
Abstract
Type II procollagen is synthesized in two forms generated by the alternative splicing of its precursor mRNA. The alternatively spliced domain, exon 2, encodes the 69-amino-acid cysteinerich region of the NH2 propeptide. Studies of mRNA expression have shown that the longer form, designated Type IIA procollagen, is synthesized by chondroprogenitor cells and various noncartilaginous tissues. The shorter form, Type IIB procollagen, is synthesized by differentiated chondrocytes. As the initial step in our investigations of the function of the Type IIA procollagen, the protein domain corresponding to exon 2 was created as a recombinant fusion protein and used to raise antibodies in rabbits. The resulting antiserum was specific for Type IIA procollagen NH2 propeptide as shown by ELISA, Western blotting, and immunofluorescent co-localization with the triple-helical domain of Type II collagen. Type IIA procollagen was identified in tissue culture medium of 54-day human fetal ribs. Confocal microscopy was used to localize the Type IIA NH2 propeptide in Day 50 and 53 human embryos. In the digital rays of the developing hand, where only Type IIA procollagen mRNA was detected, Type IIA procollagen NH2 propeptide was observed in the extracellular matrix. The presence of Type IIA procollagen NH2 propeptide was observed in the cartilage of the developing long bones of the lower arm and vertebral bodies even though these tissues synthesize Type IIB mRNA at this developmental stage. Type IIA procollagen NH2 propeptide was localized in the developing trachea, a cartilage that does not undergo endochondral bone formation. Type IIA NH2 propeptide was also localized in noncartilaginous tissues known to synthesize Type IIA mRNA, such as the intervertebral area, perichondrium, notochordal sheath, and neuroepithelium of the otic vesicle. In most tissues, co-localization with antiserum against the triple-helical domain of Type II collagen was observed. Positive immunoreactivity with the Type IIA NH2 propeptide antiserum indicates, for the first time, that this propeptide is present in the tissue. Co-localization of NH2 propeptide antibodies with the triple-helical domain of the collagen molecule suggests that Type IIA procollagen is intact in the extracellular matrix of these tissues. Taken together, these results strongly suggest that around cells that synthesize Type IIA procollagen mRNA, Type IIA procollagen NH2 propeptide is secreted and deposited into the extracellular matrix. In light of these results, we predict that Type IIA procollagen plays a role in differentiation of tissues that augments its purely architectural function.
Keywords
T
Type II collagen, like the other fibrillar collagens Types I and III, is initially synthesized as a procollagen containing propeptides at both amino (NH2) and carboxy (COOH) termini, which are generally removed before deposition of fibrils into the tissue. Unlike other fibrillar collagens, Type II procollagen is synthesized in two forms generated by alternative splicing of the precursor mRNA (Ryan and Sandell 1990). The alternatively spliced domain, exon 2, encodes a 69-amino-acid, cysteine-rich NH2 propeptide. The expression of Type II procollagen splice forms is regulated and associated with the developmental stages of various tissues. For example, the long form, encoded by Type IIA procollagen mRNA, is synthesized by skeletal progenitor cells and noncartilaginous embryonic tissues, whereas the shorter form, Type IIB procollagen mRNA, is synthesized by mature chondrocytes (Sandell et al. 1991; Ng et al. 1993; Sandell et al. 1994; Lui et al. 1995). Therefore, the Type II collagen immunolocalized in noncartilaginous tissues is or is derived from Type IIA procollagen. To date, the only way to differentiate the procollagen forms is by in situ hybridization to mRNA.
A protein domain similar to the Type IIA procollagen NH2 propeptide is present in procollagens I, III, and V, thrombospondins 1 and 2, and von Willebrand factor (Ryan and Sandell 1990). In collagens I and III, most of the NH2 propeptides are usually removed before deposition of the fibrillar collagen domain in the extracellular matrix (Olsen 1991). However, NH2 propeptide has been immunolocalized in embryonic skin (Fleischmajer et al. 1983). Various functions have been suggested for the NH2 propeptide, including regulation of fibril diameter (Fleischmajer et al. 1990) and, when removed, as a feedback regulator of collagen synthesis (Fouser et al. 1991). Recently, two new proteins have been isolated that contain multiple copies of a domain very similar to the Type IIA procollagen NH2 propeptide, the Drosophila protein sog (Francois et al. 1994) and the Xenopus protein chordin (Sasai et al. 1994). These proteins function in dorsal–ventral patterning by binding to and regulating the presentation of members of the TGF-β superfamily, i.e., the bone morphogenetic proteins (Piccolo et al. 1996). Therefore, it is plausible that the Type II collagen NH2 propeptide has a similar role in skeletal patterning.
Preceding any studies on the functional role of Type IIA procollagen it is necessary to establish whether Type IIA procollagen is deposited and remains in the extracellular matrix. We have previously shown that Type IIA procollagen can be detected in the media of primary cultures of young human costal chondrocytes (Sandell et al. 1991). However, isolation of Type IIA procollagen and production of antisera for its immunolocalization have been hampered by the inability to extract sufficient amounts of Type IIA procollagen from tissue and the lack of a cell line or primary cell culture capable of producing adequate amounts of this protein. To overcome these problems, exon 2 encoding the unique portion of the NH2 propeptide was produced as a recombinant fusion protein and expressed in bacterial cells. The recombinant protein was used to immunize rabbits and the resulting polyclonal antiserum was employed to localize Type IIA procollagen in selected human embryonic tissues.
Materials and Methods
Construction of Exon 2 Fusion Protein
PCR primers used for amplification of exon 2 were designed to amplify a 207-base pair (
The strategy for the ligation of the PCR products into the pGEX-2T vector for protein expression is shown in Figure 1B. This ligation places the codon for Gln-87 (the first residue of the exon 2) in frame right after the BamHI cloning site, adding an extra Gly-Ser dipeptide between GST and the exon 2 sequence. Thrombin cleavage of GST–exon 2 fusion protein occurs between Arg and this dipeptide, resulting in an exon 2 protein with an additional two amino acids at the amino terminal end. The ligated constructs were transformed into competent E. coli DH5α cells. Transformants containing inserts were detected by PCR and preservation of a reading frame was confirmed by sequencing across the junction of GST and exon 2.
Expression and Purification of GST Fusion Protein
A single colony of DH5α cells carrying the plasmid of interest was grown overnight at 37C in 500 ml of LB containing 100 μg/ml ampicillin until the A600 reached 1.0–1.2. Bacterial cells were induced with 0.1 mM isopropyl-β-
SDS-PAGE was performed under standard procedures (Laemmli 1970). Routinely, slabs of 1-mm thickness were used with polyacrylamide concentrations of 12 or 15% with 5% stacking gels using a Bio-Rad Mini-PROTEAN II dualslab cell apparatus (Bio-Rad; Hercules, CA). Gels were stained with Coomassie Brilliant Blue R-250 (Bio-Rad) or analyzed by Western immunoblotting.
Thrombin Cleavage of the Fusion Protein and Isolation of the Exon 2 Protein
Fusion protein was cleaved by thrombin for 16 hr at RT (RT) with an enzyme:substrate ratio of 10 U/mg fusion protein in 10 mM glutathione, 50 mM Tris-HCl, pH 8.0. After complete digestion, glutathione was removed by extensive dialysis against PBS. The GST was then removed by purification through a glutathione–Sepharose 4B column. The exon 2 protein was found in the flow-through and was analyzed by SDS-PAGE or Western blot. The N-terminal amino acid sequence of the exon 2 polypeptide was confirmed using a Porton Model 2090 E instrument with on-line reversed-phase HPLC detection on phenylthiohydantoin amino acid derivatives.
Production of Polyclonal Antibodies to the Exon 2 Propeptide
New Zealand White rabbits were immunized with purified GST–exon 2 fusion protein. Briefly, 400 μg of fusion protein in 1 ml isotonic saline was emulsified with 1 ml of Freund's complete adjuvant and injected sc in multiple sites at the back of each animal. Three booster injections of 100 μg of cleaved and purified exon 2 polypeptide in Freund's incomplete adjuvant were administered after 3, 6, and 9 weeks. Blood was collected from each rabbit after each booster injection. The antiserum with the highest titer was used for further experiments.
Antibody Titer Determination
The antiserum obtained at various time intervals after immunization and from different rabbits was tested for antibody titer by indirect ELISA. The optimal concentration of antigen to be used for coating microtiter plates was determined in preliminary experiments. ELISA reactions were carried out in a reaction volume of 50 μl. Microtiter plates (Costar, EIA/RIA; Cambridge, MA) were coated with 1 μg/ml (50 ng/well) GST–exon 2 fusion protein in PBS (pH 7.3) overnight at 4C. Purified GST was used as a control. After three washes with buffer (PBS, 0.1% v/v, Tween-20) the plates were incubated with 200 μl/well blocking buffer (PBS, 0.1% Tween-20, 3% BSA) for 2 hr at RT. The plates were washed again and antiserum at different dilutions (from 1:500 to 1:100,000) was added to each well and incubated at RT for 2 hr. The plates were washed and a secondary antibody, goat anti-rabbit IgG conjugated with alkaline phosphatase, at a dilution of 1:5000 (Bio-Rad), was added and incubated at RT for 2 hr. The plates were washed again and 75 μl of substrate solution, 3 mM p-nitrophenyl phosphate (NPP) (Sigma; St Louis, MO) in 0.05 M Na2CO3 and 0.05 mM MgCl2 was added to each well. After 1 hr the reaction was terminated by adding 25 μl of 0.5 M NaOH. The optical density values at 405 nm were measured with an automatic ELISA reader (Dynatech Laboratories; Chantilly, VA). Each sample was run in duplicate. The ELISA results were determined by taking the mean value of the two absorbance readings and then subtracting the mean value of the two absorbance readings with the GST protein.
Each experiment was repeated twice. The antibody titer was calculated as the antiserum dilution that would give an absorbance of 0.5 under the conditions described above.
Sensitivity Assay
Anti-exon 2 antibody of highest titer obtained during the immunizations was used. Each well of a microtiter plate was coated overnight at 4C with 50 μl of each antigen at concentrations from 5 pg/well to 200 ng/well. The wells were washed and vacant sites were blocked with blocking buffer. After incubation (as described above) with primary and appropriate secondary antibodies, plates were washed and substrate solution was added and incubated at RT. After 2 hr the reaction was stopped by addition of 25 μl of 0.5 N NaOH per well and the A405 measured as described above.
Immunoblot Analysis
For Western blot analysis, protein samples of reduced E. coli lysate, purified GST–exon 2 fusion, and exon 2 proteins were electrophoresed as described above. Prestained molecular weight markers were run on each gel. The gels were electroblotted onto PVDF (Immobilon-P; Millipore, Bedford, MA) membranes using a Trans-Blot SD cell apparatus (Bio-Rad). The membranes were blocked for 1 hr at RT with blocking buffer (TTBS) (100 mM Tris-HCl, pH 7.4, 150 mM NaCl, 0.1% Tween 20), containing 0.3% BSA and then incubated with rabbit polyclonal antibodies at 1:1000 dilution in the same buffer for 1 hr. After four washes of 10 min in TTBS buffer, a complex of goat anti-rabbit IgG-alkaline phosphatase was used (dilution 1:5000) to detect the first antibody. After four further washes of 10 min in TTBS, the staining reaction was carried out in Western Blue stabilized substrate solution (Promega; Madison, WI).
Specificity of Type IIA Antiserum
Culture medium from human skin fibroblasts labeled with L- [2,3,4,5-3H]-proline (Amersham; Arlington Heights, IL) was obtained from Barbara Starman and Dr. Peter Byers (University of Washington). To 700 μl labeled culture medium was added 415 μl ethanol and proteins were pelleted by centrifugation for 4 min at 10,000 rpm in a table-top centrifuge (Tomy MTX-150; Tomy Seiko, Tokyo, Japan). Supernatant was discarded and the pellet was dissolved in 150 μl sample buffer [50 mM Tris-Cl, pH 6.8, 100 mM dithiothreitol (DTT), 2% SDS, 0.1% bromphenol blue, 10% glycerol] and 25 μl was loaded in a 6% SDS-polyacrylamide gel. For detection of radiolabeled proteins, part of the gel was fixed in fixing solution (isopropanol:acetic acid:water 25:10: 65) for 15 min, soaked in Amplify (Amersham), and dried under vacuum at 80C. After 2 days' exposure at −80C, film was developed. The second part of the gel was transferred onto PVDF membrane for immunoblot analysis. The strips of the membrane were blocked for 1 hr at RT in TTBS buffer and incubated with monoclonal antibody against the α 1-chain of Type I procollagen at 1:500 dilution (Developmental Studies Hybridoma Bank; Johns Hopkins University, Baltimore, MD) and with Type IIA procollagen antiserum at the same dilution for 1 hr at RT. After four washes of 10 min in TTBS and incubation with appropriate secondary antibodies conjugated with alkaline phosphatase, strips were stained in Western Blue stabilized substrate solution.
Human fetal ribs (54 days' gestation, Stage 22) were provided by the Central Laboratory for Human Embryology at the University of Washington. Tissue was minced and incubated at 37C in Dulbecco's modified Eagle's medium (Bio Whittaker; Walkersville, MD) supplemented with 5% fetal calf serum (HyClone Laboratories; Logan, UT), 100 U/ml streptomycin, and 100 μg/μl penicillin. After 2 days the medium was changed to serum-free supplemented with 50 μg/ml ascorbate (Sigma), 100 μg/ml β-aminopropionitrile fumarate (Sigma), and 20 μCi/ml of
In Situ Hybridization to mRNA
Tissues used in this study were human embryos, Day 50 of gestation (in situ hybridization and immunohistochemistry) and Day 53 (immunohistochemistry) (embryonic Stages 20–22), provided by the Center Laboratory for Human Embryology (University of Washington, Seattle, WA). Tissues were frozen in OCT compound (Miles Laboratories; Elkhart, IN), sectioned at 8–10 μm with a cryostat, and stored at −70C until used. For detection of Type IIA procollagen mRNA, a 200-
Immunofluorescence Staining
Two polyclonal antibodies were used. Rabbit antiserum against human Type IIA procollagen NH2 propeptide (characterized in this manuscript) and rat antiserum against bovine Type II triple-helical collagen (kindly provided by Dr. Mike Cremer) (Cremer and Kang 1988). Frozen sections (8–10 μm) mounted on polylysine-coated slides (Fisher Scientific; Pittsburgh, PA) were fixed in 4% paraformaldehyde for 10 min at RT, and incubated with 1 mg/ml hyaluronidase (Bovine Testes, Type IV-S; Sigma–Aldrich Chemical, St Louis, MO) for 1 hr at 37C. Sections were blocked in PBS containing 5% (v/v) normal donkey serum (blocking buffer; Jackson ImmunoResearch Laboratories, West Grove, PA) for 1 hr at 37C. All primary antibodies were diluted in PBS containing normal donkey serum (1% v/v). Antiserum to Type NH2 propeptide was used at a dilution of 1:400 and the triple-helical Type II was used at 1:50. For double immunostaining, primary antibodies (rabbit anti-human IIA and rat anti-bovine II) were mixed completely and incubated overnight at 4C with the sections. After washing in PBS, sections were incubated sequentially with appropriate secondary antibodies (cyanine 3-conjugated donkey anti-rabbit IgG F(ab') fragment at a dilution of 1:200 and FITC-conjugated donkey anti-rat IgG F(ab') fragment at a dilution of 1:100, (Jackson ImmunoResearch Laboratories) for 30 min at RT. After washing, sections were mounted in fluorescent mounting medium (Vector Laboratories, Burlingame, CA) and viewed with a Nikon Optiphot microscope using DM510 (for FITC) and DM580 (for cyanine 3) filters. Preimmune rabbit and normal rat serum were used as controls as a substitute for primary antibodies. In addition, fluorochromes were interchanged conjugated to the appropriate secondary antibodies.
Confocal Microscopy
Images were collected on a Bio-Rad MRC600 scanning laser confocal microscope mounted on a Nikon Optiphot base. Data were collected using either a Nikon 20X/0.50 n.a. dry objective or a x40/0.70 n.a. dry objective. The Bio-Rad A1–A2 filters were used with an argon laser light source producing excitation at 514 nm and collecting emissions at 520–560 nm (green, FITC) or <600 nm (red, Cy-3). Optical sections were about 2 μm in thickness with the x20 objective and 1 μm with the x40 objective. Full frame (768 × 512) 8-bit images were collected for analysis and overlaid in 24-bit RGB using Adobe Photoshop v.3.0.
Results
Cloning and Expression of the Type II Collagen Gene Exon 2 cDNA
PCR was used to amplify a 207-
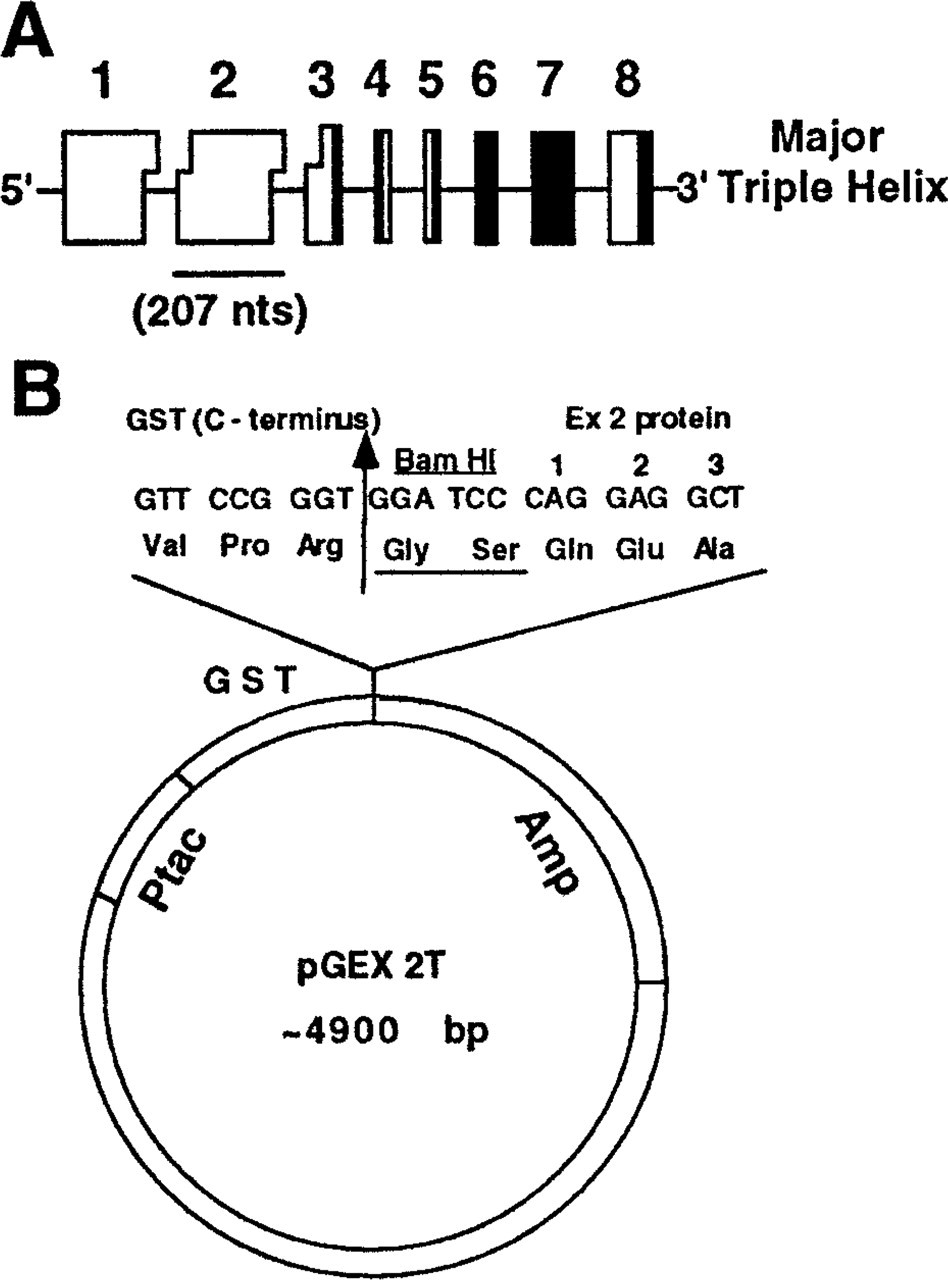
Cloning strategy and expression vector used. (
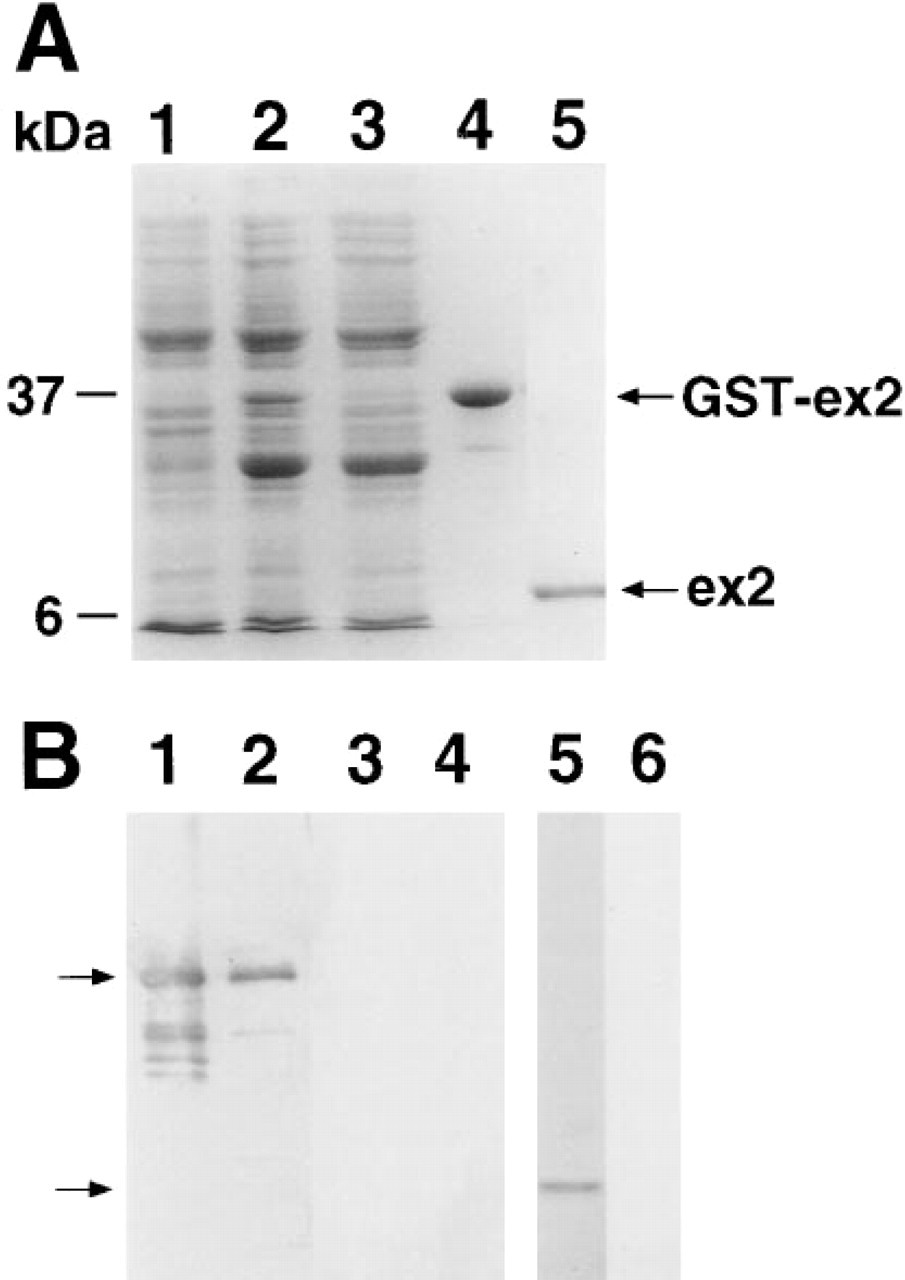
Expression and purification of exon 2 protein. (
Liberation of the exon 2 propeptide from its fusion protein was achieved enzymatically via incubation with thrombin. Thrombin is a serine protease that specifically cleaves the fusion protein at the Arg-Gly bond (site designed into the construct) (see Figure 1B). Thrombin was added directly to the purified fusion protein (10 U/mg of fusion protein) and incubation was carried out at RT for 16–24 h. SDS-PAGE was used to monitor the completeness of digestion. The digestion mixture was loaded directly onto a Glutathione Sepharose 4B column and the purified propeptide was found in flow-through. N-terminal sequence analysis of exon 2 protein confirmed the presence of a single species exactly matching the primary structure of the exon 2 domain. The absence of secondary sequences in the protein sequencing protocol was also evidence that the exon 2 domain was not significantly proteolyzed during its purification. This is supported by the single species seen on SDS-PAGE at the expected molecular weight, 7.5 kD for exon 2 propeptide (Figure 2A, Lane 5). Some disulfide binding occurs in the isolated exon 2 propeptide because the migration on SDS-PAGE is somewhat faster if the gel is run unreduced (data not shown).
Antiserum to GST–Exon 2 Fusion Protein
Western blot analysis shows that the antiserum reacted with GST fusion protein from the E. coli lysate (Figure 2B, Lane 1) whereas no reactivity was detected by preimmune serum (Figure 2B, Lane 3). Light reaction products detected in the E. coli lysate are most likely proteolytic fragments of the GST fusion protein. The antiserum reacted with purified GST–exon 2 fusion and exon 2 proteins (Figure 2A, Lanes 2 and 5, respectively). No reactivity was detected by preimmune serum (Figure 2B, Lanes 4 and 6).
Specificity of Type IIA Procollagen NH2 Propeptide Antiserum
Type IIA procollagen was identified in culture medium of Day 54 fetal ribs. Figure 3A shows an auto-radiograph of proline-labeled proteins reduced with β-mercaptoethanol and separated on a 6% SDS-PAGE (Figure 3A, Lane 1). Lanes 2 and 3 (Figure 3A) were transferred to blotting membrane and immunoreacted with antiserum to Type IIA procollagen NH2 propeptide (Figure 3A, Lane 2) or preimmune serum (Figure 3A, Lane 3). The band shown in Lane 2 (Figure 3A) is collagenase-sensitive, reacts with Type II triple-helical antiserum, and was not observed when samples were unreduced, strongly suggesting that it is pNC procollagen. To compare the migration of the Type IIA procollagen with mature collagen, Type II collagen α-chains were run in Lane 4 (Figure 3A) and immunoreacted with the rat anti-collagen triple-helical domain antiserum.
Protein sequence comparisons indicated that human Type IIA NH2 propeptide is most closely related to human α1(I) NH2-propeptide (Ryan and Sandell 1990). To determine whether there is crossreactivity between the α1(I) and α1(II) procollagens, radiolabeled proteins from human skin fibroblast culture medium were separated by electrophoresis and subjected to Western blot analysis using antisera to both Type I amino propeptide and Type IIA amino propeptide. The Type I procollagen antiserum reacts with a 19-amino-acid epitope on the amino-terminal side of the propeptide cleavage site (Foellmer et al. 1983). An autoradiograph of fibroblast culture medium labeled with [3H]-proline is shown in Figure 3B, Lane 1. Lanes 2 and 3 (Figure 3B) were transferred to blotting membrane and reacted with Type I procollagen antiserum (Figure 3B, Lane 2) or Type IIA procollagen antiserum (Figure 3B, Lane 3). The Type I propeptide antiserum reacts predominantly with proα1(I) and to a lesser extent with proα2(I) chain (Figure 3B, Lane 2), as shown previously (Foellmer et al., 1983). Type IIA antiserum does not show any crossreactivity with Type I procollagen at a dilution of 1:500 (Figure 3B, Lane 3). In addition, no crossreactivity with recombinant α1(I) propeptide (kindly provided by Dr. Paul Bornstein) was observed (data not shown).
Antibody Titer
We measured the antibody titer during the immunization period. The titer of antiserum was seen to rise up to 3 months. The antibody titer of serum from rabbits immunized with exon 2 propeptide reached 8000–11,000. The sensitivity was determined to be 5 pg/well (100 pg/ml).
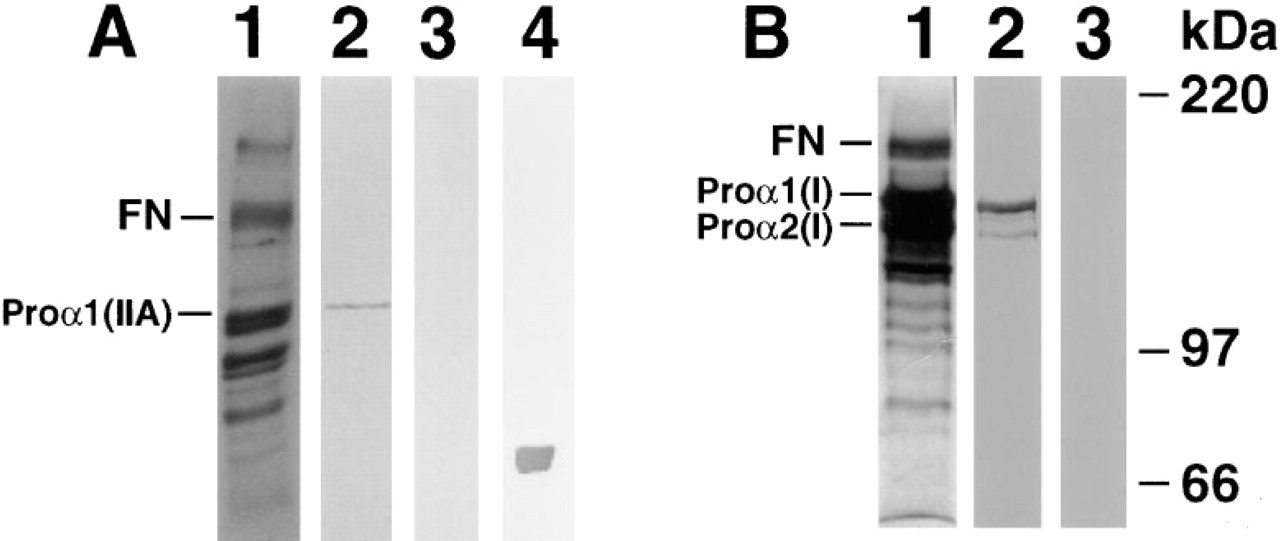
Specificity of Type IIA procollagen antiserum. [3H]-Proline labeled medium was run on SDS-PAGE and transferred onto PVDF membrane. Specific lanes of the gel were subjected to autoradiography. (
In situ hybridization to mRNA and immunohistochemistry of Day 50 chondroprogenitor tissue. (
Type IIA Procollagen mRNA and Protein Expression in Human Embryonic Tissues
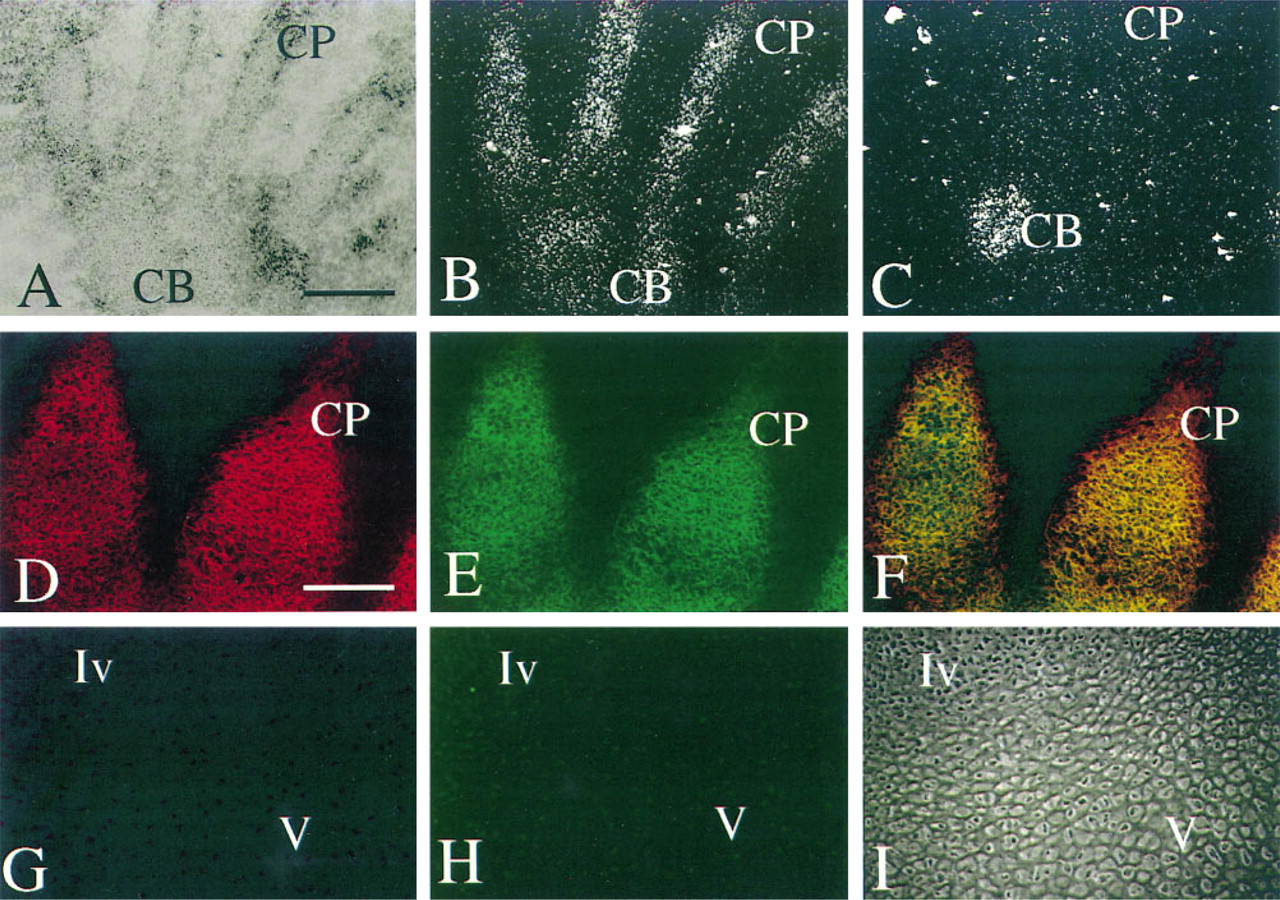
To provide direct evidence that Type IIA procollagen NH2 propeptide was present in embryonic tissues, we sought to immunolocalize Type IIA NH2-procollagen in tissues in which only Type IIA procollagen mRNA could be detected. To determine which cells were expressing Type IIA mRNA, in situ hybridization was used to detect and differentiate procollagen splice forms. To detect the tissue distribution of Type IIA procollagen, two-color indirect immunofluorescence staining was used. Figure 4A shows a brightfield image of the developing hand of a 50-day human embryo stained with cresyl violet. The mesenchymal condensations of the digital rays are outlined by darker cellular elements of the developing perichondrium. Loose connective tissue is present between and surrounding the digital rays. CP indicates chondroprogenitor cells of the digital rays, and CB indicates chondroblasts of the carpus. Figures 4B and 4C are darkfield images to silver grains resulting from in situ hybridization of probes specific for Type IIA or Type IIB procollagen mRNAs, respectively. Figure 4B, the same tissue section shown in Figure 4A, shows that Type IIA procollagen mRNA is localized to chondroprogenitor tissue of the digital ray (CP), with some detectable signal in the chondro-blasts (CB). In contrast, Figure 4C, an adjacent section, shows no detectable hybridization to Type IIB procollagen mRNA in the digital rays, whereas the chondroblasts of the carpus (CB) are positive for Type IIB procollagen mRNA expression. At this time in development, the digital rays are less mature than the carpus and within a few days will undergo endochondral bone formation. Examination of these slides at higher magnification revealed that the chondroprogenitor cells that synthesize Type IIA procollagen mRNA are fibroblastic in shape, whereas the chondroblasts synthesizing Type IIB are rounded, an observation consistent with our previous results in the developing human vertebral body and in mouse chondrogenic tissue (Sandell et al. 1991, 1994) and in chick articular cartilage development (Nalin et al. 1995). This tissue provides an opportunity to examine the expression of Type IIA procollagen protein in a chondroprogenitor tissue in which only Type IIA procollagen mRNA is expressed (as shown below).
Double immunofluorescence was performed on a tissue section adjacent to that in Figure 4A. Antisera to the collagen NH2 propeptide domain and to the triple-helical domain were used. The region indicated by CP in Figure 4A is shown at a higher magnification in Figures 4D–4F. The tissue shape was slightly different from that in Figure 4A–4C, due to the tangential orientation of the section. The Type IIA immunoreactivity was detected by reaction with a secondary antibody conjugated to the chromofluor cyanine-3 (red) and the triple-helical collagen immunoreactivity was detected by secondary antibody conjugated with FITC (green). In this chondroprogenitor tissue both domains of the Type IIA procollagen molecule were colocalized: NH2 propeptide (red fluorescence, Figure 4D) and triple-helical domain (green fluorescence, Figure 4E). Because the emission wavelengths of the two chromofluors do not overlap, the antisera can be colocalized (orange/yellow fluorescence, Figure 4F). No reactivity was observed in the surrounding mesenchymal tissue in which Type I collagen is present. In addition, no reactivity of the Type IIA procollagen antiserum was observed with tissues that contain Type III procollagen, such as blood vessels (data not shown). Negative control tissue from the same experiment is shown in Figure 4G–4I, in which preimmune sera replaced the primary antisera.
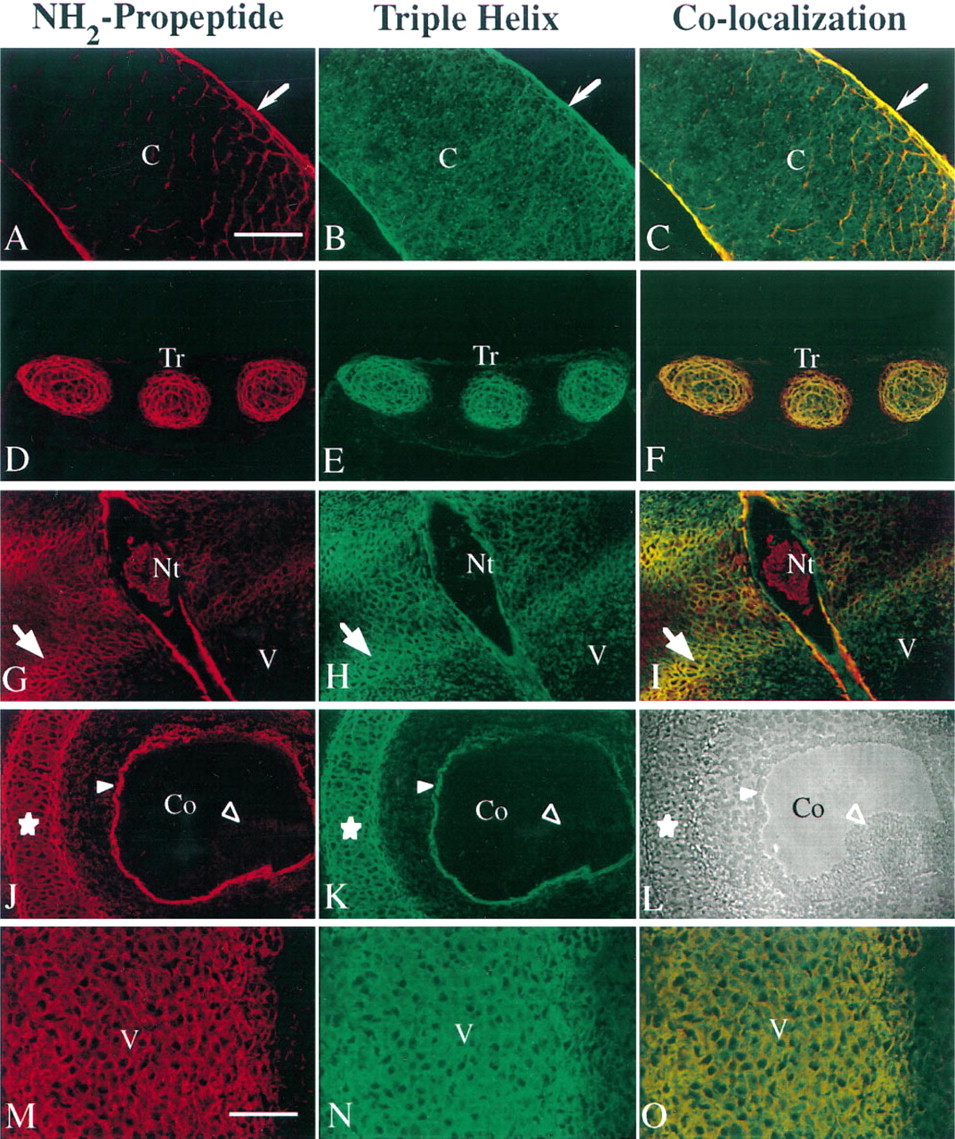
Examples of other locations of Type IIA procollagen protein in a day 53 embryo are shown in Figures 5A–5L. The tissue in Figure 5A–5C is from the proliferative zone of a developing radius. This tissue was more mature than the tissue of the digital rays shown above. This tissue is cartilaginous by histological examination and the cells synthesize the Type IIB procollagen mRNA, as determined by in situ hybridization (data not shown). The Type IIA procollagen NH2 propeptide is localized in a lace-like distribution pattern in the interterritorial matrix throughout the tissue. Note the distinct Type IIA procollagen presence in the perichondrium. The triple-helical collagen domain shows a more uniform distribution (Figure 5B). This can be seen clearly when both colors are shown in Figure 5C. The green fluorescence lies within the boundaries of the yellow fluorescence in the cartilage, and the perichondrium is yellow. Because these cells are producing Type IIB procollagen mRNA, the yellow staining represents previously synthesized Type IIA procollagen and the green staining represents more recently synthesized Type IIB collagen (without NH2 propeptide).
Tracheal rings (Figures 5D–5D), were examined as an example of a tissue that does not undergo endochondral bone formation. At this stage of development, the tracheal rings appear cartilaginous by cell morphology and staining with cresyl violet, but they have only begun to develop into mature cartilage (O'Rahilly and Muller 1992). Similar to the staining pattern of the cartilage of the radius (Figures 5A–5A), but less obvious, the NH2 propeptide co-localized with the triple-helical portion of the collagen molecule in the interterritorial matrix, whereas the triple-helical collagen alone is localized close to the cells. In both cartilages, this color distribution is caused by the cells switching their synthesis from Type IIA procollagen, laid down previously when the cells were less differentiated, to Type IIB procollagen, synthesized currently and displacing the Type IIA procollagen out into the interterritorial matrix.
Previous studies have localized Type II collagen in the notochord (Wood et al. 1991). Here we show the presence of Type IIA procollagen (Figures 5G–5G). The notochordal remnant is visible in the developing disk of the vertebral column (indicated Nt), and the chondroprogenitor domain of the developing vertebral bodies (called centra at this stage in development) is indicated with an arrow. V indicates the vertebral body. This is a complex distribution of Type II procollagen splice forms. By in situ hybridization, we have shown that Type IIA procollagen mRNA is abundant in the chondroprogenitor cells surrounding the centra and also observed in the middle of the disc (Sandell et al. 1991). In Figures 5G–5I, Type IIA procollagen can be observed in these locations. The perinotochordal sheath, an acellular tissue, also stains strongly for Type IIA procollagen. This staining pattern is identical to that shown by Wood and colleagues for triple-helical Type II collagen staining in the mouse perinotochordal sheath (Wood et al. 1991). Interestingly, the matrix surrounding the cells of the notochordal remnant, itself shown to synthesize Type IIA procollagen mRNA (Sandell et al. 1994), appears to contain primarily NH2 propeptide lacking the fibrillar domain. In the tissues examined, it was rare to find the NH2 propeptide without the triple-helical collagen domain. The complex biosynthetic pattern of the intervertebral disk is under further investigation.
An example of Type IIA procollagen localization in epithelial tissue, the developing otic vesicle and cochlea, is shown in Figures 5J–5L. Immunoreactivity for Type II collagen has been reported for many mesenchymal/epithelial tissue interfaces during chondrocranial development of the mouse (Wood et al. 1991), particularly where chondrogenesis will occur. In the developing cochlea shown here, the chondrogenic mesenchyme is undergoing differentiation into cartilage (star) to become the otic capsule and is strongly stained for Type IIA procollagen. Epithelial localization is shown in the basal surface of the organ rudiment (closed arrowhead), just inside a layer of epithelial cells. A phase-contrast picture is shown in Figure 5L so that the tissue structure can be compared to the fluorescence immunolocalization. A thickening of epithelial cells within the organ rudiment can be seen at lower right within the basal surface (Figure 5L, closed arrowhead).
Double immunofluorescence of Type IIA procollagen in Day 50 (
Figures 5M–5O demonstrate the distribution of the collagen splice forms in early cartilage of the vertebral body (Day 50). This tissue is very young and has just begun to synthesize Type IIB mRNA and aggrecan, producing a cartilaginous extracellular matrix, and stains with Alcian blue. The presence of triple-helical staining indicates all Type II collagen, whereas the Type IIA procollagen remaining in the tissue is red in Figure 5M and orange/yellow in Figure 5O. As this tissue matures and the cells synthesize more Type IIB procollagen that does not contain the NH2 propeptide, the cells will clearly demonstrate an extracellular matrix that stains green surrounding the cells (triple-helical collagen without the NH2 propeptide) and yellow/orange in the interterritorial matrix (triple-helical collagen with the NH2 propeptide).
Discussion
A great deal of information is available on the localization of mRNA encoding the Type IIA alternate splice form of Type II procollagen. However, no data have existed correlating the mRNA with the tissue distribution of the protein. For a number of years we have known that the protein can be secreted into the culture medium of chondrogenic costal cells (Sandell et al. 1991), and recently the Type IIA and Type IIB pN procollagen splice forms have been isolated and identified in bovine vitreous (Bishop et al. 1994). Here, for the first time, we produce and characterize an antiserum that reacts specifically with the cysteine-rich NH2 propeptide domain of Type IIA procollagen and localize this Type IIA procollagen NH2 propeptide in regions of embryonic tissue that contain Type IIA procollagen mRNA. Furthermore, antiserum specific for the Type II collagen triple-helical domain is co-localized with the NH2 propeptide. These results, taken together with the isolation of Type IIA pN collagen from bovine vitreous, strongly suggest that in the extracellular matrix a population of the Type IIA NH2 propeptides remain bound to the triple-helical portion of the secreted collagen molecule.
Previous studies have shown that Type IIA procollagen mRNA precedes Type IIB procollagen mRNA expression during formation of the endochondral skeleton. For example, Type IIA procollagen mRNA is present in the somites, notochord, neuroepithelia, and prechondrogenic mesenchyme of the mouse (Ng et al. 1993; Sandell et al. 1994), human (Sandell et al. 1991; Sandberg et al. 1993; Lui et al. 1995), and precartilaginous condensations and perichondrium of developing chick long bones (Nalin et al. 1995). During chondrogenesis, the splice form switches to Type IIB procollagen mRNA when mesenchymal cells differentiate into chondroblasts. Even after cells switch splice forms to Type IIB, Type IIA NH2 propeptide remains in the tissue. The localization of Type IIA procollagen in the human embryonic tissues is strikingly similar to that shown for Type II collagen in noncartilaginous tissues of mouse embryos by Wood et al. (1991). The results presented here and the previous results from in situ hybridization of mRNA also predict that the Type II collagen localized in tissues of chick and mouse embryos that do not undergo chondrogenesis, e.g., at epithelial/mesenchymal interfaces, basement membranes, and epithelia, (Hayashi et al. 1988; Fitch et al. 1989; Kosher and Solursh 1989; Wood et al. 1991) is the Type IIA procollagen splice form.
The distribution of Type IIA procollagen NH2 propeptide provides insight into the potential roles of this protein domain. We stress that these tissues are very young, approximately comparable to mouse embryonic day 13. Type IIA procollagen NH2 propeptide is found in the perichondrium surrounding proliferative cartilage as well as in the cartilage of the developing radius. Although these cells are not currently undergoing chondrogenesis, the presence of Type IIA suggests that they may at some later time. The Type IIA procollagen in the cartilage remains in the tissue from when the cells secreted it at an earlier phase of chondrogenesis. How long the Type IIA procollagen remains in the matrix is not known at this time. However, there appears to be a steady decrease from the chondroprogenitor tissue to the hypertrophic cartilage. The presence of Type IIA procollagen in the developing trachea is interesting because it indicates that, even in tissue that will not progress to endochondral bone, Type IIA NH2 propeptide is expressed and deposited in the tissue. This finding supports a role for Type IIA procollagen in early differentiation of chondrocytes rather than in a later function. The patterns of Type IIA procollagen distribution in the intervertebral disk and otic vesicle are similar to that shown by Wood et al. (1991) in the mouse, except here the two domains of collagen can be differentiated. Clearly, in the disk, the distribution of Type IIA NH2 propeptide and triple-helical domain is not always the same. This could be due to a number of factors such as: (a) cells synthesized different amounts of Type IIA and Type IIB mRNA; (b) the NH2 propeptide is not always bound to the triple-helical domain; and (c) certain epitopes may become masked in the tissue and not available for reaction with the antiserum. A detailed analysis of the cellular synthesis of mRNA splice forms and deposition of collagen will be necessary to understand the role of Type II collagens in this complex tissue. Wood et al. (1991) have previously shown that Type II collagen is present in the subepithelium of the developing cochlea of the otic vesicle. mRNA studies have shown that the neuroepithelium synthesizes Type II procollagen mRNA (Cheah et al. 1991) and that the mRNA is Type IIA (Sandell et al. 1994). This neuroepithelium is believed to play a role in the induction of chondrogenesis in mesenchymal cells.
Although the molecular form of Type IIA procollagen NH2 propeptide in vivo cannot be directly determined from these studies, various data suggest that it is present in a procollagen form. In tissue culture of embryonic ribs, pNC collagen was present, but no pN collagen was detected. This result is not unexpected because the enzymes necessary to cleave the propeptides are not accessible. Consistent with this interpretation, the fibroblast culture medium only contained pNC forms of Type I collagen. In the interesting studies of Bishop et al. (1994), Type IIA and Type IIB pN collagens were isolated from bovine vitreous. Consistent with the developmental expression of Type II collagen splice forms in chondrogenesis, in young bovine vitreous the ratio of Type IIA pN to Type IIB pN was 5:1, whereas in the older tissue the ratio was closer to 1. These investigators have recently used the Type IIA NH2 propeptide antiserum reported here and show positive reactivity with their Type IIA pN procollagen (unpublished results). Probably due to crosslinking between molecules, isolation of Type IIA procollagen from embryonic skeletal tissues has proved difficult. Lastly, in most tissues in which Type IIA is synthesized, immunolocalization of both the NH2 propeptide and triple helix is the same, inasmuch as it can be resolved by confocal microscopy. One exception is in the intervertebral disk tissue, where the notochordal remnant reacts only with the NH2 propeptide antiserum, indicating that, in some tissues, the NH2 propeptide may be bound in the matrix without the triple helix.
There are other proteins that contain the Type IIA procollagen NH2 propeptide domain, which begins to suggest diverse functional roles for this domain. Three procollagen chains, α1(I) (Chu et al. 1984), α1(III) (Liau et al. 1985), and α2(V) (Woodbury et al. 1989) possess similar cysteinerich NH2-propeptides, including the same placement of the cysteine residues as well as many others (compared by Ryan and Sandell, 1990). A similar domain is found in thrombospondins I and II (reviewed by Bornstein and Sage 1994), in which the cysteines and other amino acids are conserved, and in von Willebrand factor, in which 9 of 10 cysteine residues are conserved. The function of the collagen propeptides is predicted to be a feedback inhibitor of collagen synthesis (Fouser et al. 1991) and a regulator of fibril diameter (Fleischmajer et al. 1990). In thrombospondins, this domain is believed to be involved in regulation of angiogenesis (Good et al. 1990). It has been shown recently that two additional proteins, sog (short of gastrulation) in Drosophila (Francois et al. 1994) and chordin in Xenopus (Sasai et al. 1994), have multiple homologous domains conserving all of the cysteines and many other amino acids. The NH2 propeptide-like domains in sog and chordin function to regulate the availability of members of the TGFβ superfamily, decapentaplegic and bone morphogenetic protein 4, respectively, by direct binding (Piccolo et al. 1996), and thus participate in establishing the dorsal–ventral body pattern. Now that we have shown Type IIA NH2 propeptide in developing skeletal tissue, future studies can be directed at determining whether such a binding function is possible for Type IIA procollagen.
In summary, the presence of Type IIA procollagen in cells before terminal differentiation suggests that the unique domain in the Type IIA procollagen, the NH2 propeptide, is involved in developmental events. The finding that Type IIA procollagen is deposited into the matrix of human embryos and retains the unique NH2 propeptide domain, taken together with the recent discovery that a homologue of this domain is capable of binding to a bone morphogenetic protein, lends support to the hypothesis that Type II collagen, i.e., Type IIA procollagen, functions in the early embryo and, potentially, is the portion of the Type II collagen molecule that plays a role in the induction of chondrogenesis in receptive cells, as suggested previously (Thorogood et al. 1986; Wood et al. 1991).
Footnotes
Acknowledgements
Supported by funds from the National Institute of Arthritis, Skin and Musculoskeletal Disease (RO136994), the Department of Veterans Affairs, and by a Bristol-Myers Zimmer Center of Excellence Grant to the Department of Orthopaedics, University of Washington School of Medicine.
We thank Michael Cremer for providing antibodies, Dr Paul Goodwin, Fred Hutchinson Cancer Research Institute, for use of the confocal microscope, Drs David Eyre and James Wu, University of Washington, for protein sequence determination, and Margo Weiss for assistance in preparing the manuscript.