
Abstract
Merkel cells represent a population of epithelial cells in the skin and oral mucosa. Although Merkel cells are reliably distinguishable from other epithelial cells at the ultrastructural level, these cells are usually not discernible by standard light microscopy and need special techniques for their identification. Villin is an actin-crosslinking protein that is associated with the actin filament cores of brush border microvilli. In this study we show that an antibody against villin is an excellent marker of Merkel cells and their microvilli even at the light microscopic level. The surrounding keratinocytes and subepithelial connective tissue cells do not show any significant affinity for the antibody against villin. Confocal laser micrographs reconstructed from serial images 0.5 μm thick of Merkel cells that were immunostained with villin clearly reveal the three-dimensional morphology of Merkel cells and their microvilli. The presence of villin in Merkel cell microvilli lends support to the idea that these cells might have a mechanoreceptor function.
The function(s) of Merkel cells is(are) still controversial. The widely held belief that the Merkel cell is a mechanoreceptor cell is probably based on the presence of microvilli at the cell surface and the accumulation of dense-cored granules in the cytoplasm facing the nerve terminals. These microvilli may act as “feelers” sensitive to mechanical distortion, which may cause the release of transmitters from the Merkel cell. The present study was undertaken to identify Merkel cells by the use of an antibody specific for villin, the actin-crosslinking protein, which is associated with the actin filament cores of microvilli. In the present report, we show that the antibody against villin is an excellent marker of Merkel cells and their characteristic microvilli for both light and confocal laser microscopy.
Materials and Methods
Rabbits were sacrificed by IV injection of an overdose of sodium pentobarbital. The hard palate membrane was removed and immediately fixed in 4% paraformaldehyde in 0.1 M phosphate buffer for 4 hr. After rinsing overnight with the same buffer containing 30% sucrose, the tissues were quickly frozen to the temperature of a dry ice-isopentane mixture and cut at 8 or 20 μm thickness in a freezing microtome. The sections were then either processed for diaminobenzidine (DAB) immunohistochemistry (8-μm sections) or immunofluorescence histochemistry (20-μm sections). DAB immunohistochemistry was done with the avidin-biotin-peroxidase complex (ABC) method (Hsu et al. 1981) employing the Vectorstain Elite ABC kit (Vector; Burlingame, CA). The sections were washed in PBS, incubated in normal goat serum for 30 min, and then incubated for 6 hr at 4C in a solution containing a mouse monoclonal antibody against human villin (Transformation Research; Framingham, MA) at a dilution of 1:100 in PBS. Immunoblotting against total cell proteins of human colon carcinoma resolved in SDS gels revealed that this antibody was highly specific for villin (Transformation Research). After several rinses in PBS, sections were treated with the biotinylated secondary antibody for 2 hr, rinsed several times, and visualized with DAB. For fluorescence immunohistochemistry, fluorescein isothiocyanate (FITC)-conjugated goat anti-mouse IgG1 (Southern Biotechnology Associates; Birmingham, AL) was used as a secondary antibody for 1 hr at room temperature (RT) and examined in a confocal laser scanning microscope. Negative controls to immunoperoxidase and immunofluorescence staining were performed by replacing the primary antibody with PBS or a nonimmune serum.
For double immunolabeling, the sections were incubated in an equal part mixture of a 1:100 dilution of monoclonal antibody against villin and a 1:20 dilution of monoclonal antibody against CK 20 (Progen Biotechnik; Heidelberg, Germany). Reaction products were sequentially labeled with FITC-conjugated goat anti-mouse IgG1 (Southern Biotechnology Associates) and tetrarhodamine isothiocyanate (TRITC)-conjugated goat anti-mouse IgG2a (Southern Biotechnology Associates) by incubation for 90 min at RT for each labeled antibody, and were observed under a confocal laser scanning microscope.
For electron microscopic immunohistochemistry, the preembedding technique was employed on cryostat sections of rabbit hard palate that was fixed in 4% paraformaldehyde. Sections were incubated with the villin antibody and then treated with the ABC method as described above. The sections were postfixed for 2 hr in 2% osmium tetroxide, dehydrated in ethanol, and embedded in Epon 812. Ultrathin sections were obtained from selected areas containing immunopositive Merkel cells, as assessed by light microscopic evaluation of semithin sections, and were observed without further staining. A paired adjacent ultrathin section was stained routinely with uranyl acetate followed by lead. Both types of sections were examined in a JEM-1200 EX electron microscope.
Results
Merkel cells were clustered in the basal cell layer at the base of epithelial rete pegs of every palate raphe. Ultrastructurally, Merkel cells were characterized by the following features: (a) many dense-cored granules 80–120 nm in diameter; (b) Golgi complexes in the cytoplasm opposite the major aggregation of densecored granules; (c) many microvilli extending randomly from the cell surface; (d) desmosomal attachments to neighboring epithelial cells; and (e) frequent association with nerve terminals (Figure 1A). The core of each microvillus contained a bundle of parallel actin filaments extending from its tip into the cell body (Figures 1B and 1C).
In cryosections of hard palate, the monoclonal antibody specific for villin gave intense labeling of Merkel cells scattered in the basal layer of the epithelium (Figures 2A and 2B). In contrast to Merkel cells, the surrounding keratinocytes and subepithelial connective tissue cells did not show any significant affinity for the antibody against villin. The villin antibody displayed a strong affinity both for microvilli and for a narrow cytoplasmic band beneath the Merkel cell membrane. In addition, diffuse or reticular staining of variable density was detected in the cell body. In favorable planes of tangential sections, the cross-sectioned microvilli appeared as densely labeled dots. Confocal laser microscopy of Merkel cells labeled for villin showed that the microvillar processes were more brightly fluorescent than the cell bodies (Figure 2C).
A particular advantage of using confocal microscopy reconstructions from 20 serial images at steps of 0.5 μm was the identification of the three-dimensional extent of characteristic microvilli of Merkel cells (Figures 2D and 2E). There were at least 50 microvilli in each Merkel cell. These microvilli measured about 0.3–0.5 μm in width and 2–4 μm in length, and usually had a straight configuration. However, bent or bifurcated microvilli also were occasionally observed. In double-labeling experiments with antibodies against CK20 (Figure 2F) and villin (Figure 2G), the CK20-positive Merkel cell population also could be labeled specifically with villin.
Electron microscopic immunostaining for villin clearly revealed that immunoreactions were exclusively confined to the Merkel cells (Figure 3A). Immunoperoxidase label was found mainly in association with microvilli of Merkel cells. When a paired adjacent ultrathin section was stained with uranyl acetate followed by lead, dense-cored granules and microvilli characteristics of Merkel cells could be recognized in the same cell (Figure 3B).
Discussion
This study shows that a specific marker, villin, allows reliable identification of Merkel cells and their characteristic microvilli by immunohistochemistry. Villin, a calcium-regulated actin binding protein associated with the axial microfilament bundle, was originally isolated from intestinal microvilli (Bretscher and Weber 1979; Matsudaira and Burgess 1979). Subsequently, villin has been reported to occur in a limited number of epithelial cell types of the digestive, respiratory, and urogenital systems (Robine et al. 1985; Rodman et al. 1986; Horvat et al. 1990; Höfer and Drenckhahn 1992,1996; Kasper et al. 1994).
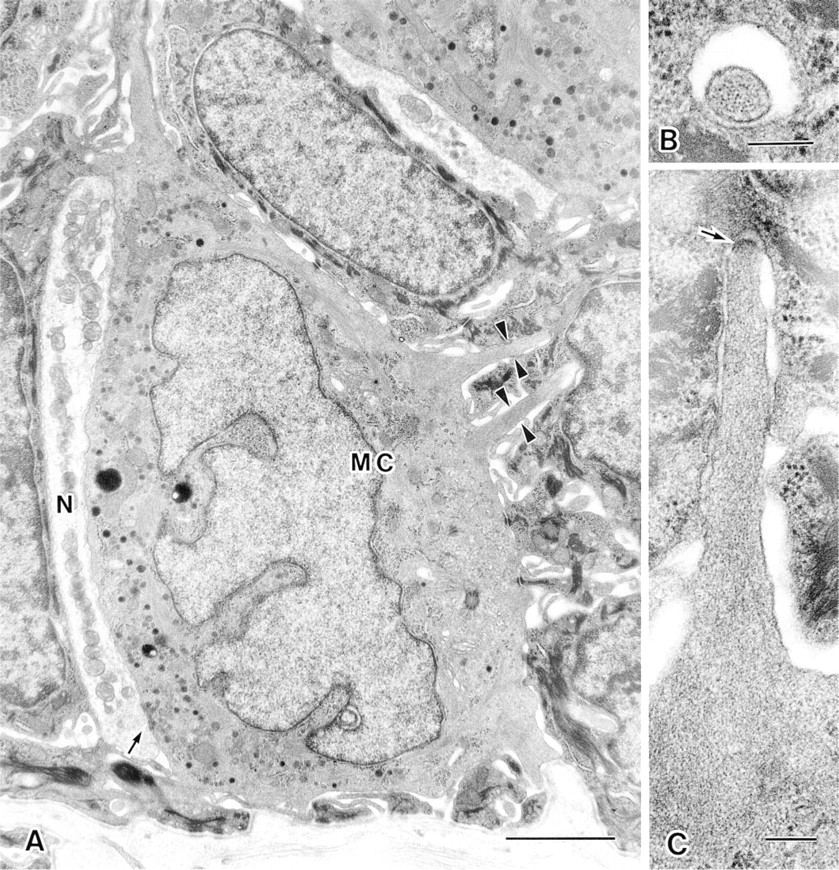
Electron micrograph of a Merkel cell (MC) and associated nerve terminal (N) in the rabbit hard palate. The Merkel cell has a concentration of dense-cored granules in the cytoplasm facing the nerve terminal and has microvilli (arrowheads) extending from the cell surface. The arrow points to a synapse-like junction between the Merkel cell and nerve terminal (
It is known that villin plays an important role in the organization of the brush border cytoskeleton (Mooseker et al. 1980; Drenckhahn and Dermietzel 1988; Friederich et al. 1989). The expression of villin is usually confined to the apical tuft of microvilli and their rootlets. Höfer and Drenckhahn (1992,1996) first showed that, in addition to the apical brush border, the basolateral microvilli of brush cells in the alimentary and respiratory system also were immunoreactive for antibodies against villin and fimbrin. According to Luciano and Reale (1979,1990), major characteristics of lateral microvilli of brush cells are: (a) the axial bundle of microfilaments, (b) their stiffness and their course penetrating more than 3 μm into corresponding invaginations of the flanking epithelial cells, and (c) different angles at which the microvilli arise from the lateral cell surface. Both apical and lateral microvilli appear to be supported internally by actin filaments that are bundled by villin and fimbrin (Höfer and Drenckhahn 1992). Luciano and Reale (1990) noted that the lateral microvilli of brush cells closely resemble those of Merkel cells, and suggested that brush cells could be involved not only in chemoreception but also in mechanoreception.
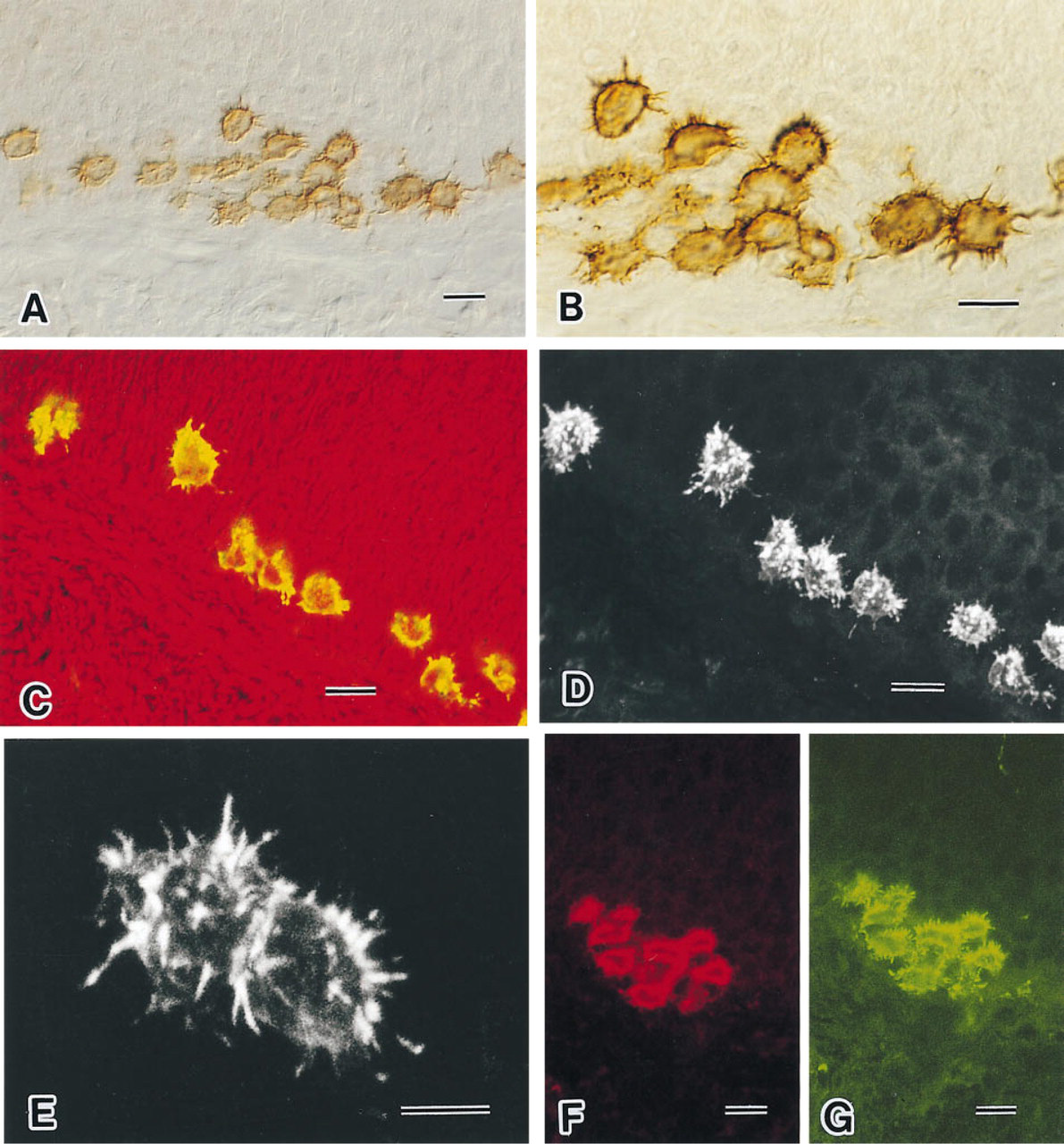
Light micrographs of palatal Merkel cells visualized by DAB immunohistochemistry using an antibody to villin (
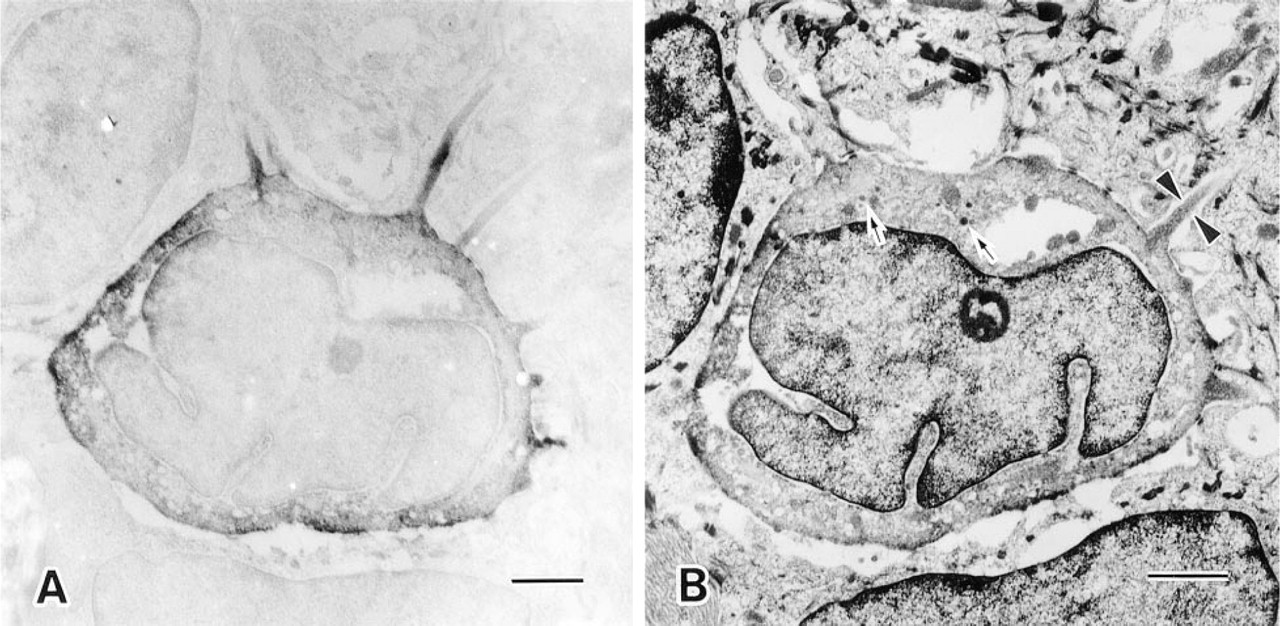
Immunoelectron micrograph showing a Merkel cell with immunoreactive villin (
The function of Merkel cells is still under debate. Since Merkel (1875) first discovered these cells in the epidermis of pig snout skin and called them “Tastzellen,” it has been widely believed that Merkel cells are involved in mechanoreception. The presence of many feeler-like microvilli at the cell surface favored this concept. In the present study, more than 50 microvilli could be counted in each Merkel cell. This appeared to be more numerous than was anticipated on the basis of previous electron microscopic observations. Ultrastructurally, these microvilli appear to be rigid. Actin filaments crosslinked by villin might be responsible for the rigidity of microvilli, a condition that may amplify and transmit movement of surrounding keratinocytes to the Merkel cell body. Such deformation might cause the generation of a receptor potential, which may be transmitted to the apposed nerve terminals through the synaptic release of chemical transmitter(s) from the dense-cored granules (for reviews see Hartschuh et al. 1986; Ogawa 1996). Immunohistochemistry has provided further evidence for the association of several peptides and amines, including neurotransmitter candidates (Hartschuh and Weihe 1988; Hartschuh et al. 1989), with Merkel cells.
However, controversy continued as to whether or not the Merkel cell is a mechanoreceptor (Zelená 1994). Scott et al. (1981) hypothesized that Merkel cells are the target for mechanosensory axons during development. It is known that not all Merkel cells are associated with a nerve fiber (Garant et al. 1980). Furthermore, there is evidence that Merkel cells appear in the epidermis before the arrival of somatosensory nerve fibers (Moll et al. 1984; Dell and Munger 1986; Pasche et al. 1990; Narisawa and Hashimoto 1991). It has been suggested that Merkel cells release a nerve growth factor-like attractant substance for the ingrowing nerve fibers, or provide a stop signal to them. They plausibly could be involved in providing positional information to the nerves that contact them (Diamond et al. 1988; Mills and Diamond 1995).
If the Merkel cell is not a mechanoreceptor, then the significance of microvilli at the cell surface remains unexplained. The presence of villin in Merkel cell microvilli lends support to the notion that these cells engage in mechanoreception.
Footnotes
Acknowledgments
We thank Dr B. Tandler for critical reading of the manuscript and M. Ueda and T. Tanaka for assistance with the CLMS.