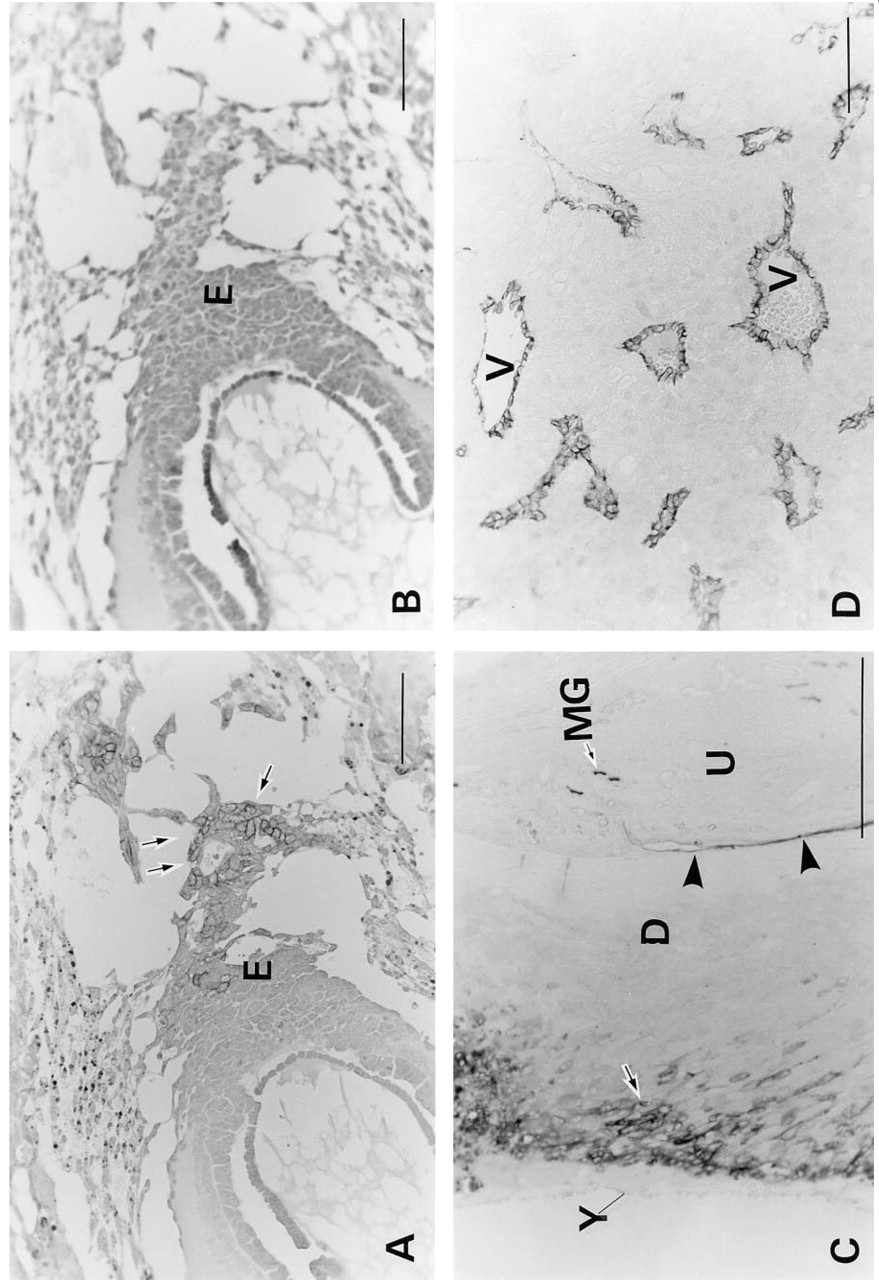
Light microscopic immunohistochemical staining for C-CAM in rat placentae (low magnification). Placental tissues were treated and immunoperoxidase-stained for C-CAM as described in Materials and Methods. (
In addition to the trophoblast and embryonic endothelial cell lineages, C-CAM was also expressed in maternal cells in the uterine cavity epithelium and in the metrial gland on Day E10.5 (Figure 1C), and in the endothelial cells of the endodermal sinus yolk sac on Day 17.5 (Figure 1H).
It is well known that pregnancy-specific glycoproteins (PSGs) which, like C-CAM, belong to the CEA gene family, are expressed at high levels in placentae of both humans and rodents (Rebstock et al. 1993; Ogilvie et al. 1989). Because there can be considerable immunological crossreactivity among different members of the CEA family, it was important to demonstrate that the staining observed with anti-C-CAM antibodies indeed reflected expression of C-CAM and not of PSG. For this purpose, we stained rat placentae of various stages with a PSG-specific antibody that has been shown to crossreact with rat PSG. As shown in Figure 3, this resulted in a completely different staining pattern compared with anti-C-CAM staining. The anti-PSG staining was cytoplasmically localized, whereas the anti-C-CAM antibodies stained the cell surface. At Day E12.5 the PSG staining was seen in the secondary giant trophoblasts, and only a few spongiotrophoblasts of the junctional layer were stained (Figure 3A). At Days E14.5 (Figure 3B) and E15.5 (Figure 3C), PSG was detected in the cytoplasm of the spongiotrophoblasts of the junctional layer and in secondary giant trophoblasts. At Day E17.5 the spongiotrophoblasts were very weakly stained and the secondary giant trophoblasts were not stained at all (Figure 3D). No staining of the labyrinth was seen at any stage. It can therefore be concluded that there was no crossreactive staining between C-CAM and PSG in rat placentae.
Subcellular Localization of C-CAM
Light microscopic immunohistochemistry demonstrated that C-CAM was localized to the cell surfaces of the trophoblasts at all stages of placental development (Figures 1 and 2). To determine the localization of C-CAM at higher resolution, we made immunoelectron microscopic analyses of placental tissues from gestational Days E12.5 and E17.5. At Day E12.5 C-CAM was located on the surface membranes of spongiotrophoblasts of the junctional layer (Figure 4A) and of trophoblasts invading maternal blood vessels (Figures 4B and 4C). Areas of close cell–cell contact were not significantly stained, but C-CAM was strongly expressed in areas of loose cell–cell association and on free cell surfaces. In the areas of loose cell–cell association, primarily microvillar structures were C-CAM-positive (Figures 4A and 4B). Invasive trophoblasts were also stained on microvilli facing the maternal blood vessels (Figure 4C).
At Day E17.5 the distribution of C-CAM was different from that on Day E12.5 at the light microscopic level (Figure 2C). At Day E17.5 it was primarily the luminal surfaces of the trophoblasts of the blood lacunae and the embryonic endothelial cells that showed strong C-CAM staining. At the electron microscopic level it was seen that the microvilli of the labyrinth trophoblasts, which protruded into the maternal blood circulation, were stained. The embryonic endothelial cells expressed C-CAM both on the luminal surfaces and intracellularly, presumably in intracellular membranes (Figures 4D and 4E).
Molecular Analyses of C-CAM
The expression of C-CAM was investigated at the protein level by immunoblotting. At Days E12.5 and E14.5 C-CAM appeared as a broad band with an apparent molecular mass of 110–170 kD. The broad band probably reflected a high but heterogeneous degree of glycosylation. All detectable C-CAM could be extracted by Triton X-114, and none was left in the insoluble tissue residue. All of the extracted C-CAM partitioned into the detergent phase of Triton X-114 (Figure 5), demonstrating that rat placental C-CAM behaves as an integral membrane protein.
No discrete bands were seen in the broad C-CAM band, and therefore this analysis did not reveal whether different C-CAM isoforms were expressed. Because no isoform-specific antibodies that could be used for this purpose were available, we investigated C-CAM expression at the mRNA level. This was done in Days E12.5, E14.5 and E17.5 placentae using the RT-PCR technique. The primer set of F5,Xho-1/B6,Xho-1-ampli-fied fragments of approximately 1500 BP (Figure 6A) at all the developmental stages, whereas the primer combination F3/B6,Xho-1 amplified a poorly resolved doublet containing two bands of 638 and 585 BP (Figure 6A), respectively. This suggested that transcripts for both C-CAM1 and C-CAM2 occurred.
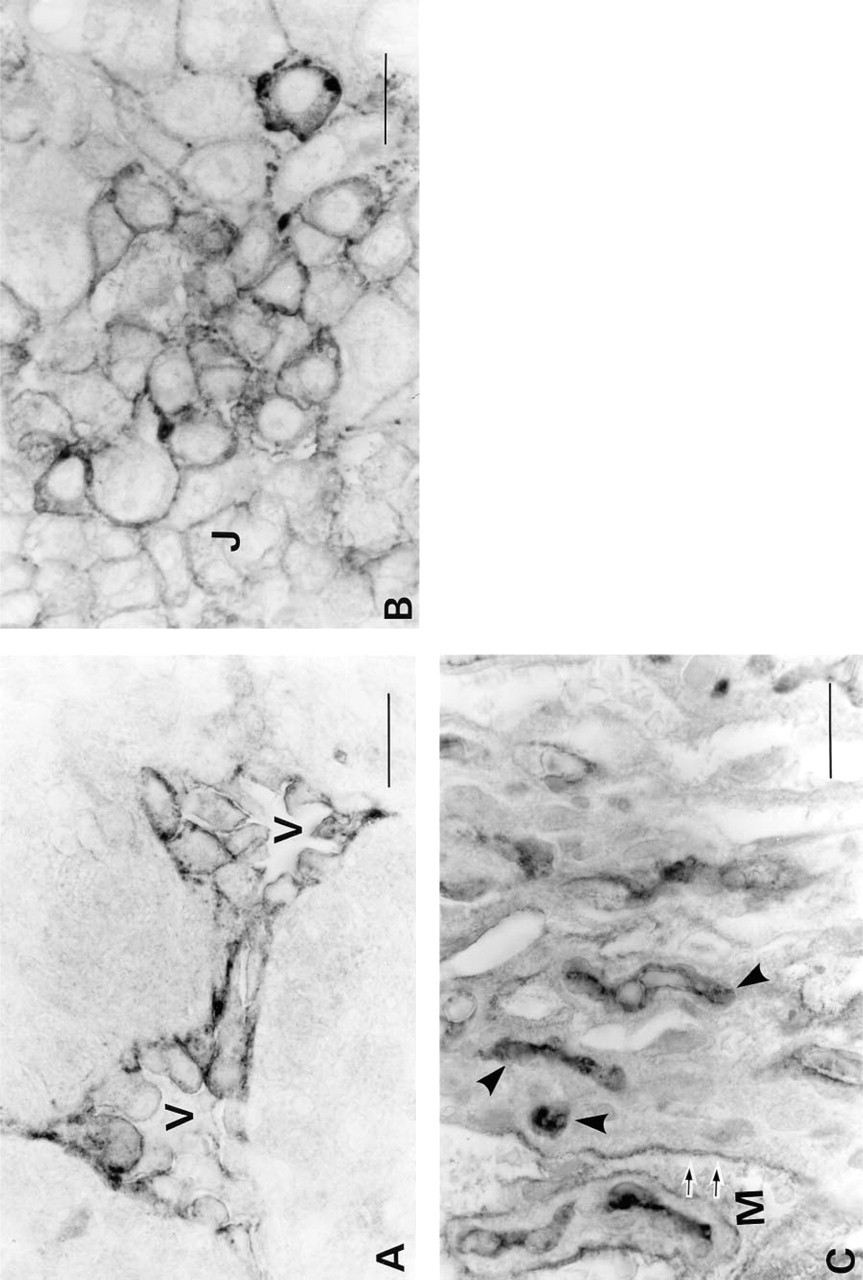
Light microsopic immunohistochemical staining for C-CAM in rat placentae (high magnification). Placental tissues were treated and immunoperoxidase-stained for C-CAM as described in Materials and Methods. (
It was not possible to determine if the 1500-BP band, amplified by the primer set F5,Xho-1/B6,Xho-1, contained more than one band. Therefore, this product was reamplified using the primer set combination F3/B6,Xho-1. This resulted in two fragments of 638 and 585 BP (Figure 6B), which were seen in rat placentae of all developmental stages. Southern blot analysis of these cDNAs demonstrated that probe F hybridized with the 638-BP fragment, and probe G hybridized with both the 638 BP and the 585 BP fragment (Figure 6D, Lanes 1 and 3). Nonlabeled oligonucleotides abolished the hybridization of the labeled probes F and G (Figure 6D, Lanes 2 and 4). These results showed that the 638-BP fragment represented C-CAM1 and the 585-BP fragment represented C-CAM2.
Discussion
Employing specific antibodies in light and electron microscopic analyses, we found that the cell adhesion molecule C-CAM is expressed in rat placenta in a characteristic spatiotemporal pattern in two cell lineages of embryonic origin, trophoblasts and capillary endothelial cells. C-CAM is a member of the CEA gene family, which comprises several structurally related proteins (Thompson et al. 1991), and it has previously been demonstrated that rat placenta expresses proteins belonging to the PSG subgroup of the CEA family (Rebstock et al. 1993; Ogilvie et al. 1989). However, antibodies recognizing rat PSGs gave a completely different spatiotemporal staining pattern, demonstrating that the antibodies against C-CAM did not crossreact with PSG. The expression of C-CAM was confirmed at the RNA level, which revealed expression of both major isoforms, C-CAM1 and C-CAM2.
Previous investigations by one of us showed that C-CAM is expressed early in the preimplantation embryo by the trophectoderm of the blastocyst (Svalander et al. 1987). The trophectodermal expression disappeared at the time of implantation, and no C-CAM expression was seen in the implanted embryo or the trophoblasts up to 8 days of gestation (Svalander et al. 1990). This was confirmed in the present investigation, in which the first appearance of C-CAM after implantation was noted on Day E8.5, in the trophoblasts of the ectoplacental cone. From the gestational Day 9 the developing trophoblasts exhibited a characteristic dynamic expression pattern of C-CAM. The downregulation of C-CAM in the trophoblasts at the time of implantation and its reappearance at a later developmental stage agree well with the dynamic expression pattern that has been observed for many cell adhesion molecules during embryonic development. A well-known example is the disappearance and reappearance of N-CAM and N-cadherin during migration, reorganization and differention of neural crest cells (Thiery et al. 1988). The absence of C-CAM early after implantation may be a prerequisite for trophoblast migration and proliferation.
At Day E15.5 the rat placenta has acquired its mature histological structure as a result of cell proliferation, cell differentiation, and reorganization of the tissue architecture. Two significant features of the expression pattern of C-CAM of relevance for placental development and function could be distinguished. First, C-CAM was expressed in cells that are highly active and instrumental in forming the placental histostructure, i.e., the trophoblasts of the ectoplacental cone, the spongiotrophoblasts of the junctional zone, and the trophoblasts invading the maternal blood vessels. However, C-CAM was not expressed in the trophoblasts of the labyrinth before the mature placental structure was reached. When C-CAM appeared in the labyrinthal trophoblasts it was no longer seen in the trophoblasts of the junctional layer. This expression pattern suggests that C-CAM has important roles in the development of the tissue structure of the placenta. Second, in the mature placenta C-CAM was expressed both in the trophoblasts lining the maternal blood vessels and in the endothelial cells lining the fetal capillaries. These cells are important for the function of the mature placenta but are probably not involved in tissue remodeling. This expression pattern suggests that C-CAM is important for the physiological functions of the placental vasculature.
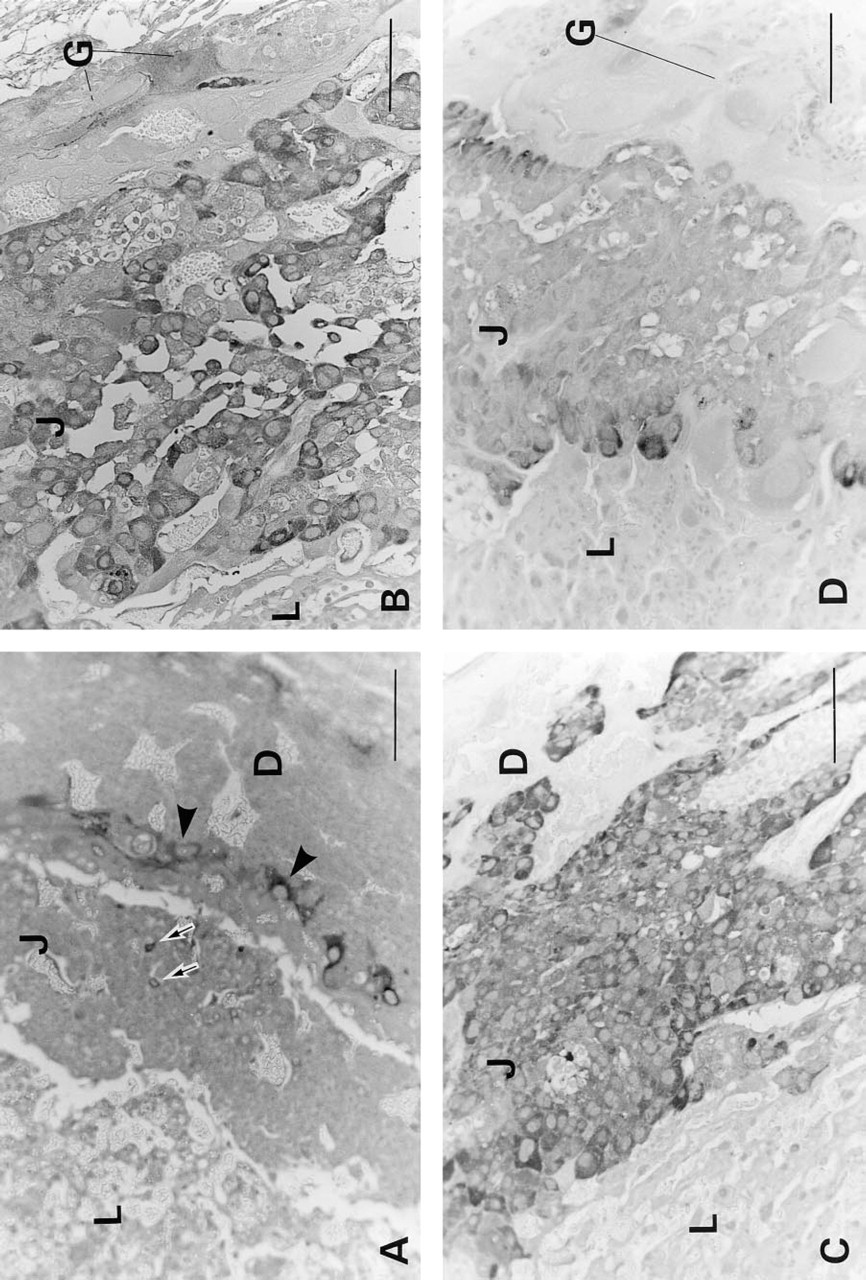
Light microscopic immunohistochemical staining for PSG in rat placentae. Placental tissues were treated and immunoperoxidase-stained for PSG as described in Materials and Methods. (
Much remains to be learned about the physiological and cell biological functions of C-CAM, but available data from other systems agree with the idea that it can have different functions in various cells. The best documented property of C-CAM is its ability to mediate cell–cell adhesion by homophilic binding (Ocklind and Öbrink 1982; Tingström et al. 1990). It has recently been found that both of the isoforms, C-CAM1 and C-CAM2, can mediate adhesion (Lukka et al. 1995; Olsson et al. 1995), and that this is caused by reciprocal binding of the D1 domains of C-CAM molecules on opposing cells (Wikström et al. 1996). However, additional functions for C-CAM have been suggested. It has been claimed that C-CAM1 is an ecto-ATPase, because the amino acid sequence of C-CAM was identical to that of a cloned putative ecto-ATPase (Lin and Guidotti 1989), but it has not been possible to verify that purified C-CAM has ATPase activity. It is now believed that the ecto-ATPase activity resides in a distinct protein that co-purifies with C-CAM (Stout et al. 1995). Another activity that has been ascribed to C-CAM1 is active transport of bile acids (Sippel et al. 1993, 1994). However, the bile acid-transporting activity is probably mediated by a different protein, but it remains a possibility that C-CAM can regulate this activity by participating in supramolecular complexes in the plasma membrane. This idea is supported by the recent discovery that another membrane transporter, a voltage-gated sodium channel protein in brain, is associated with two subunits having immunoglobulin-like domains (Isom et al. 1994). It has been suggested that they function as adhesion molecules that organize the sodium channel in specific locations in neurons (Isom and Catterall 1996). Supramolecular complexes of C-CAM have been described in several cell types (Hunter et al. 1996). Finally, it has recently been found that C-CAM1 is a negative regulator of cellular growth and that it can suppress tumorigenesis of prostate and colon carcinoma cells (Hsieh et al. 1995; Kunath et al. 1995).
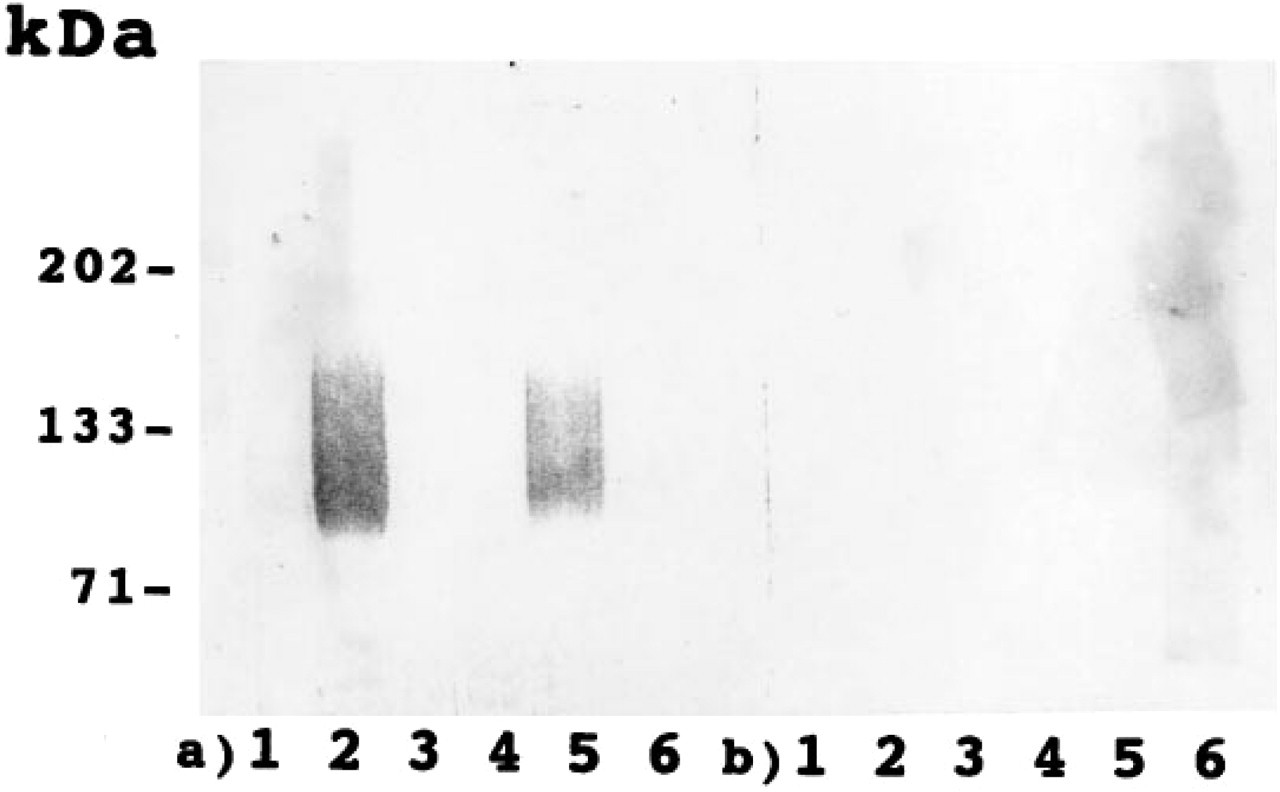
Immunoblotting of rat placenta. Rat placental tissue was solubilized and phase-separated in Triton X-114. The samples were separated by SDS-PAGE under reducing conditions and analyzed by immunoblotting with antibodies against C-CAM (
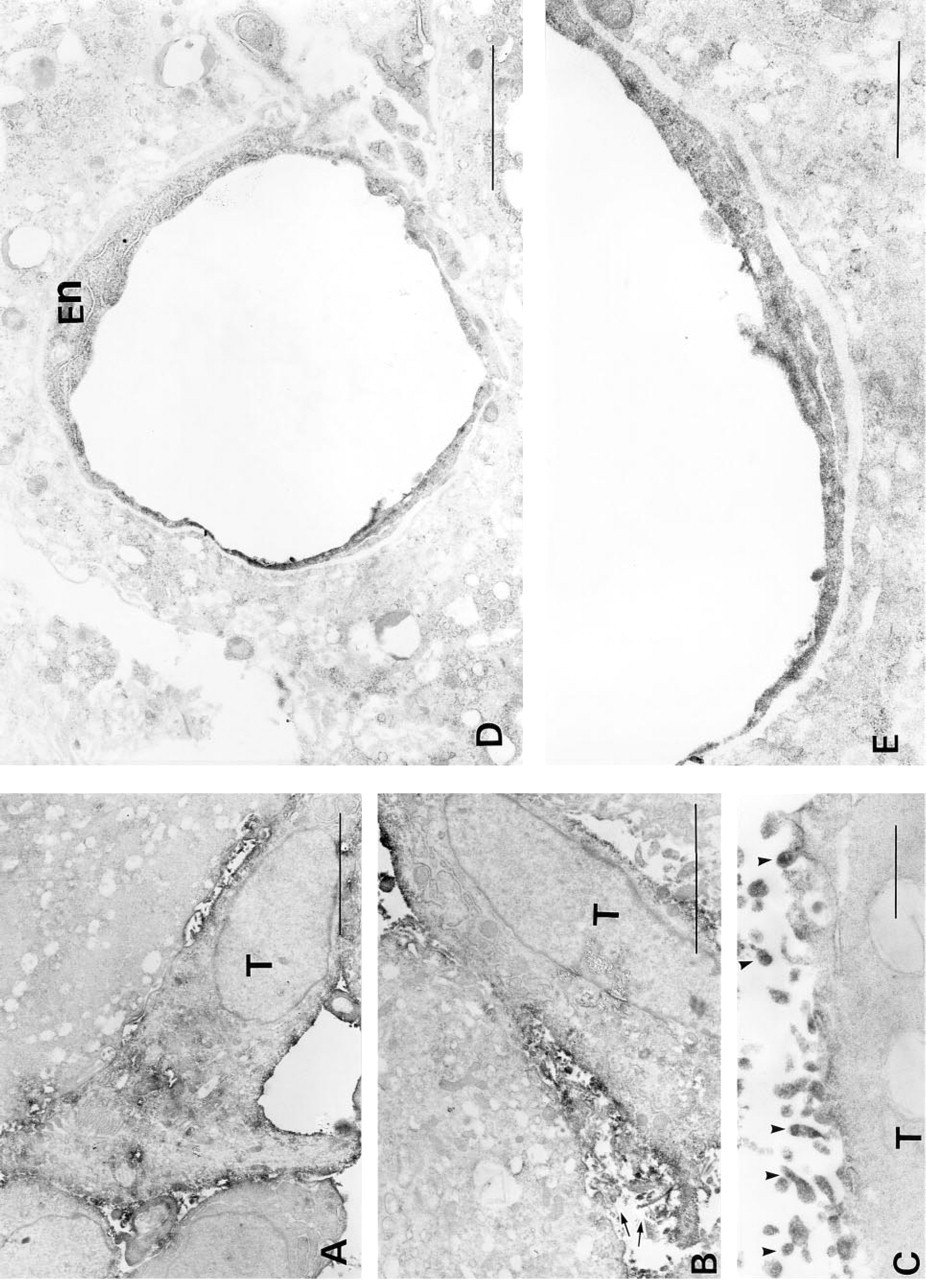
Electron microscopic immunohistochemical staining for C-CAM in rat placentae. Placental tissues were treated and immunoperoxidase-stained for C-CAM as described in Materials and Methods. (
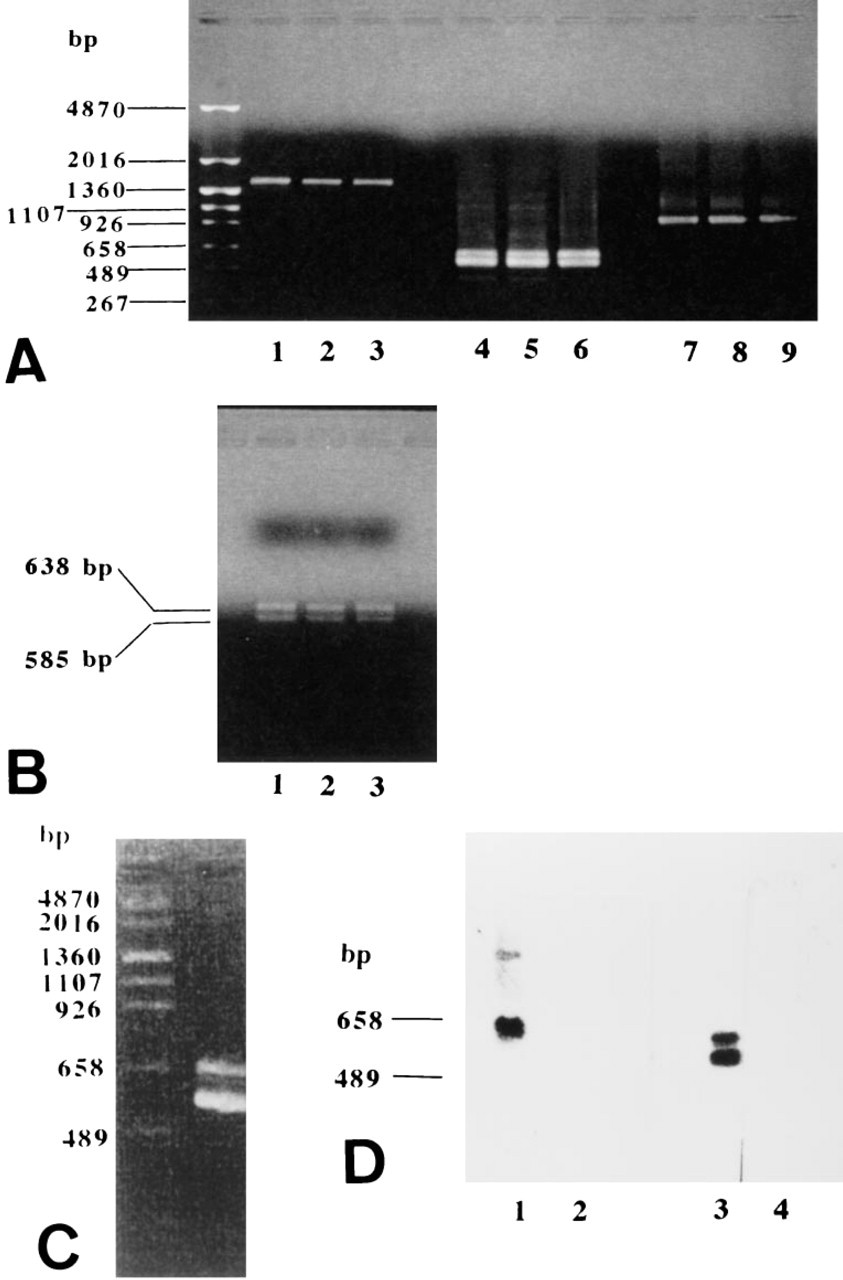
RT-PCR and Southern blot analyses of rat placentae. PolyA(+) RNA was extracted, transcribed, amplified, electrophoresed, and hybridized as described in Materials and Methods. (
It appears plausible that C-CAM-mediated adhesive interactions between the ectoplacental cone trophoblasts, the spongiotrophoblasts, and the vessel-invading trophoblasts are involved in development of the placental tissue structure. C-CAM probably does not contribute to strong intercellular bonds in the same way as cadherins, because it was not localized in close cellular contacts but in loosely organized contact regions with extensive microvillar structures. Microvillar localization of C-CAM has been observed in several other cell types (Odin et al. 1988). One appropriate example is the junctional epithelium of the teeth. This stratified epithelium, which attaches to the enamel of the teeth, has a strikingly different morphology at the ultrastructural level compared to the adjacent gingival epithelium (Hashimoto et al. 1986; Listgarten 1972). It consists of loosely associated cells with wide intercellular spaces. Very few desmosomes are seen, but the intercellular spaces are filled with cellular microvilli that contact each other. Interestingly, we have found high expression levels of C-CAM in these areas, contrasting to the very low expression in the surrounding gingival epithelium (Lüning et al. 1995). Furthermore, the junctional epithelium does not express any E-cadherin (Terling et al. submitted for publication). C-CAM-mediated intervillar adhesion in the junctional epithelium is probably of a dynamic nature, necessary for the integrity of the tissue but at the same time allowing local cell motility. A similar function for C-CAM in the ectoplacental cone trophoblasts, spongiotrophoblasts, and invasive trophoblasts is an attractive possibility.
Another possibility is that C-CAM's major function in the developing placenta involves signal transduction, which may regulate both proliferative and secretory activities of the trophoblasts. It has indeed been found that C-CAM1 is a signal transduction molecule that can bind protein tyrosine kinases, e.g., c-src (Brümmer et al. 1995), to phosphorylated tyrosine residues in its cytoplasmic domain. Furthermore, the calcium-regulated protein calmodulin can bind to the cytoplasmic domains of C-CAM1 and C-CAM2 and regulate their supramolecular organization (Edlund et al. 1996a; Brümmer et al. 1995). The cytoplasmic domain of C-CAM1 has been demonstrated to be essential for both its growth regulatory properties (Hsieh et al. 1995; Kunath et al. 1995) and for bile acid transport activity (Sippel et al. 1994). Strictly regulated growth control of the actively proliferating and invading trophoblasts may be crucial to prevent placental malformation and tumor formation.
The function of C-CAM in the cells lining the vessel walls of the mature placenta is probably not mediated by cell–cell contacts, since C-CAM was preferentially found on their free apical surfaces. However, intervillar contacts are still a possibility in these locations. A likely role for C-CAM in these cells is regulation of secretory processes, but it might also influence proliferatory activity.
All tissues and cells in which C-CAM has been found express both C-CAM1 and C-CAM2 simultaneously. This was also found in the placenta throughout its development. In most cell types the shorter isoform, C-CAM2, dominates. The PCR data of the present investigation, however, indicated that the two C-CAM isoforms were expressed at fairly equal levels in the placenta. This is interesting because there are indications that not only the expression levels but also the ratio between C-CAM1 and C-CAM2 are of functional importance. One possibility is that C-CAM2 regulates the activity of the cytoplasmic domain of C-CAM1. The recent finding that C-CAM can form dimers (Hunter et al. 1996) supports such an idea.
Footnotes
Acknowledgements
Supported by the Swedish Medical Research Council (project number 05200), the Petrus and Augusta Hedlunds Foundation and the Karolinska Institute, and by a grant-in-aid for the Science Research Promotion Fund from the Japan Private School Promotion Foundation.
We thank Mr Isao Naito for photographic assistance and Ms Hideko Shimizu and Ms Hiyomi Murakami for technical assistance.