
Abstract
Rat ameloblastin is a recently cloned tooth-specific enamel matrix protein containing 422 amino acid residues. We investigated the expression of this protein during the matrix formation stage of the rat incisor immunohistochemically and immunochemically, using anti-synthetic peptide antibodies that recognize residues 27–47 (Nt), 98–107 (M-1), 224–232 (M-2), 386–399 (M-3), and 406–419 (Ct) of ameloblastin. Immunohistochemical preparations using antibodies Nt and M-1 stained the Golgi apparatus and secretory granules of the secretory ameloblast and the entire thickness of the enamel matrix. Only M-1 intensely stained the peripheral region of the enamel rods. Immunostained protein bands were observed near 65, 55, and below 22 kD. Immunohistochemical preparations using antibodies M-2 and Ct stained the Golgi apparatus and secretory granules of the ameloblast and the immature enamel adjacent to the secretion sites, but not deeper enamel layers. Immunostaining using M-2 and Ct revealed protein bands near 65 and 40–56 kD, and 65, 55, 48, 36, and 25 kD, respectively. M-3 stained the cis side of the Golgi apparatus but not the enamel matrix. This antibody recognized a protein band near 55 kD, but none larger. After brefeldin A treatment, immunoreaction of the 55-kD protein band intensified, and dilated cisternae of rER of the secretory ameloblast contained immunoreactive material irrespective of the antibodies used. These data indicate that ameloblastin is synthesized as a 55-kD core protein and then is post-translationally modified with O-linked oligosaccharides to become the 65-kD secretory form. Initial cleavages of the 65-kD protein generate N-terminal polypeptides, some of which concentrate in the prism sheath, and C-terminal polypeptides, which are rapidly degraded and lost from the enamel matrix soon after secretion.
Keywords
D
Recent investigations of rat cDNA libraries revealed full-length sequences of tooth-specific proteins homologous to porcine sheath protein, named ameloblastin (Krebsbach et al. 1996) and amelin (Cerny et al. 1996). Ameloblastin consists of 422 amino-acid residues including a putative signal peptide that may be cleaved after residue 26 (Krebsbach et al. 1996). Amelin 1 was identical to ameloblastin, with the exception that it lacked a 15 amino-acid segment corresponding to residues 104–118 of ameloblastin (Cerny et al. 1996).
We also characterized two novel clones from a porcine enamel organ-specific cDNA library encoding sheath proteins, and proposed that the porcine sheath proteins and their proteolytic cleavage products be designated sheathlin (Hu et al. 1997). Ameloblastin and the larger sheathlin isoform share 79% identity in their nucleotide alignment of coding regions and 66% identity in their amino acid sequences. The short form of sheathlin lacked a 15 amino-acid segment corresponding to residues 99–113 of long form. The difference between the two isoforms was presumably due to alternative splicing of the sheathlin primary RNA transcript(s) (Hu et al. 1997). On the basis of sequence alignments, the 15 amino-acid segment that distinguishes the long and short forms of sheathlin corresponds exactly in position to the 15 amino-acid segment present in ameloblastin but lacking in amelin 1.
Ameloblastin and amelin contain two consensus sequences for peptide domains, DGEA and VTKG, interacting with cell surface proteins (Cerny et al. 1996), whereas sheathlin contains no such sequence (Hu et al. 1997). Amelin mRNA transcripts can be detected not only in the secretory ameloblast but also in the maturation ameloblast, reduced enamel epithelium, and cells of the Hertwig's epithelial root sheath (Fong et al. 1996). Recent immunohistochemical study using antibodies against recombinant ameloblastin revealed immunolocalization of ameloblastin in the presecretory, secretory, and maturation ameloblast (Lee et al. 1996). Our investigation in rat has shown that immunoreactivity of ameloblastin was detected within the cells of the epithelial root sheath adjacent to the peripheral surface of newly deposited dentin of the molar root end and those adjacent to the lingual side of incisors (unpublished data). On the other hand, immunolocalization of sheathlin has been found only in the ameloblast at the stage of late differentiation, matrix formation, and transition (Uchida et al. 1995; Murakami et al., in press). These data suggested that, despite the homology, ameloblastin/amelin may serve functions different from those of sheathlin.
We have already demonstrated immunolocalization of sheathlin in the porcine tooth germ using region-specific antibodies. N-terminal-specific anti-sheathlin antibodies stained the prism sheath throughout the full thickness of immature enamel at the stages of matrix formation, transition, and early maturation (Uchida et al. 1995). On the other hand, the C-terminal region of sheathlin in porcine immature enamel was restricted to the surface layer of immature enamel at the stage of matrix formation and transition. The middle to inner enamel layers showed almost no immunoreactivity (Murakami et al., in press). Whereas the amino acid sequence of ameloblastin is similar to that of sheathlin, little is known about the localization of ameloblastin in the immature enamel. Krebsbach and co-workers (1996) have shown that ameloblastin is contained in the secretory ameloblast, especially in the Tomes' process, and in the enamel matrix, using an antibody raised against recombinant ameloblastin. Most recently, colloidal gold immunolabeling using the same antibody further demonstrated that ameloblastin is present in the Golgi apparatus, secretory granules, and throughout immature enamel (Lee et al. 1996) However, precise localization of nascent ameloblastin and its cleaved polypeptides was not shown.
In this study we investigated the synthesis, secretion, degradation, and fate of ameloblastin at the matrix formation stage of the rat incisor by immunochemical and immunohistochemical methods, using affinity-purified antibodies raised against synthetic peptides.
Materials and Methods
Antibodies
Five antibodies were used in this study. Because the amino acid sequence of ameloblastin is similar to that of sheathlin (Hu et al. 1997), two antibodies generated against the sheathlin sequence were used in this study. The production and characterization of these antibodies, termed antibodies Nt and M-1, have been reported previously (Uchida et al. 1995; Hu et al. 1997). The amino acid alignment of sheathlin vs ameloblastin shows that antibodies Nt and M-1 should recognize ameloblastin residues 27–47 and 98–107, respectively. For generating other antibodies, synthetic peptides including the amino acid sequences MSYGANQLN (residues 224–232), DGEATMDITMSPDTQ (residues 386–399), and NKVHQPQVHNAWRF (residues 406–419) of ameloblastin were assembled with a PSSM-8 automatic peptide synthesizer (Shimazu; Kyoto, Japan) using a standard solid-phase method. Cysteine residues were introduced at the C-terminus of these peptides for crosslinking with amino groups present in the carrier protein. Fmoc-amino acids and amino acid-attached resin were obtained from Watanabe Chemical Industries (Hiroshima, Japan) and Shimazu, respectively. After purification using reversed-phase HPLC equipped with a column of TSK-gel ODS-80TS (TOSOH; Tokyo, Japan), the synthetic peptides were coupled to either bovine serum albumin (BSA; Sigma, St Louis, MO) or ovalbumin (Sigma) using 3-maleimidobenzoic acid N-hydroxy-succinimide ester (MBS; Sigma) and were used to immunize rabbits as reported previously (Uchida et al. 1991b, 1995). Specific antibodies were purified on affinity columns of the synthetic peptides coupled to amino-cellulofine (CHISO; Tokyo, Japan) using MBS as described previously (Uchida et al. 1991b). They were termed antibodies M-2 (residues 224–232), M-3 (residues 386–399), and Ct (residues 406–419).
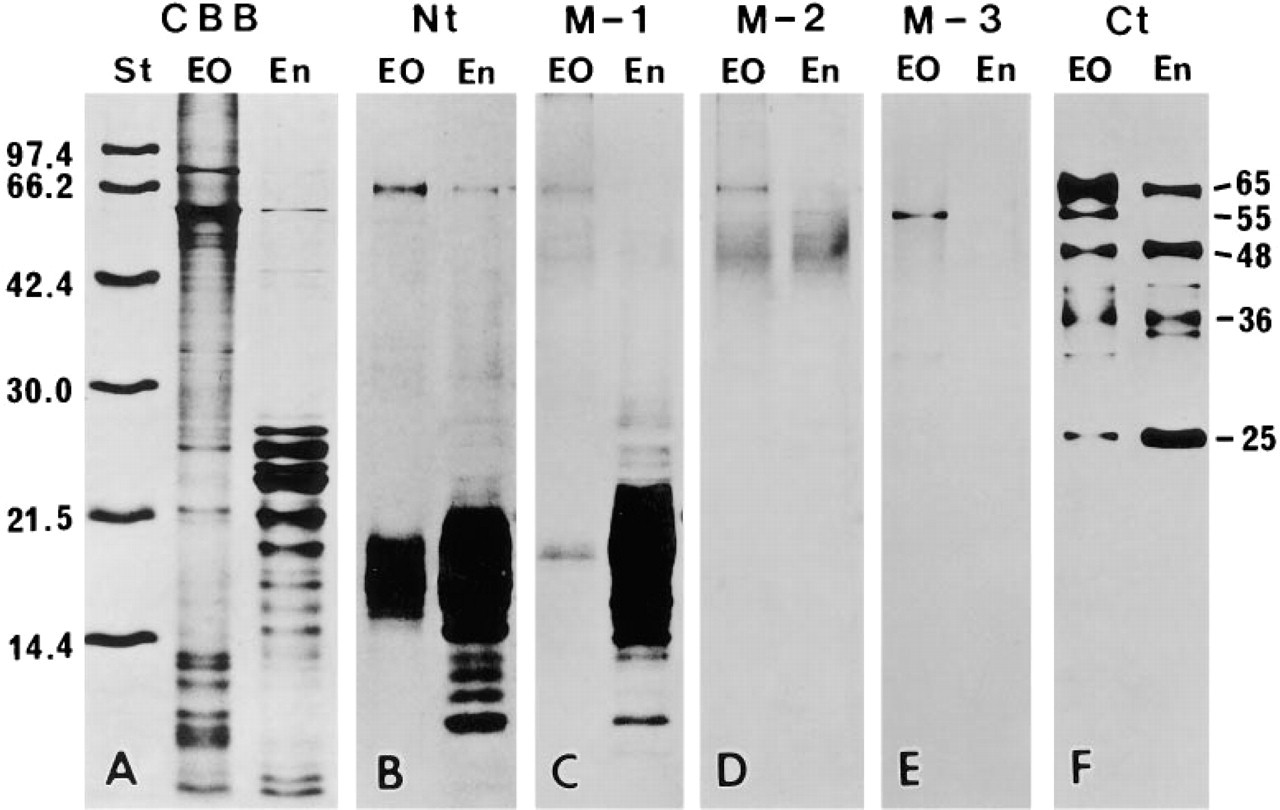
Detection of proteins extracted from enamel organ (EO) and immature enamel (En) by SDS-PAGE (A) and immunochemistry (
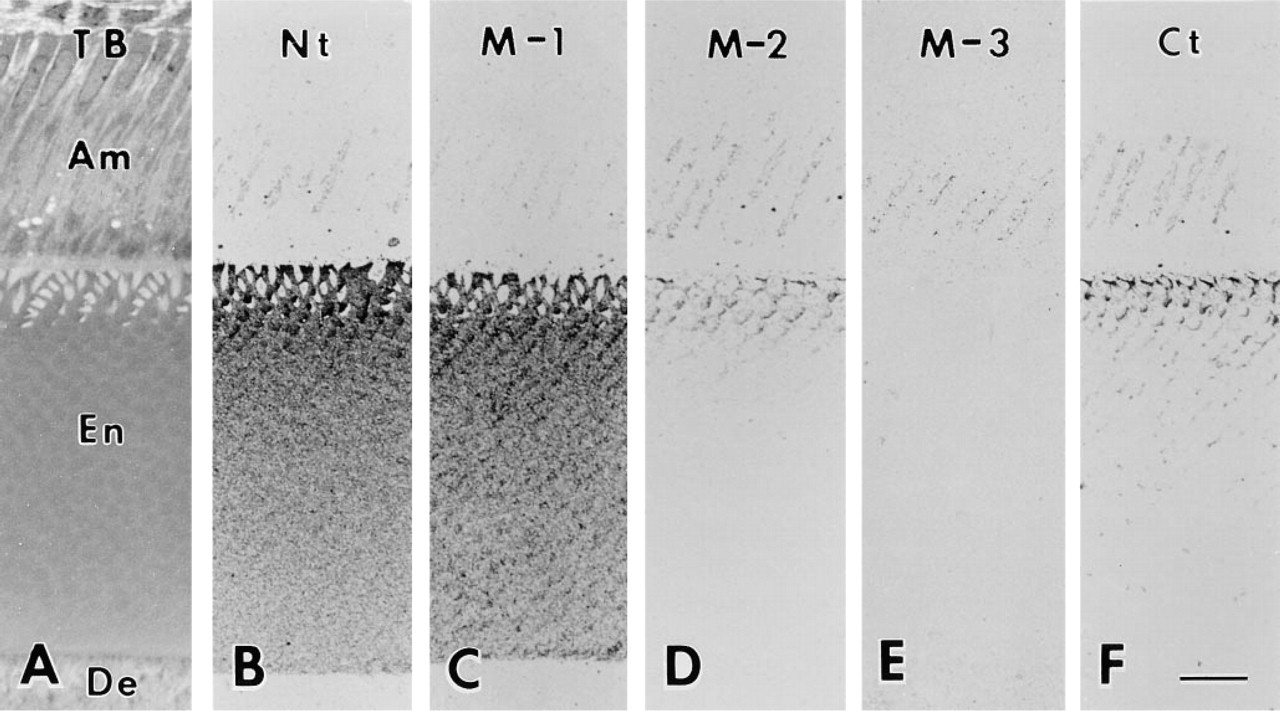
Light micrographs of toluidine blue-stained (
Because antibodies Nt, M-1 and M-2 were raised against BSA–peptide conjugates, they sometimes reacted faintly with rat serum albumin, which appeared as a protein band near 57 kD in immunochemical analyses under nonreducing condition. Therefore, these antibodies were preabsorbed with an excess amount of BSA before use. Antibodies M-3 and Ct did not react with rat serum albumin because they were generated against ovalbumin-peptide conjugates.
Animals
Male Wistar rats weighting 100–150 g, purchased from Charles River Japan (Osaka, Japan), were used throughout this study. Experimental animals were anesthetized with sodium pentobarbital (Abbott; North Chicago, Illinois; 5 mg/100 g bw) and injected with brefeldin A (Sigma; 0.2 mg/100 g bw) dissolved in 0.1–0.15 ml of distilled water containing 10% methanol and 40% propylene glycol via the external jugular vein 1 hr before sacrifice. Control animals were injected with the same amount of this solution without brefeldin A. Brefeldin A is a potent inhibitor of protein transport from the rough endoplasmic reticulum (rER) to the Golgi apparatus (Fujiwara et al. 1988), an effect that has been confirmed for secretory ameloblasts both in vitro (Fabian 1996) and in vivo (Hashimoto and Nanci 1996; Smith and Nanci 1996).
Immunochemical Analysis
For immunochemical analyses, four untreated, four experimental (brefeldin A-injected), and two control rats were used. The animals were anesthetized with sodium pentobarbital and decapitated. The upper and lower incisors were removed from the alveolar bone and the enamel organ of the matrix formation stage was dissected with a fine forceps. The immature enamel surface was then gently wiped with a small cotton ball to remove remnants of secretory ameloblasts, and the immature enamel was dissected with scalpels. The samples obtained from each rat were further processed individually. Total protein was extracted with 0.5 M acetic acid and lyophilized. Samples were electrophoresed on 15 % polyacrylamide gel containing 0.1% sodium dodecyl sulfate (SDS-PAGE) under nonreducing conditions and then transblotted onto nitrocellulose membranes as described previously (Uchida et al. 1991a). The membranes were immunostained with an avidin–biotin–peroxidase kit (Vectastain ABC Elite Kit; Vector Labs, Burlingame, CA) using 3,3-diaminobenzidine and nickel ammonium sulfate as chromogens. The concentration of primary antibodies was either 5 or 10 μg/ml.
Immunohistochemical Analysis
For immunohistochemical analyses, five untreated, four experimental (brefeldin A-injected), and two control rats were used. The animals were anesthetized with sodium pentobarbital and perfusion-fixed with 4% paraformaldehyde/1% glutaraldehyde in 0.067 M phosphate buffer, pH 7.4. The lower incisors were removed from the alveolar bone and decalcified with 10 % EDTA for about 1 week at 4C. Without postfixation in osmium tetroxide, they were dehydrated with graded N,N-dimethylformamide at progressively lower temperatures from 4 to −20C and embedded in a glycol meth-acrylate mixture at −20C (Uchida 1985).

Light micrographs of secretory ameloblast (Am), immature enamel (En), and dentin (De) at the stage of outer enamel secretion from longitudinal sections of a rat incisor. (
The region of the inner and outer enamel secretion (War-shawsky and Smith 1974) was selected and cut in various planes of section. Thick sections (1 μ) were mounted on glass slides and stained with toluidine blue for histological observation. Ultrathin sections (about 80 nm) were mounted on glass slides for light microscopic immunohistochemistry or on nickel grids for electron microscopic immunohistochemistry. They were processed for immunogold–silver staining as described previously (Uchida 1991a, b), with slight modifications. The sections were first immersed for 5 min in a blocking solution consisting of 0.2% casein, 0.2% ovalbumin, 0.1% Tween-20, and 0.1% Triton X-100 dissolved in Tris-buffered saline (TBS; 20 mM Tris-HCl, 150 mM NaCl, pH 8.2). They were then incubated for 15 min with antibodies diluted with blocking solution. The concentration of antibody was 5–10 μg/ml for both light and electron microscopy. After rinsing excess antibodies with TBS, the sections were immersed in colloidal gold particles (average diameter 3 nm) conjugated with goat anti-rabbit IgG antibodies diluted with blocking solution for 15 min. They were washed with TBS followed by distilled water and air-dried. Silver intensification was carried out as described previously (Uchida et al. 1991b). The intensification time was about 30 min for light microscopy and 6 min for electron microscopy. The sections were then immersed in a diluted photographic fixative, rinsed in distilled water, air-dried, and mounted with Entellan for light microscopy. For electron microscopy, the sections were stained with uranyl acetate followed by lead citrate and examined with a Hitachi H-300 electron microscopy (Nissei Sangyou; Tokyo, Japan). All the processes described above were carried out at room temperature.
Immunohistochemical specificity was checked by the absorption test as described previously (Uchida et al. 1995). Diluted antibody was preincubated overnight with an excess of purified synthetic peptide and then used to immunostain tissue sections.
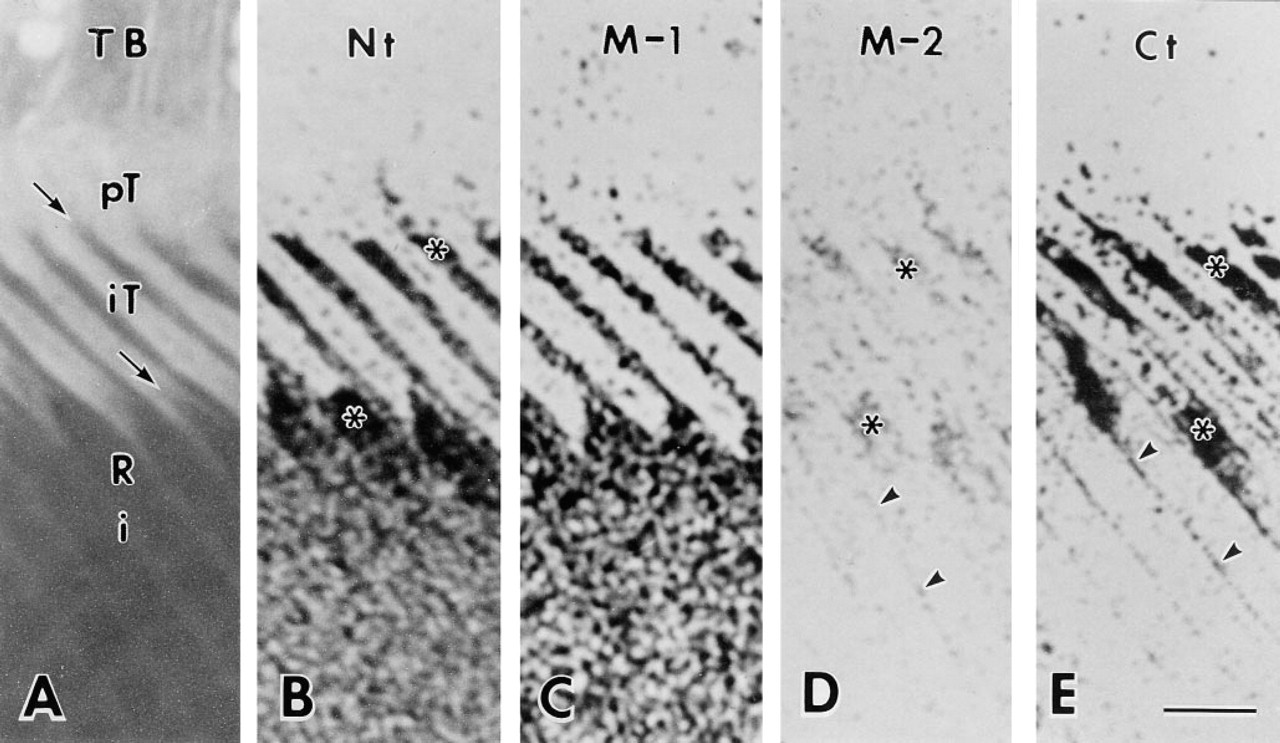
Light micrographs of toluidine blue-stained (
Results
Immunochemical Analysis
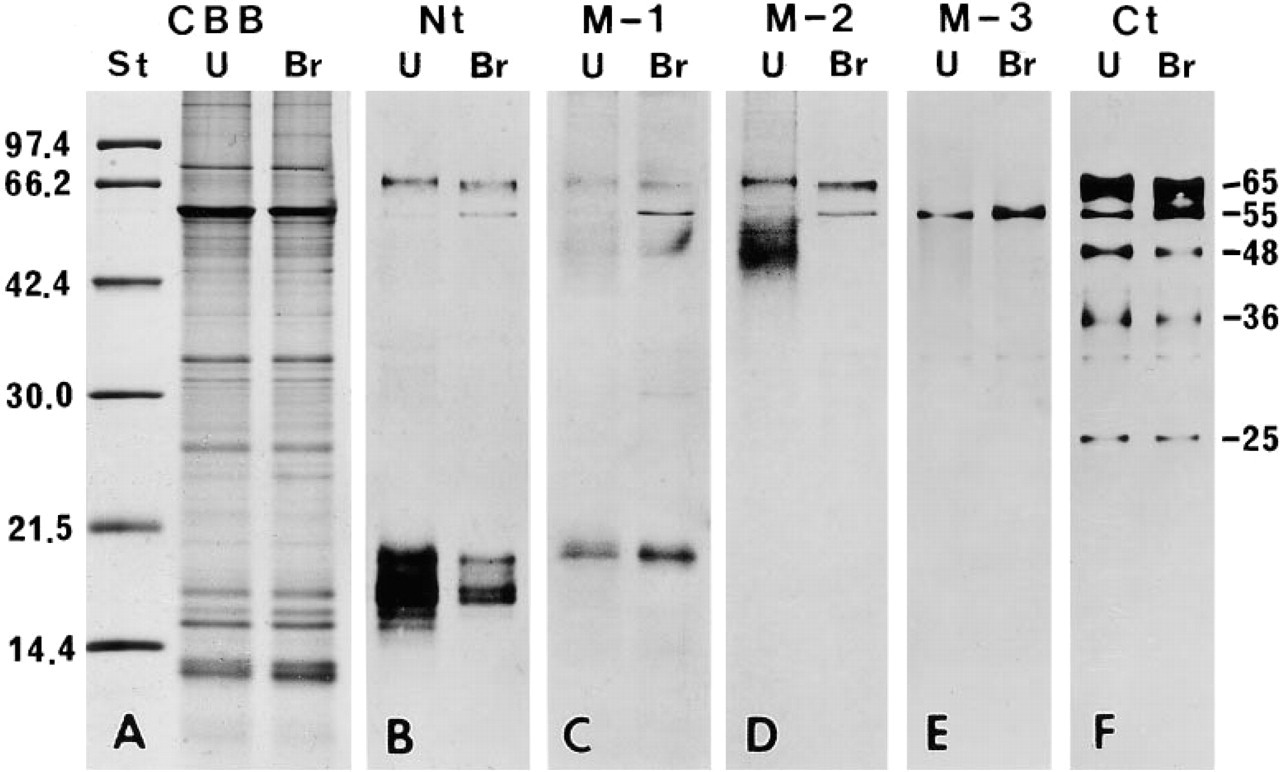
Figure 1 shows the results of SDS-PAGE and Western blot analysis. There was almost no variation among the four animals used in this study. In the enamel organ sample, antibody Nt reacted with protein bands having molecular weights of 65 kD and 15 to 18 kD, and a faint band near 55 kD. In the enamel matrix sample, the 65-kD protein band stained weakly, and proteins ranging from 10 to 22 kD stained intensely. No reaction was observed near 55 kD (Figure 1B). Antibody M-1 stained many protein bands ranging from 10 kD to 23 kD in the enamel matrix and a single protein band near 18 kD in the enamel organ. The 65-kD and 55-kD protein bands were scarcely discernible in the enamel organ sample and not at all in the enamel matrix sample (Figure 1C). Several faintly stained protein bands ranging from 23 to 50 kD appeared after staining with antibodies Nt and M-1, especially in the enamel matrix sample (Figures 1B and 1C). Electrophoresis using a freshly prepared sample showed that almost no band was found between 23–50 kD. Therefore, the proteins ranging from 23 to 50 kD stained by the antibodies Nt and M-1 were probably aggregates of lower molecular weight proteins. Antibody M-2 faintly reacted with a protein band near 65 kD and a diffuse band around 40–56 kD of the enamel organ. In the enamel matrix sample the 65-kD protein band was not detected, but a diffuse band around 40–56 kD was observed (Figure 1D). Antibody M-3 stained a protein band near 55 kD distinctly and 33 kD faintly from the enamel organ. No staining band was found in the enamel matrix (Figure 1E). Antibody Ct stained protein bands near 65 kD, 55 kD, 48 kD, 41 kD, 36 kD, 35 kD, 33 kD, and 25 kD of the enamel organ. In the enamel matrix, a similar staining pattern was obtained. However, no band was detected near 55 kD and 33 kD and the immunoreaction of the 65-kD protein band was reduced, whereas that of the 25-kD protein was intensified (Figure 1E). The 57-kD protein band, corresponding to rat serum albumin under non-reducing conditions, was distinct in the enamel organ and the enamel sample after staining with CBB (Figure 1A), but was not reactive with all the antibodies used (Figure 1B-1F). Only antibody M-2 faintly stained a protein band near 56 kD. However, comparing the staining patterns after CBB and M-2, it was clear that the 56-kD protein band stained by the antibody M-2 did not correspond to rat serum albumin (Figures 1A and 1D).
Light Microscopic Immunocytochemistry
The five antibodies displayed characteristic staining patterns. There was no variation in the staining pattern among animals. Antibody Nt intensely stained the Golgi apparatus of the ameloblast and the enamel matrix. Over the enamel matrix, this antibody produced a decreasing gradient from the surface towards the dentino–enamel junction (Figure 2B). Antibody M-1 weakly stained the Golgi apparatus. Staining over the enamel matrix was intense and was similar to that produced by antibody Nt. However, in the middle- to innerlayer enamel, immunoreaction concentrated over the peripheral region of the enamel rod (Figure 2C). This staining pattern was clearer at the stage of outer enamel secretion (Figure 3B). Antibody M-2 stained the Golgi apparatus and the superficial layer of enamel matrix. The middle and inner layers were almost devoid of immunoreactivity (Figure 2D). Antibody M-3 stained the Golgi apparatus, but no reaction was found over the enamel matrix (Figure 2E). Antibody Ct intensely stained the Golgi apparatus of the ameloblast and immature enamel near the Tomes' processes. At the middle layer of immature enamel, the periphery of the enamel rod was also stained (Figure 2F). In addition to the Golgi apparatus and enamel matrix, antibodies Nt, M-1, M-2, and Ct occasionally stained small spherical bodies in the ameloblast. All the antibodies used stained neither dentin nor odontoblasts (Figures 2B–2F).
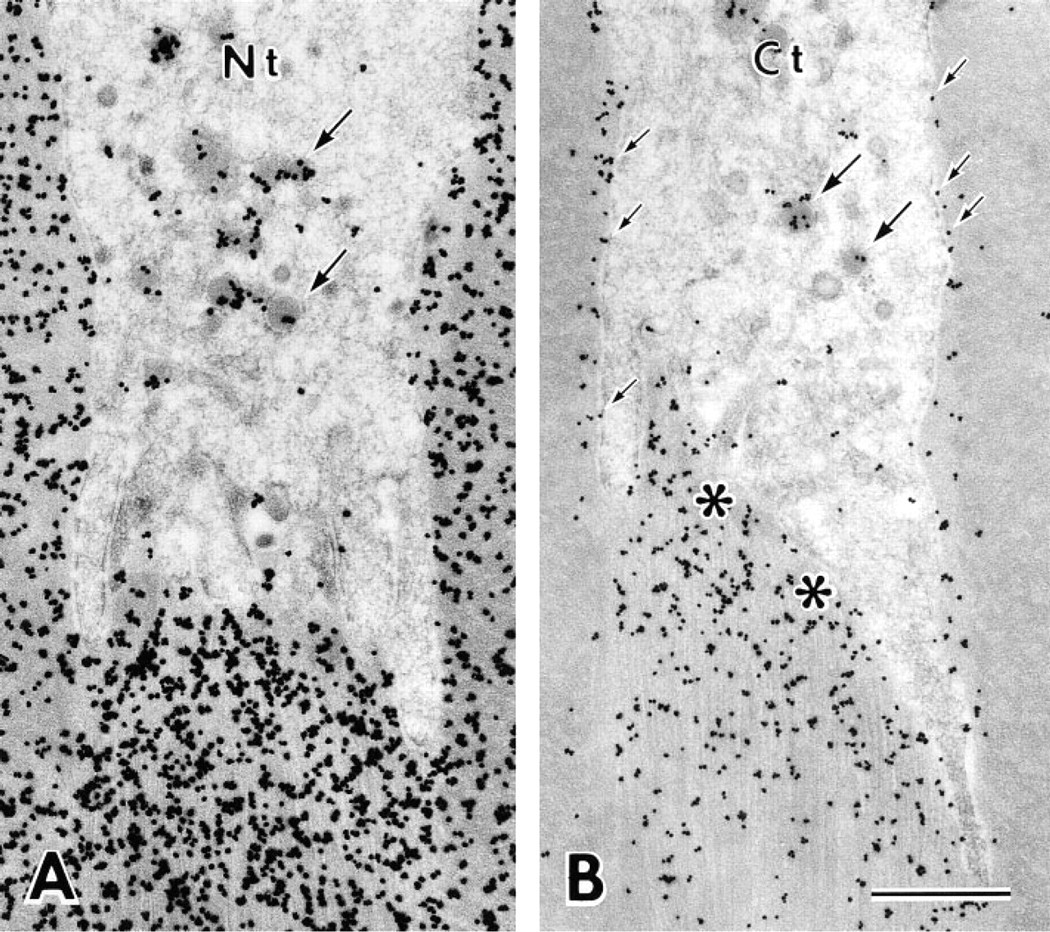
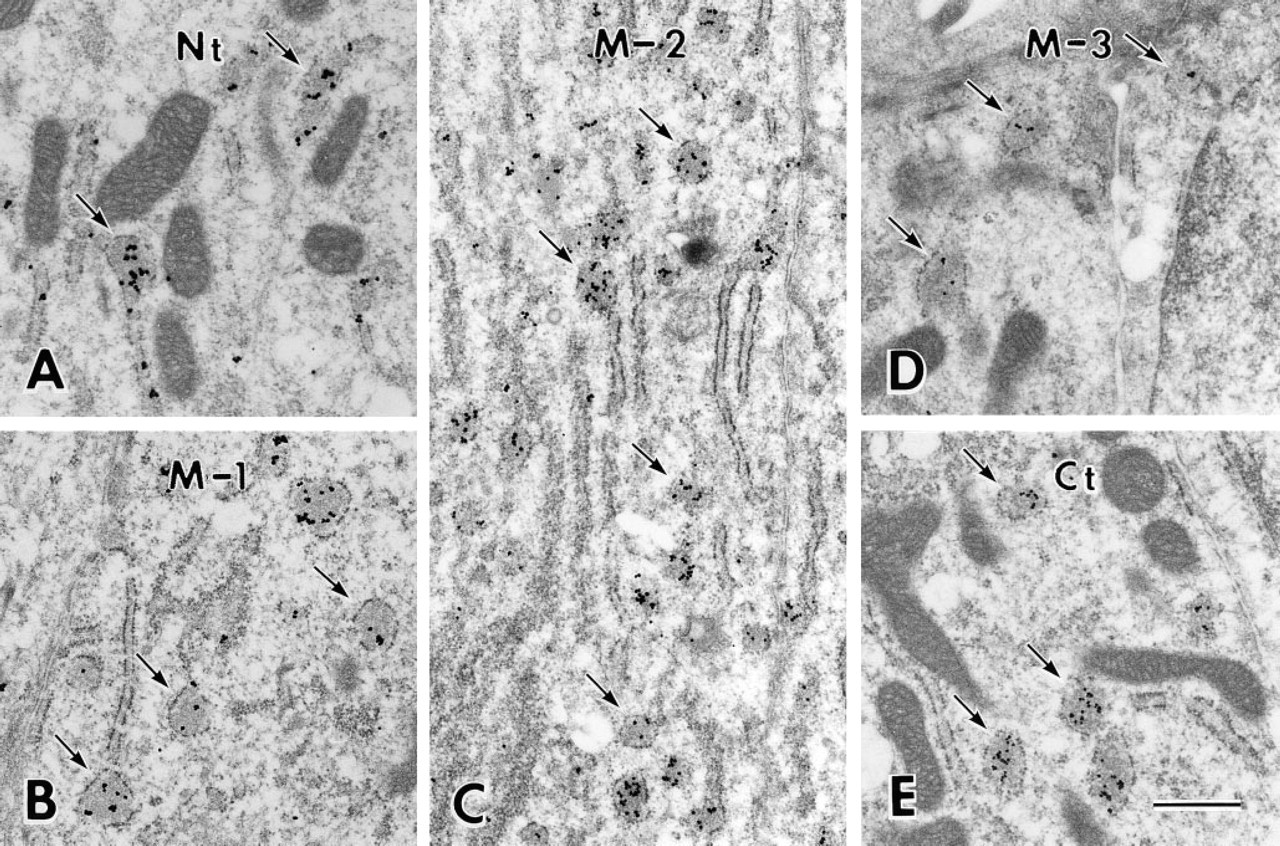
Sections cut parallel to the long axis of the interdigitating portion of the Tomes' process clearly demonstrated the relationship between the immunoreaction over the immature enamel and Tomes' processes, which are divided into proximal and interdigitating portions in the rat incisor (Warshawsky 1968) (Figure 4A). Antibody Nt intensely stained immature enamel around the Tomes' process, especially at positions adjacent to the rod and at inter-rod secretion sites, i.e., on one surface of the interdigitating portion and between two adjacent proximal portions of the Tomes' processes (Figure 4B). Antibody M-1 also stained immature enamel near the putative secretion sites, although not as distinctly (Figure 4C). Immunoreaction of antibody M-2 was weak and was restricted to near the secretion sites and the periphery of rod enamel (Figure 4D). Antibody Ct intensely stained immature enamel adjacent to secretion sites and along the lateral plasma membrane of the in-terdigitating portion. The staining along the lateral plasma membrane of the interdigitating portion of the Tomes' process appeared to be connected to the weak immunoreaction at the periphery of enamel rods (Figure 4E). In the cytoplasm of the Tomes' process, especially in the interdigitating portion, many small, dotlike immunoreactivities were observed after staining with antibodies Nt and Ct (Figures 4B and 4E).
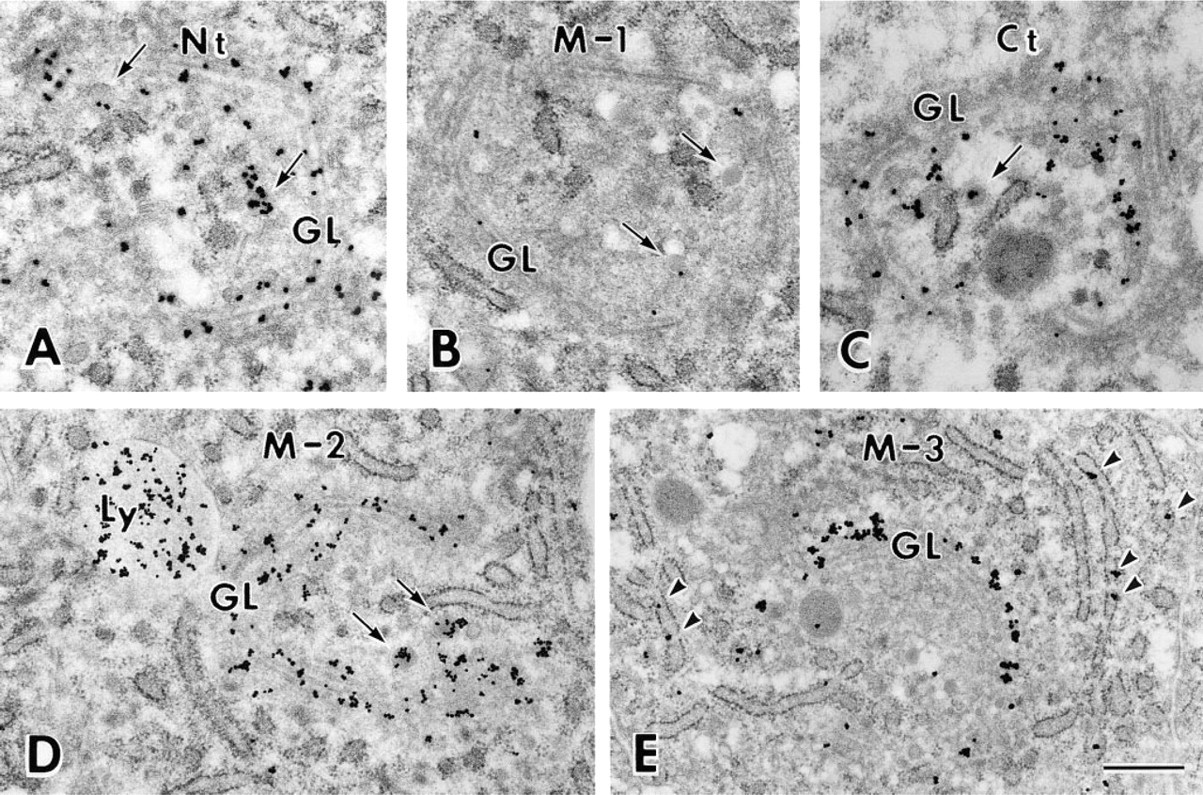
Electron micrographs of transverse sections of ameloblast showing Golgi region stained with Nt (
Electron Microscopic Immunocytochemistry
At ultrastructural resolution, silver grains representing immunoreactivity in the secretory ameloblast were found over the cell organelles involved in the synthesis and secretion as well as the degradation and absorption of proteins. A few silver grains were occasionally found over nuclei, mitochondria, and cytoplasmic matrix after staining with all the antibodies (Figures 5–7).
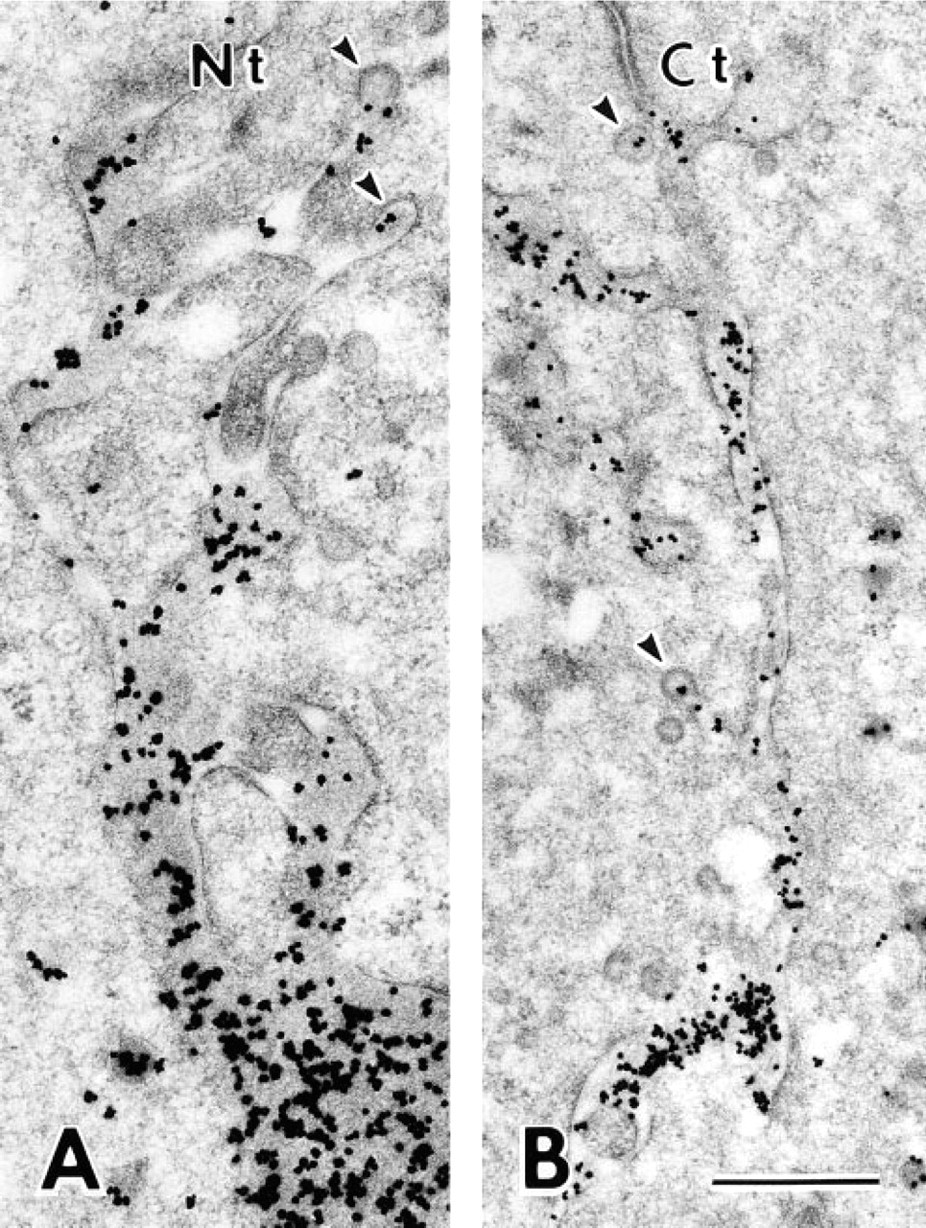
The Golgi apparatus of a secretory ameloblast is a long, cylindrical structure (Kallenbach et al. 1963). Therefore, cross-sections of ameloblasts were used to clearly demonstrate the ordered structure of the Golgi apparatus. Silver deposits representing immunoreactivity were clearly seen over the cis, medial, and trans compartments of the Golgi apparatus and secretory granules after immunostaining with antibody Nt (Figure 5A). A similar distribution of silver grains was obtained after immunostaining with antibodies M-2 (Figure 5D) and Ct (Figure 5C). After staining with antibody M-1, immunoreactivity was weak and was mainly found over the medial to the trans compartment of the Golgi apparatus. Secretory granules showed weak or no reaction (Figure 5B). Large lysosomal spherical bodies with various electron densities exhibited various intensities of immunoreactivity after staining with antibodies Nt, M-1, M-2 (Figure 5C), and Ct. The lumen of the rER was not labeled by these antibodies (Figures 5A–5D). After staining with antibody M-3, silver deposits representing immunoreactivity were restricted to the cis compartment of the Golgi apparatus. Large lysosomal spherical bodies were not stained, but the lumen of the rER was occasionally labeled (Figure 5E).
Electron micrgraphs showing interdigitating portion of Tomes' processes near the rod secretion site and surrounding enamel matrix stained with Nt (
The immunolocalization over immature enamel around the Tomes' process was mostly identical to that found at the light microscopic level, with the following additional observations obtained by immunoelectron microscopy. After staining with antibody M-2 and Ct, silver grains were located along the lateral plasma membrane of the interdigitating portion of the Tomes' process (Figure 6B). Many secretory granules in the core region of the interdigitating portion were intensely stained by antibodies Nt, M-2, and Ct (Figure 6), whereas most secretory granules were unlabeled or only weakly labeled after staining with antibody M-1. Tubulo–vesicular structures in the interdigitating portion of the Tomes' process were not labeled (Figure 6). In the proximal cytoplasm of the Tomes' process, coated and uncoated pits and vesicles, both of which apparently differed from the secretory granules, sometimes showed immunoreactivity after staining with antibodies Nt and Ct (Figure 7). No immunoreaction was found over the enamel matrix or secretory granules after staining with antibody M-3.
Effects of Brefeldin A
After the treatment with brefeldin A, immunochemical analysis of enamel organ samples revealed that the immunoreactivity of the 55-kD protein band was intensified, whereas the 65-kD protein bands were reduced or unchanged (Figure 8). The 15–20-kD protein bands stained with antibody Nt were dramatically reduced by the brefeldin A injection. In light microscopic immunocytochemical preparations, all of the antibodies stained granular structures diffusely distributed in the cytoplasm of secretory ameloblasts affected by brefeldin A. These structures were very clear when antibodies M-2, M-3, and Ct were used. The Golgi apparatus could not be recognized at the light microscopic level (Figure 9). At electron microscopic resolution, rER located in supra- and infranuclear compartments was sometimes dilated and contained flocculent material showing immunoreactivity (Figure 10). The Golgi apparatus was small and undulated but appeared to retain its normal organization. Secretory granules in the Golgi area were significantly reduced in number but were numerous in the Tomes' process. Mitochondria and nuclei remained unstained (Figure 10). There was almost no variation among the experimental animals concerning the results of immunochemical and immunohistochemical analyses. The immunochemical and immunohistochemical data obtained from the control rats injected with solutions used for dissolving brefeldin A were the same as those of nontreated rats.
Specificity Control
Immunostaining over the ameloblast and the immature enamel almost disappeared after preabsorption of the antibodies with purified peptide.
Discussion
Immunochemical analyses showed that the highest molecular weight protein band that reacted to all the antibodies used (excepting M-3) was 65 kD in each case. In addition, all antibodies stained cell organelles involved in the production and secretion of proteins in secretory ameloblasts, although intensities varied. Therefore, we conclude that the 65-kD protein is likely to be the largest form of ameloblastin.
Immunoelectron microscopy demonstrated that antibody M-3, which specifically recognizes residues 386–399 of ameloblastin, stained only the rER and the cis side of the Golgi apparatus. Downstream Golgi elements, secretory granules, and enamel matrix showed no immunoreactivity. In immunochemical preparations of the enamel organ, the M-3 antibody reacted to a protein band near 55 kD but not the 65-kD band. After treatment with brefeldin A, which inhibits protein transport from the rER to the Golgi apparatus (Fujiwara et al. 1988), immunoreaction of the 55-kD protein band intensified and was found within the dilated cisternae of the rER irrespective of the antibody used.
Electron micrographs of two adjacent proximal portions of the Tomes' processes stained with Nt (
These results indicate that an epitope(s) located at residues 386–399 of nascent ameloblastin is masked by post-translational modifications occurring in the Golgi apparatus. Ameloblastin contains no putative N-glycosylation sites (Krebsbach et al. 1996). Therefore, we suggest that ameloblastin is initially synthesized as a 55-kD core protein in the lumen of the rER and that O-glycosylation of a serine or threonine residue located between residues 386–399 generates the 65-kD glycoprotein within the Golgi apparatus.
Detection of proteins extracted from brefeldin A-affected (Br) and unaffected (U) enamel organs by SDS-PAGE (
The calculated molecular mass of ameloblastin in the absence of post-translational modifications is 45 kD (Krebsbach et al. 1996) and that without the putative signal peptide is about 42 kD. Lee et al (1996) performed Western blot analysis using antibody raised against recombinant ameloblastin that contained residues 205–378 of ameloblastin, and suggested that the protein bands appearing near 44 and 47 kD correspond to the core protein. These are significantly lower than the apparent molecular weight, near 55 kD, shown by SDS-PAGE in the present study. This is not necessarily surprising, considering the anomalous behavior of a bone matrix protein, osteopontin, whose apparent molecular weight on SDS-PAGE depends upon the concentration of the gel used (Kubota et al. 1989), since we electrophoresed on 15% gels and Lee et al. (1996) used 4–20% gradient gels.
In the present Western blot analyses, no immunoreactive protein band larger than 65 kD was found in either the enamel organ sample or the enamel matrix sample. On the other hand, Lee et al. (1996) have found immunoreactive protein bands near 84 and 120 kD. These proteins might represent a protein complex with other enamel proteins, as Lee et al. (1996) speculated.
Immunochemical analyses showed that the antibodies that specifically recognize ameloblastin residues 27–48 (Nt) and 98–107 (M-1) reacted with 10-kD to 23-kD proteins but not with proteins whose molecular weight was higher than 25 kD, except for the 55-kD and 65-kD proteins. In immunohistochemical preparations, these antibodies stained the entire thickness of immature enamel. On the other hand, the antibody that recognizes residues 224–232 (M-2) stained protein bands near 65 kD and diffuse bands from 56 to 40 kD. The antibody that recognizes residues 406–419 (Ct) stained several protein bands ranging from 25 kD to 65 kD. These two antibodies (M-2 and Ct) stained enamel matrix located near the putative secretion sites in immunocytochemical preparations. These results indicate that the initial cleavages of the 65-kD amelo-blastin generate relatively small polypeptides containing the N-terminal and relatively large polypeptides containing the C-terminal portions of the secreted protein. The N-terminal polypeptides appear to be rather stable and are gradually degraded, but are not lost, from the enamel matrix during the matrix formation stage. The C-terminal large polypeptides appear to be degraded rapidly and are lost from the immature enamel soon after secretion.
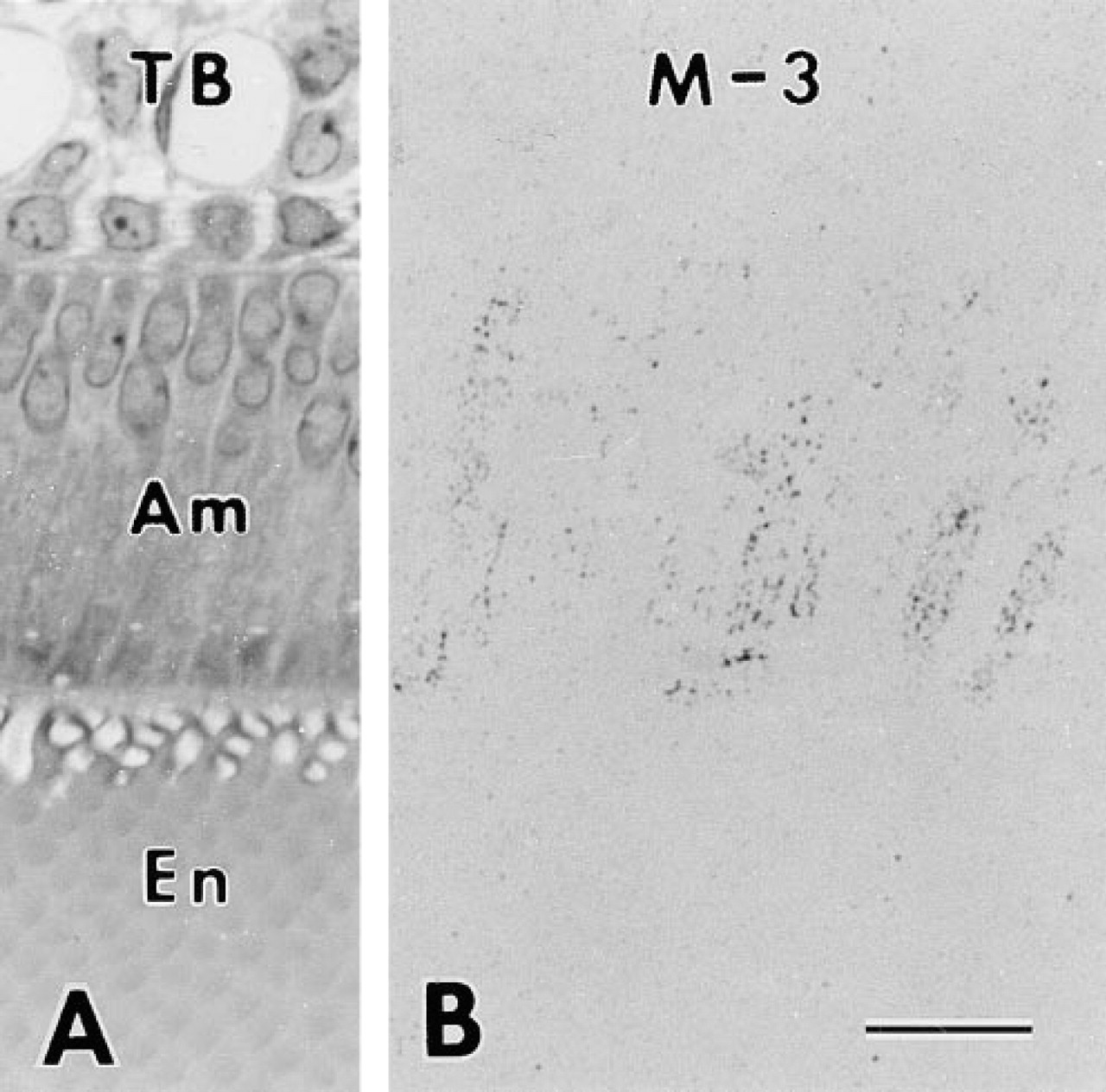
Light micrographs of secretory ameloblast (Am), immature enamel (En), and dentin (De) at the stage of inner enamel secretion from longitudinal sections of a rat incisor after injection of brefeldin A. (
Electron micrographs of proximal (
Electron microscopic immunocytochemistry identified coated pits or vesicles containing ameloblastin and/or its cleaved polypeptides. These data support the hypothesis that secretory ameloblasts absorb enamel matrix proteins (Katchburian and Holt 1969; Ozawa et al. 1983). The absorptive activities might be partially responsible for the loss from the enamel matrix of cleavage products containing the ameloblastin C-terminal region soon after their secretion.
Our previous immunohistochemical study using antiserum against porcine low molecular weight sheath proteins demonstrated no specific staining pattern over the immature enamel of the rat incisor (Uchida et al. 1991a). In this study, however, an antibody was used that specifically recognized the amino acid sequence of residues 98–107 and intensely stained the periphery of each enamel rod in the middle to inner layer of immature enamel, especially at the late matrix formation stage. These results indicate that some of the N-terminal-cleaved peptides of ameloblastin concentrate in the prism sheath of immature enamel as the porcine sheath protein does. The immunolocalization of the C-terminal region of ameloblastin shown in this study is very similar to that of the sheathlin (Murakami et al., in press). In addition, the possible degradation pattern of sheathlin (Murakami et al., in press) is very similar to that of ameloblastin. Taking all of these observations into consideration, we propose that sheathlin and ameloblastin share the same physiological role in amelogenesis and should both be classified into the group of enamel proteins designated as sheath proteins. However, the initial formation of prism sheath along the nonsecretory face of the Tomes' process found in the porcine tooth germ (Uchida et al. 1995) was not observed in the rat incisor. Therefore, we also suggest that the mechanisms through which prism sheath is developed in the rat incisor are somewhat different from those of the pig.
Silver deposits representing immunoreactivity of the C-terminal region of ameloblastin were found along the plasma membrane of the interdigitating portion of the Tomes' process. These results, together with the fact that ameloblastin contains consensus sequences for peptide domains interacting with cell surface proteins (Cerny et al. 1996), suggest that the plasma membrane of the interdigitating portion of the Tomes' process is connected to the enamel matrix via ameloblastin and/or its cleavage products. The functional significance of ameloblastin as a cell binding protein has not been elucidated. It is well known that the Tomes' process in rat is extremely long relative to other animal species such as pig, cat, and human, and is deeply embedded in immature enamel. Hence, the distal portion of the Tomes' process of the rat has been called the interdigitating portion (Warshawsky 1968). It is not known how these long processes form and maintain their shape. At one surface of the process, there is a putative rod secretion site (Weinstock and Leblond 1971; Nanci and Warshawsky 1984) at which the membrane of secretory granules are fused continuously via exocytosis. To support the peculiar shape of the Tomes' process, it appears likely that secretory ameloblasts must maintain a steady state, balancing the addition of new membrane via exocytosis with membrane retrieval (Uchida and Warshawsky 1992). It is tempting to speculate that ameloblastin prevents the separation of the enamel matrix from the plasma membrane during this dynamic turnover of membrane material.
Using autoradiography of sections of rat incisor and fluorographs of proteins extracted from rat enamel organ and immature enamel after injection of various radioactive molecules, Smith and co-workers have revealed basic biochemical properties of short-lived sulfated glycoproteins (Smith et al. 1995; Smith and Nanci 1996). The apparent molecular masses of some of these proteins (65, 49, 42, 38, 35, and 25 kD) were very similar to those of the post-translationally modified ameloblastin shown in this study (65, 48, 41, 36, 34, and 25 kD). Furthermore, the molecular mass of the 57-kD protein, which Smith and Nanci (1996) have supposed to be a nascent protein that undergoes post-translational modifications to become the 65-kD sulfated enamel protein, is also very similar to that of the 55-kD protein identified in this study. Density scans of fluorographs for proteins in the enamel organ at 1 or 10 min after single injections of radioactive methionine and tyrosine have shown that intensity of radiolabeling of this 57-kD protein was the highest next to the 27-kD protein, which corresponds to amelogenin (Smith and Nanci 1996). Matsuki et al. (1995) constructed an unidirectional cDNA liberally from the growing end of the rat incisor, sequenced cDNA clones, classified their sequence, and showed that, among the enamel matrix-specific genes, the most redundant mRNAs next to amelogenin were for ameloblastin. These results raise the possibility that ameloblastin corresponds to the sulfated glycoprotein demonstrated by Smith et al. (1995). However, experiments with glycosidases suggested that the majority of sulfate groups were attached to sugars that were connected to the protein via asparagine side-chains (N-linkages), resulting in the 65-kD sulfated glycoprotein (Smith et al. 1995). Computer analysis of the ameloblastin cDNA sequence recognized no N-glycosylation target sites (Krebsbach et al. 1996). Further studies are needed to clarify the relationship between ameloblastin and the sulfated enamel proteins reported by Smith et al. (1995).
Footnotes
Acknowledgements
Supported in part by Grants-in-aid for Scientific Research from the Ministry of Education, Science and Culture of Japan (Nos. 08672071, 09671855 and 09771502).
We thank Profs J.P. Simmer and J.C-C. Hu, Department of Pediatric Dentistry, University of Texas Health Science Center at San Antonio, School of Dentistry, who read the manuscript and made helpful comments. We also thank the Research Center for Molecular Medicine, Hiroshima University, for use of the peptide synthesizer PSSM-8.