
Abstract
Incorporation of halogenated nucleotide analogues is often used to assess DNA synthesis and to quantitate cellular proliferation. Multiple antibodies have been developed to bromodeoxyuridine (BrdUrd) and it is the most frequently utilized substrate. Because the immunodetection of incorporated BrdUrd requires DNA denaturation or nuclease digestion, most of these antibodies are not reactive in tissues or cells fixed with crosslinking agents. Antigen retrieval techniques utilizing protease digestion restore BrdUrd antigenicity and permit the detection of BrdUrd in formalin-fixed tissue. However, during the development of a double label immunohistochemical protocol to quantitate proliferating alveolar Type II cells, we noted nucleus-specific staining in lung sections from animals that had not received BrdUrd. Therefore, we systematically analyzed the specificity of the immunohistochemical detection of incorporated BrdUrd in formalin-fixed tissue after protease digestion. Enzymatic antigen recovery diminished the specificity of the BrdUrd reaction product and caused false-positive staining with the BU-1, B44, and BR3 monoclonal antibodies. Staining was less prominent with Bu20a but was more specific. Protease antigen recovery may decrease the specificity of BrdUrd immunodetection. Appropriate controls are required when enzymatic digestion is used to detect incorporated BrdUrd in formalin-fixed tissue. The type and duration of fixation, antibody to BrdUrd, protease, and tissue may affect the specificity of the staining pattern.
In the development of a double label immunohis-tochemical staining protocol to quantitate proliferating alveolar Type II cells, we found that antigen retrieval with several different proteases decreased the specificity of various MAbs for BrdUrd (Panos et al. 1995, 1996). Therefore, in this study we systematically determined the effect of various protease antigen retrieval methods on the detection of BrdUrd incorporated into tissue fixed in formalin. We show that protease antigen recovery may decrease the sensitivity of antibodies to BrdUrd, causing false-positive results. Therefore, appropriate negative controls should be utilized in immunohistochemical protocols using enzymatic antigen recovery to detect BrdUrd in formalin-fixed tissue sections.
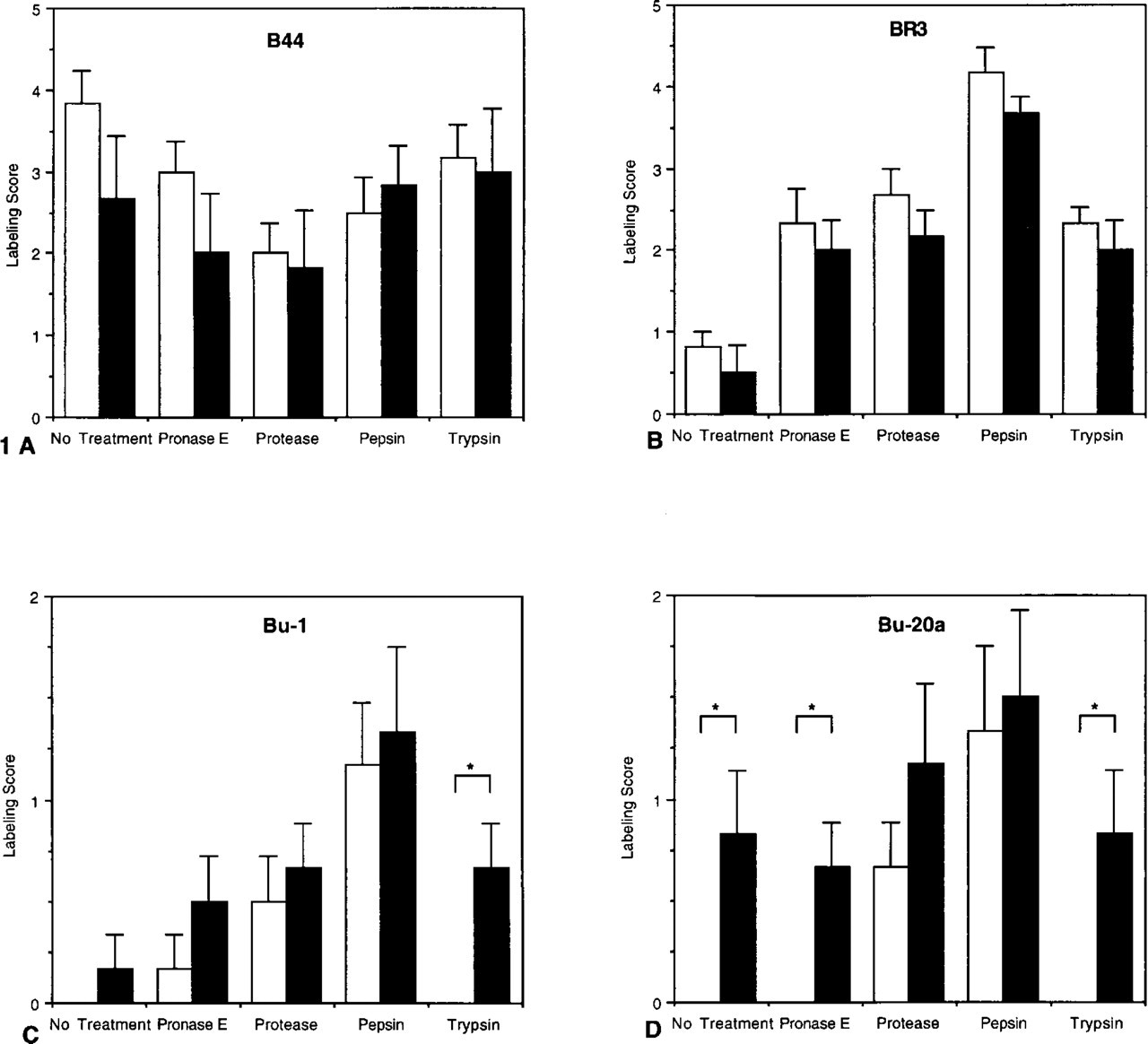
Labeling scores for tissue sections from animals treated with BrdUrd (solid bars) or PBS (open bars) and fixed for 18 hr. Enzyme concentrations for antigen recovery, B44 (
Materials and Methods
Male Sprague–Dawley rats (225–250 g) were injected
The sections were then incubated with biotinylated horse anti-mouse (heavy and light chain-specific) IgG (Vector Laboratories; Burlingame, CA) diluted 1:200 in 3% horse serum/0.1% Triton X-100 for 1 hr at RT and then washed in two changes of PBS for 5 min each. Biotin–streptavidin–horseradish peroxidase diluted 1:100 in PBS was added for 30 min at RT. After washing in two changes of 0.1 M phosphate buffer, pH 7.4, the horseradish peroxidase activity was detected using Ni/Co enhancement of the diaminobenzidine reaction product (Adams 1981). The sections were then washed twice in ddH2O, dehydrated by passing through a graded series of ethanol solutions (30%, 50%, 70%, 100%, 100%, v/v), cleared in xylene, and mounted with Permount.
All sections were coded and independently graded by both investigators, who rated the slides on a scale of 0 to 5: 0, no staining; 1, sporadic nuclear staining in basal cells; 2, nuclear staining in all basal cells; 3, staining in basal cells throughout the section and extending to the intermediate cells intermittently; 4, staining in basal cells and extending to the intermediate cells throughout the section; 5, nuclear staining throughout the section. Data are expressed as the mean ± SEM of three separate experiments. Statistical analysis was performed using an unpaired t-test (StatView; Abacus Concepts) and p>0.05 was considered significant.
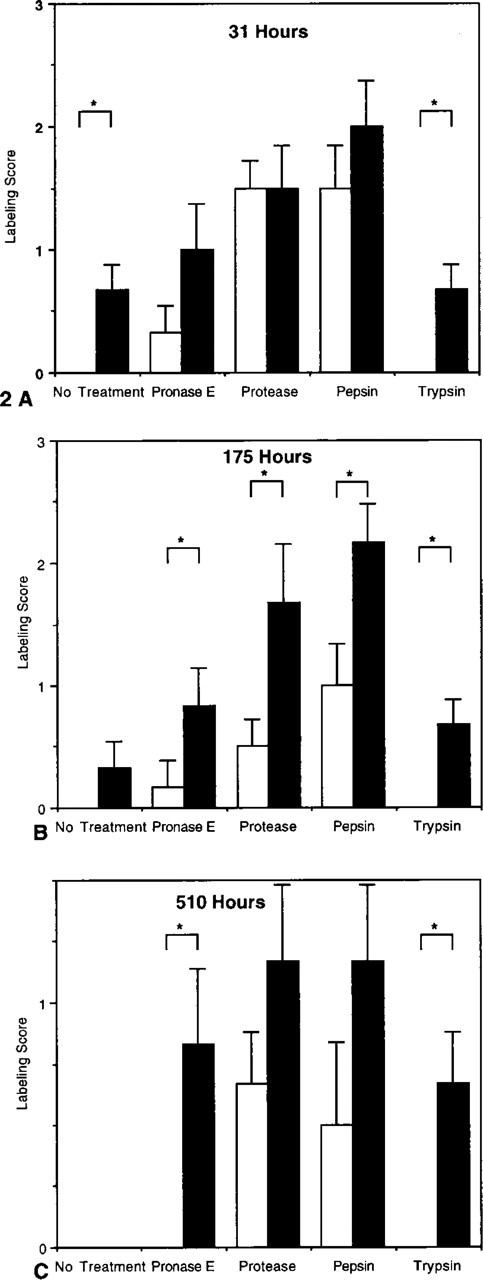
Labeling scores for tissue sections from animals treated with BrdUrd (solid bars) or PBS (open bars) and fixed for 31 (
Results
Antigen recovery with the different enzymes significantly affected the detection of BrdUrd (Figure 1). With both the B44 and BR-3 anti-BrdUrd MAbs, nuclear staining was present in unlabeled tissues that were not treated with proteases as well as in the enzymatically digested specimens. There were no significant differences in the labeling scores between specimens from BrdUrd-treated and control animals. No nuclear labeling was present in tissues from saline-instilled rats using the BU-1 MAb and either no antigen recovery or trypsin treatment. Pronase E, protease, and pepsin digestion produced equivalent labeling scores in BrdUrd-labeled and unlabeled specimens.
Bu20a was the most specific MAb and accurately discriminated BrdUrd-labeled tissues from unlabeled sections after either no treatment, pronase E, or trypsin digestion. However, after protease or pepsin antigen recovery, there were no significant differences between BrdUrd-treated and control animals.
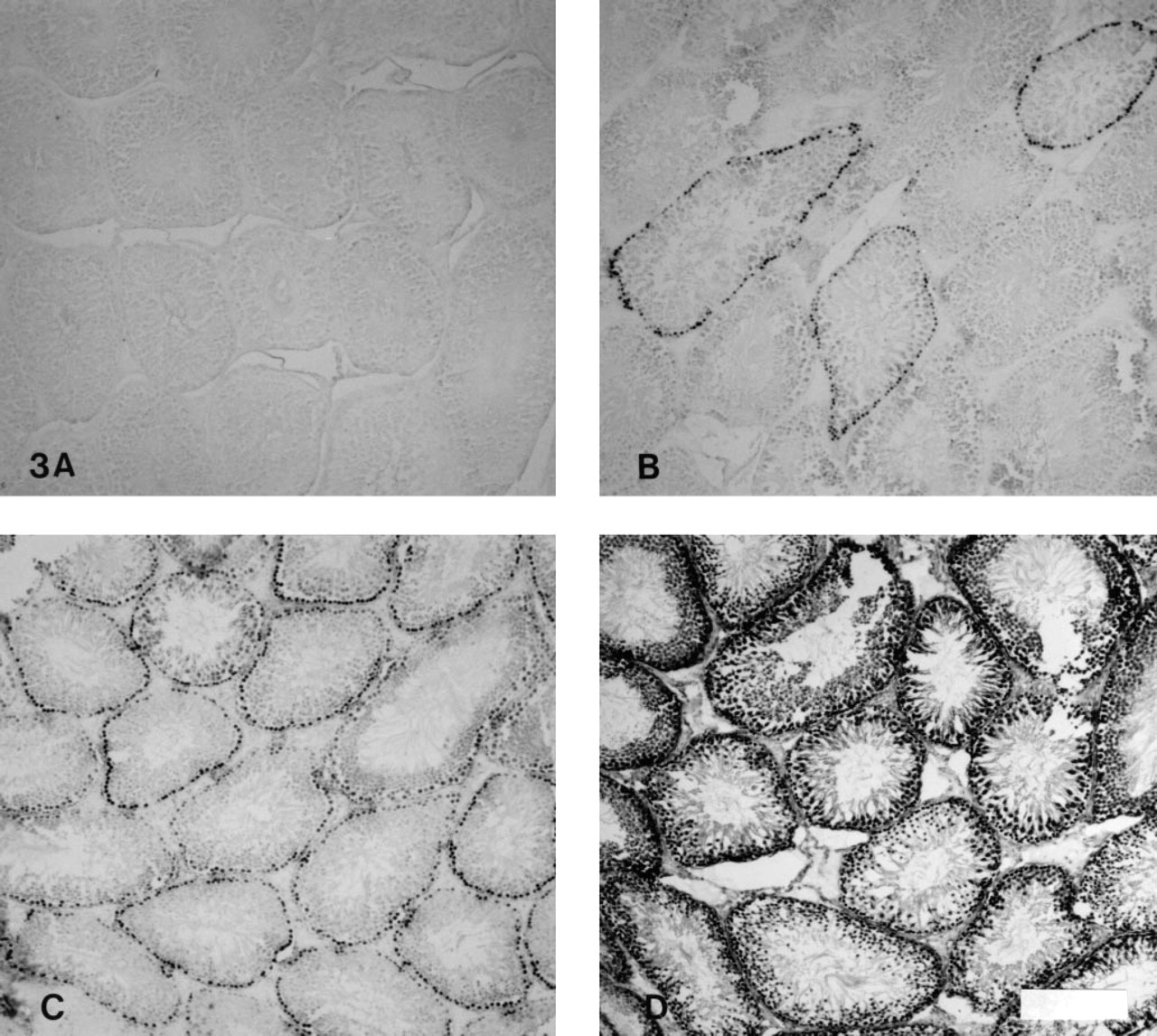
(
To determine the effect of fixation duration on Bu20a detection of incorporated BrdUrd, tissue was fixed for 31–510 hr in buffered formalin (Figure 2). After 510 hr of fixation, no staining was detected in untreated sections, whereas at earlier time points Bu20a accurately discriminated between labeled and unlabeled tissues. Nuclear staining was present in unlabeled tissues with all the enzymatic treatments except trypsin. The highest levels of nonspecific staining occurred with protease and pepsin antigen recovery (Figure 3).
Discussion
Many MAbs to BrdUrd have been developed. A halogenated ribouracil conjugated to a carrier protein has been the immunogen used in the creation of many of these antibodies. They are generally of low affinity (Ka approximately 106 M-1 for BrdUrd in solution) and detect the halogenated nucleotide only in single-stranded but not in double-stranded DNA (Adams 1981). Most methods for the immunodetection of incorporated BrdUrd require tissue to be frozen or fixed in dehydrating agents such as methanol/acetic acid (Gratzner 1982), acetone (Morstyn et al. 1983), or ethanol (Vanderlaan and Thomas 1985), and that DNA be denatured by acid hydrolysis, exposure to strong bases or formamide, or degraded by nuclease digestion. BrdUrd detection is prevented or significantly impaired by fixation in crosslinking agents such as formalin owing to prevention of DNA denaturation or irreversible crosslinking of nuclear proteins to DNA, thus blocking BrdUrd recognition. DNA containing BrdUrd or IdUrd has higher affinity for chromatin proteins, including histones and regulatory proteins, which may increase crosslinking of nuclear protein to areas of DNA containing halogenated nucleotides (Hamada 1985; Lin 1978). Enzymatic digestion of formalin-fixed tissue has been shown to restore BrdUrd antigenicity and permit immunodetection of BrdUrd within formalin-fixed tissue (Hayashi et al. 1988; Shutte et al. 1987; Sugihara et al. 1986).
We utilized protease antigen recovery in the development of a double label immunohistochemical technique to quantitate proliferating alveolar Type II cells (Panos et al. 1995, 1996). During the development of this protocol, we noted decreased specificity of BrdUrd detection and occasional nuclear staining in specimens that had not been exposed to BrdUrd. In this study, we systematically analyzed the specificity of the immunohistochemical reaction product in formalin-fixed tissue after protease digestion. Enzymatic antigen recovery diminished the specificity of the BrdUrd reaction product and caused false-positive staining. The BR3 MAb, which uses nuclease digestion to expose incorporated BrdUrd, produced the most intense staining and the greatest nonspecificity. In contrast, the Bu20a MAb yielded the lightest staining but was the most specific.
The cause of the false-positive staining is uncertain. Other investigators have suggested that enzymatic digestion may unmask or create antigenic epitopes common to unrelated tissue constituents (Heyderman 1979). Therefore, it is possible that proteolytic antigen recovery might produce common epitopes between halogenated and nonhalogenated nucleotides. In an analysis of IU-1, IU-2, and B-44 MAbs to BrdUrd, Vanderlaan and co-workers (Vanderlaan and Thomas 1985) demonstrated reactivity with BrdUrd incorporated into single-stranded but not double-stranded DNA. These antibodies also bound weakly to thymidine and to unsubstituted single-stranded DNA. In our study, staining was localized to the nucleus in specimens that were not labeled with BrdUrd, suggesting nucleus-specific rather than nonspecific background staining. Finally, the specificity of BrdUrd detection after protease antigen recovery may differ for various MAbs to BrdUrd. DNAse treatment does not permit BrdUrd detection with B44 but does work with the BU-1 MAb (Gonchoroff et al. 1986).
In a study comparing BrdUrd with [3H]-thymidine labeling in spermatagonial stem cells and Leydig cells in the testes and small intestinal crypt cells, Thoolen (Thoolen 1990) used microwave antigen recovery after fixation in Carnoy's fluid and found that 90% of the BrdUrd-labeled cells were also identified by [3H]-thymidine autoradiography. These studies used the B44 anti-BrdUrd MAb. Although the correlation between the BrdUrd and [3H]-thymdine labeling indices was highly significant statistically, no explanation was given for the increased recognition of proliferating cells with microwave antigen recovery and BrdUrd immunohistochemistry compared with [3H]-thymdine autoradiography.
Immunohistochemical detection of BrdUrd has been widely used to determine cell proliferation in cells and tissues. Enzymatic antigen recovery allows BrdUrd immunodetection in tissues preserved with aldehyde-containing fixatives. Careful and appropriate controls are required when protease digestion is used to determine a proliferative index based on BrdUrd immunodetection in formalin-fixed tissue specimens.
Footnotes
Acknowledgements
This work was funded by the Department of Veterans Affairs.