
Abstract
We investigated the numbers of DNA-synthesizing cells in mouse submandibular glands (SMGs) during chronic isoproterenol (IPR) administration, using an immunohistochemical method with anti-bromodeoxyuridine (BrdU) antibodies. Adult and immature female mice were injected with IPR daily or every other day up to a total of seven injections. Whereas hypertrophic enlargement of acinar cells continued with repeated IPR injections, induction of DNA synthesis was transient. With one IPR injection, more than 50% of the acinar cells were induced to synthesize DNA, but after five daily injections the numbers of cells in the DNA synthetic phase decreased and returned to control values. This pattern was basically the same for immature mice and for adult mice stimulated by IPR every other day. The responsiveness of the SMGs of immature mice to IPR was somewhat lower than that of adults. Prolongation of the interval between injections of IPR, compared to daily injections, did not cause sustained induction of DNA synthesis. These data suggest that the hypertrophic enlargement and the induction of DNA synthesis by IPR are mediated through different intracellular pathways after the stimulation of β-adrenergic receptors, and that the regulatory mechanism for the induction of DNA synthesis is complex.
Enlargement of the PAG and SMG caused by chronic administration of IPR is believed to involve both hypertrophy, which reflects the excessive activation of protein synthesis and the enlargement of acinar cells, and hyperplasia, associated with cell proliferation. Barka et al. (1965a) reported that a single injection of IPR in rats caused conspicuous stimulation of DNA synthesis in the SMGs. However, whether the hyperplasia and hypertrophy involve the same intracellular pathway after the β-adrenergic stimulation is not known.
Over the past 10 years, immunohistochemistry for the detection of bromodeoxyuridine (BrdU) incorporated into DNA has been widely used to detect cells in S-phase that are synthesizing DNA, instead of autoradiography of [3H]-thymidine (Gratzner 1982; Yu et al. 1992; Dolbeare 1995). Immunohistochemical staining of plastic-embedded sections with anti-BrdU (Harms et al. 1986) has improved the localization of DNA synthesizing cells in tissues.
This study focused on the localization pattern of DNA-synthesizing cells in the SMG during chronic IPR administration in adult (10-week-old) and immature (4-week-old) female Balb/c mice, using the BrdU method, to confirm whether the cascade of cellular processes that leads to hypertrophy and hyperplasia is similar.
Materials and Methods
Animals and Tissue Preparation
Female Balb/c mice 3 and 9 weeks of age were purchased from Japan SLC (Shizuoka, Japan). The animals had free access to water and laboratory chow and were kept in alternating 12-hr periods of light (0600–1800) and darkness. The experimental protocol was approved by an institutional panel. In the first experiment, 10-week-old mice were given daily
All animals were sacrificed at 24 hr after the last IPR injection and were injected with 5-bromo-2'-deoxyuridine (Sigma) at a dose of 0.3 μmole/g body weight 1 h before sacrifice. SMGs were dissected out under
Immunohistochemistry
DNA-synthesizing cells were detected by peroxidase–antiperoxidase (PAP) immunostaining with anti-BrdU antibodies by the method of Harms et al. (1986), which was slightly modified as follows. Sections were treated with 2 N HCl at 60C for 15 min and washed in running water for 15 min. After dehydration in an ethanol series, the sections were etched with xylene (15 min, twice). Nuclear proteins, which mask incorporated BrdU, were digested with 0.025% protease (Type V; Sigma) in PBS at 37C for 10 min, and the sections were then washed with cold PBS three times for 5 min. For immunostaining, the sections were first treated for 30 min at room temperature (RT) with normal rabbit serum at a 1:20 dilution to reduce the nonspecific background. Sera were wiped away and then the sections were incubated with a mouse monoclonal antibody against BrdU (Boehringer Mannheim Biochemicals; Indianapolis, IN) at a dilution of 1:200 at 4C for 20 hr, and then with rabbit antibody against mouse IgG (Kirkeggard & Perry Laboratories; Gaithersburg, MD) at a dilution of 1:100 for 2 hr at RT, and finally with mouse monoclonal–PAP complex (Sigma) at a dilution of 1:500 for 1 hr. Peroxidase activity was visualized by immersion for 20 min in 0.01% 3,3'-diaminobenzidine (Sigma) and 0.005% H2O2 in 0.05 M Tris buffer, pH 7.6. Immunostainings for each study were done at the same time. The immunostained sections were counterstained with hematoxylin.
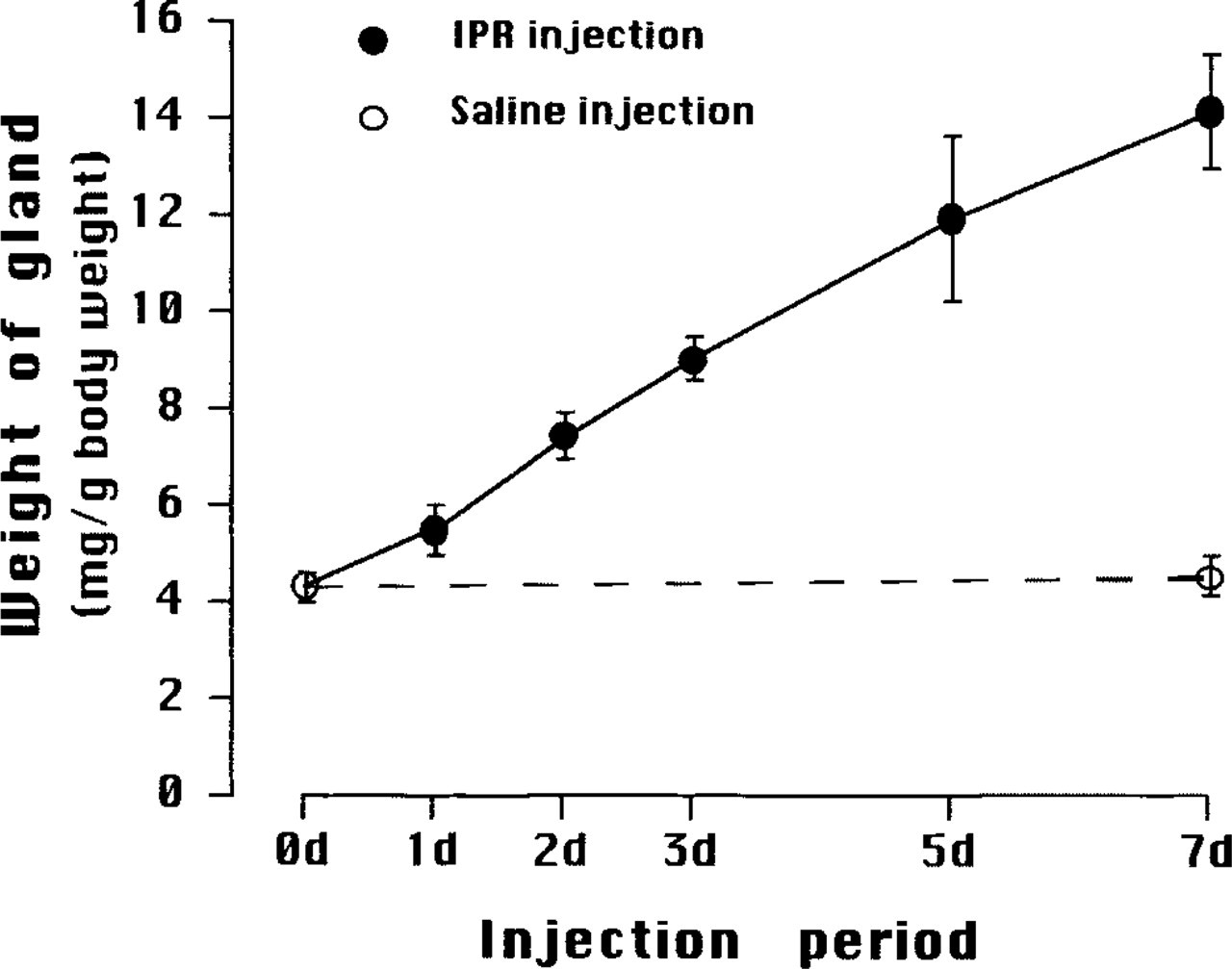
Changes in relative weight of SMGs of adult mice during IPR injections. Control mice were intact (0 day, 0 d) or saline-injected daily for 7 days (open circles), and experimental mice received daily IPR injections for up to 7 days (7 d) (solid circles). Mean values ± SD.
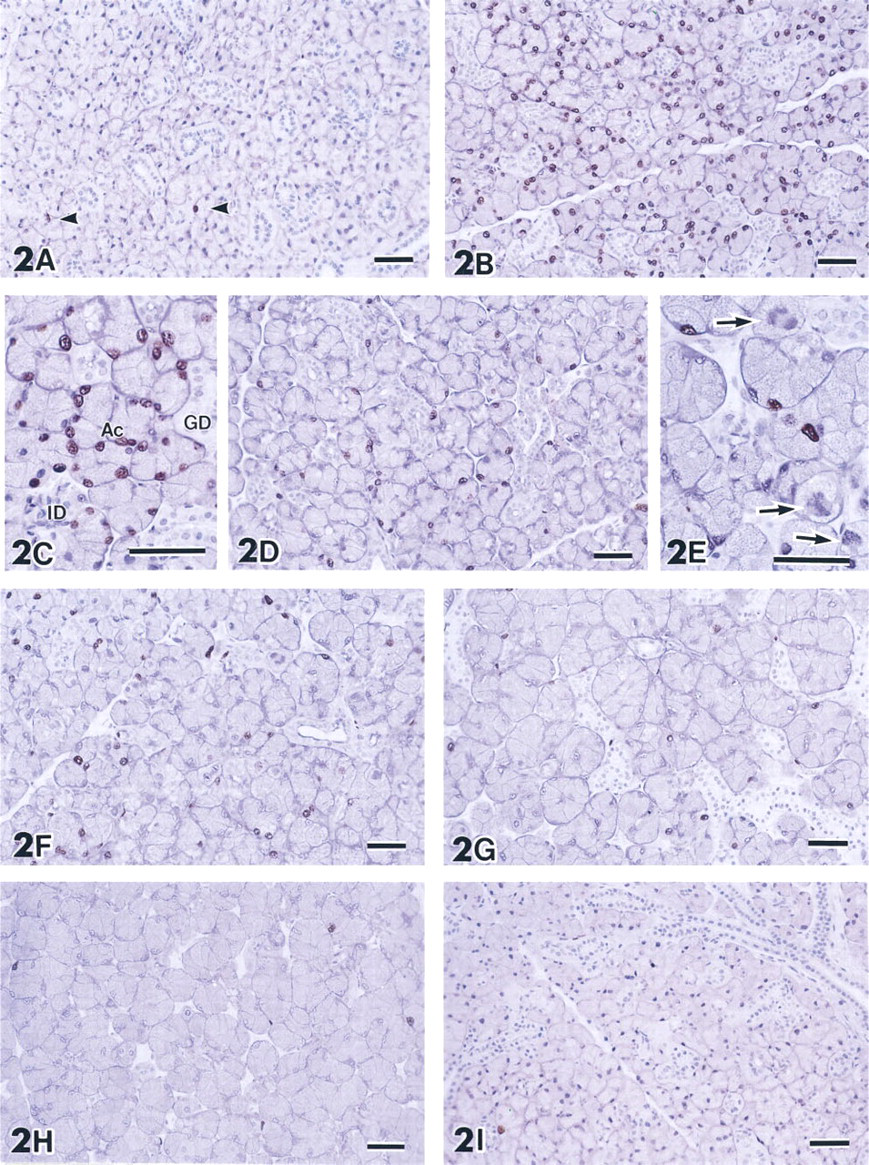
SMGs of adult mice immunohistochemically stained with anti-BrdU antibodies. (
In micrographs enlarged to a final magnification of x 250, the immunolabeled and nonlabeled acinar cells nuclei were counted, and the percentages of cells in S-phase were calculated for each experimental group. From each gland, 800–1000 nuclei were counted.
Results
In control mice, the relative weight of the SMG (4.3 ± 0.3 mg/g bw) did not change after 10 weeks of age. On the other hand, the relative SMG weights of mice treated with IPR increased linearly with the numbers of daily injections, reaching about 3.5 times control (14.12 ± 1.2 mg/g bw) after seven injections (Figure 1).
Immunohistochemical staining of SMGs of untreated mice with anti-BrdU showed that only a few cells incorporated BrdU, indicating that almost all cells in SMGs of mature mice are quiescent, i.e. in the G0-phase (Figure 2A). This was also the case for the glands of control mice that received saline injections for 7 days (Figure 2I). One injection of IPR caused a marked increase in the number of nuclei immunolabeled with anti-BrdU (Figure 2B). Immunolabeled nuclei were specifically localized in acini. Nuclei of duct cells, including intercalated duct (ICD), granular convoluted tubule (GCT), and striated duct (SD), did not incorporate BrdU (Figure 2C). After further IPR injections the number of immunolabeled nuclei decreased (Figures 2D and 2F–2H). In the SMGs of mice that received daily IPR injections for 2 or 3 days, mitotic figures were often seen in the acinar cells (Figure 2E). The acinar cells increased in size after 2 days of IPR treatment. The numbers of DNA-synthesizing cells after 5 (Figure 2G) or 7 (Figure 2H) days of IPR treatment were the same as in intact or saline-injected control mice. Mitotic figures in acinar cells were also rare in the SMGs of mice treated with IPR for 5 or 7 days. The duct system of the SMGs of the mice treated with IPR for 7 days appeared to be atrophic, perhaps due to hypertrophy of the acinar cells.
The structure of both labeled and unlabeled nuclei of acinar cells in the glands of mice that received IPR was less well-preserved after the hydrolysis and digestion procedures, which are essential for immunostaining with anti-BrdU, compared to that of the duct cells or acinar cells in the glands of control animals.
The percentage of BrdU-positive nuclei in acinar cells during chronic IPR administration is shown in Figure 3. In control SMGs, only 1.5 ± 0.5% of acinar cells incorporated BrdU. In contrast, after one injection of IPR more than half of the nuclei of acinar cells, 52.5 ± 3.0%, incorporated BrdU. However, with repeated stimulation with IPR the numbers of labeled nuclei declined to 24 ± 3.5% after two IPR injections, 13.5 ± 3.0% after three injections, and 5.0 ± 2.5% after five injections. After seven daily injections of the drug the labeling index, 0.7 ± 0.5%, was comparable to that of controls. Therefore, the induction of DNA synthesis by chronic IPR stimulation was a transient response, whereas the hypertrophic effect was progressive and the size of the gland increased with the time of IPR treatment. To analyze this phenomenon further, we carried out two experiments. In the first experiment we used immature 4-week-old female mice and the same experimental design as in the first study. In the second experiment, adult mice were utilized but the time between the IPR injections was increased.
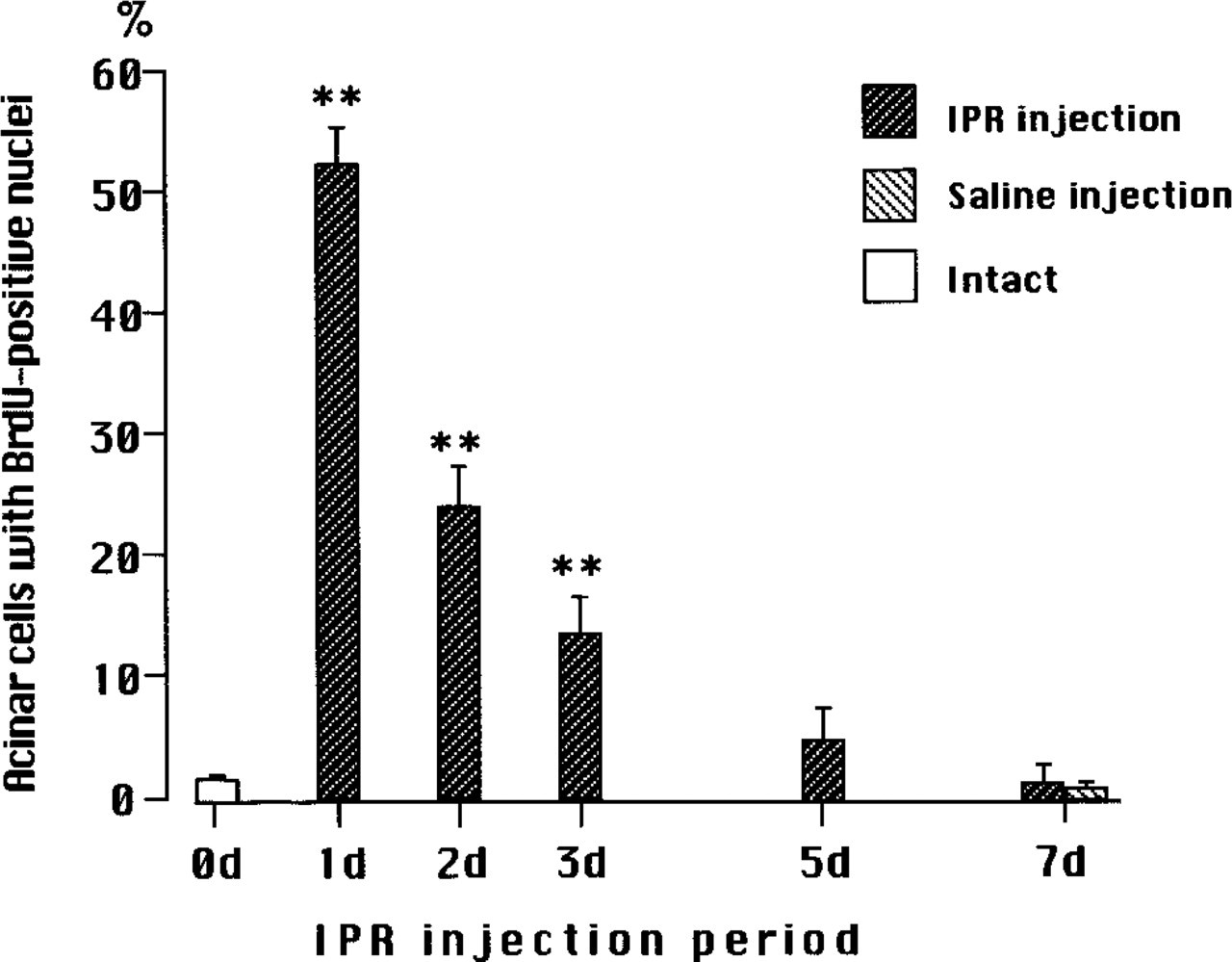
Changes in percentages of BrdU-positive nuclei in acini of SMGs of adult mice during chronic IPR administration. Mean values ± SD. Intact mice are represented as 0 day (0 d). Statistical significances between intact and IPR-treatment are indicated by ∗∗ (p > 0.001) using Student's t-test.
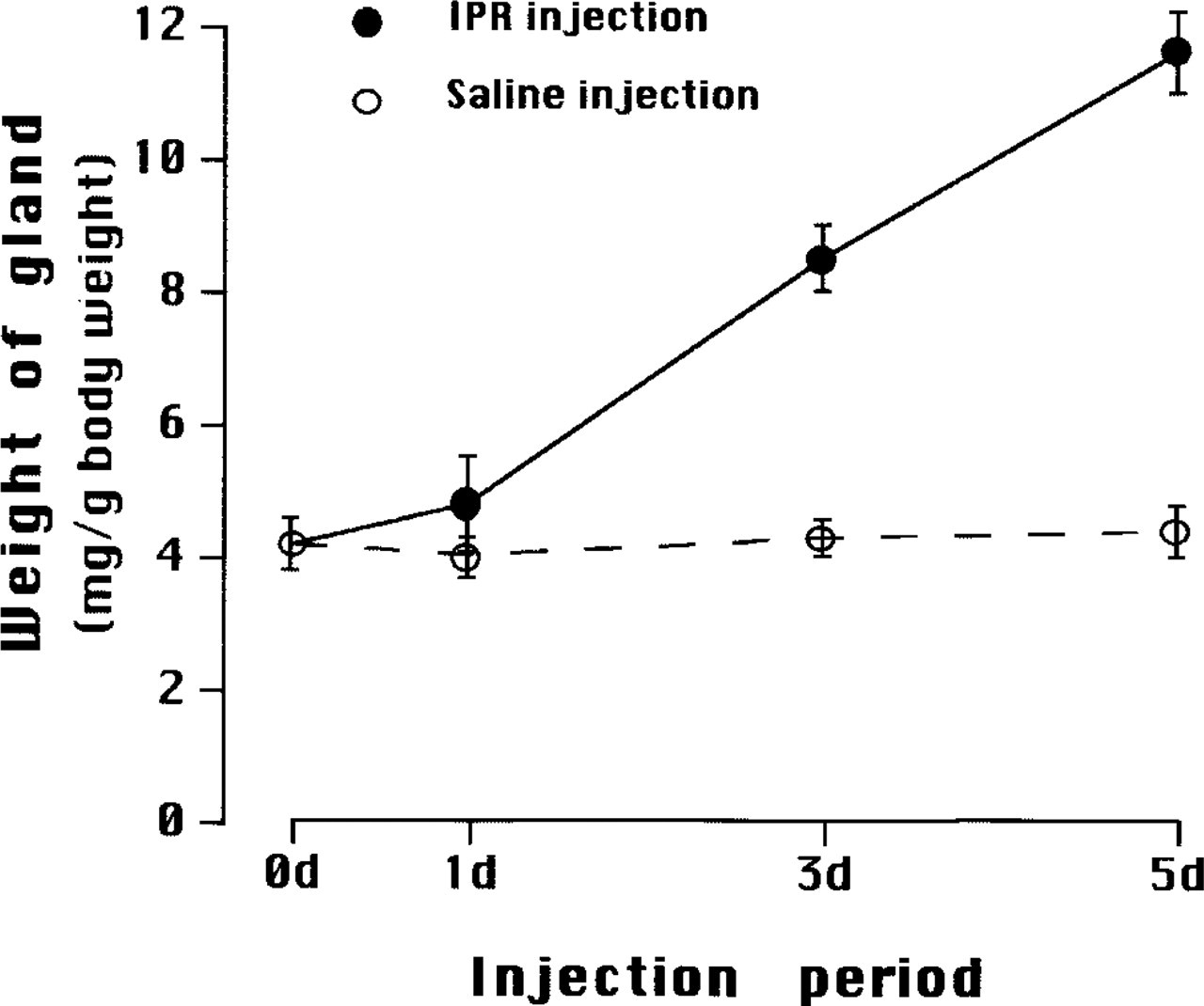
Changes in relative weight of SMGs of immature, 4-week-old mice during daily IPR injections. Intact mice are represented as 0 day. Control mice and experimental mice received daily saline (open circles) and IPR injections (solid circles) for up to 5 days (5 d). Mean values ± SD.
The structure of acinar cells in the SMGs of 4-week-old and adult female mice is similar. However, the glands of 4-week-old animals are still in a growing phase. During the experimental period, between 4 to 5 weeks of age, the absolute weight of SMGs of saline-injected control mice increased from 62.0 ± 0.6 mg to 80.5 ± 0.5 mg, but the relative weight was constant, about 4 mg/g bw. On the other hand, the relative weights of the SMGs of IPR-treated mice increased in proportion to the number of IPR injections; The average weight of SMGs of mice that received IPR injections for 5 days was about three times that of control (11.6 ± 0.6 mg/g bw) (Figure 4). The rate of increase was the same as that of adults.
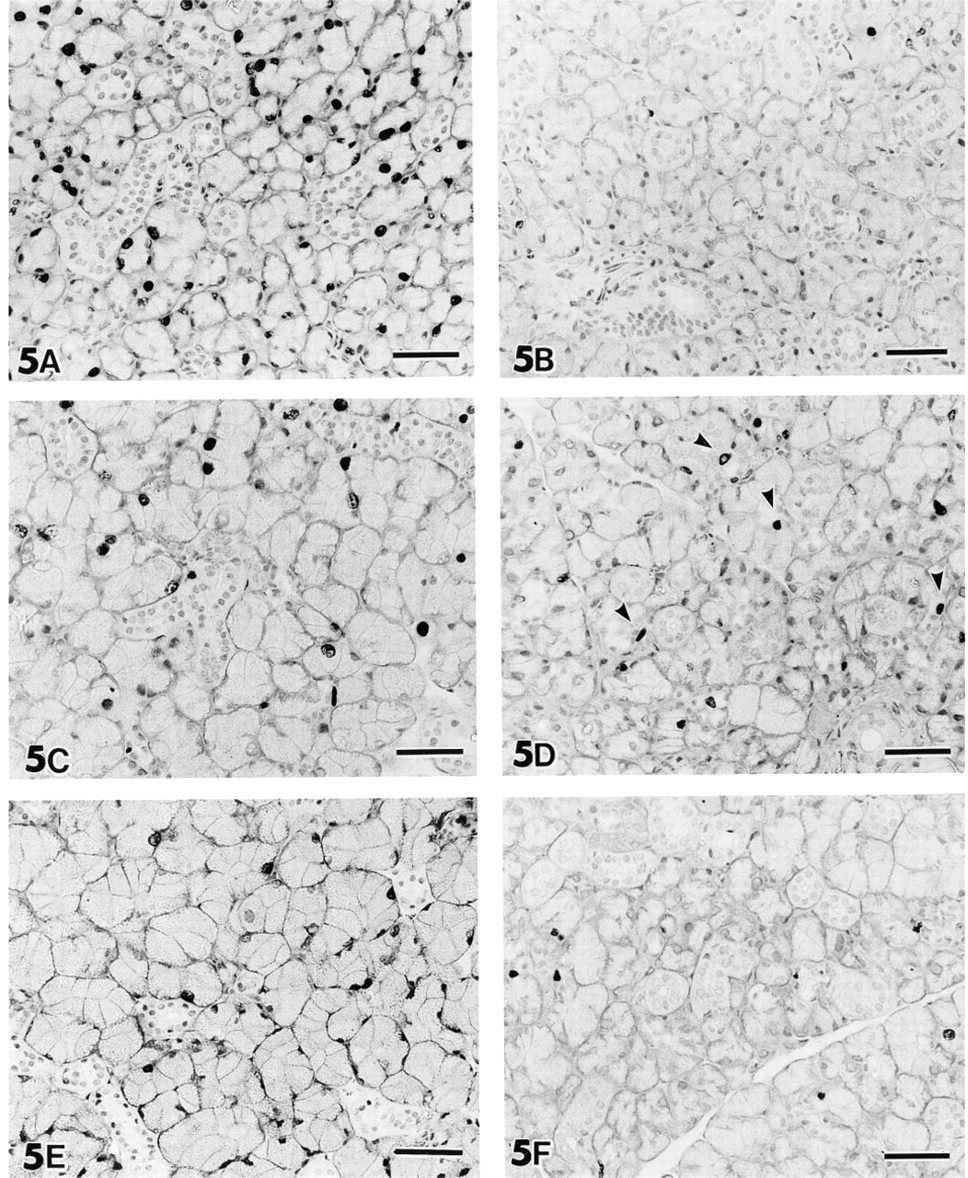
BrdU immunohistochemistry on SMG of 4-week-old mice. (
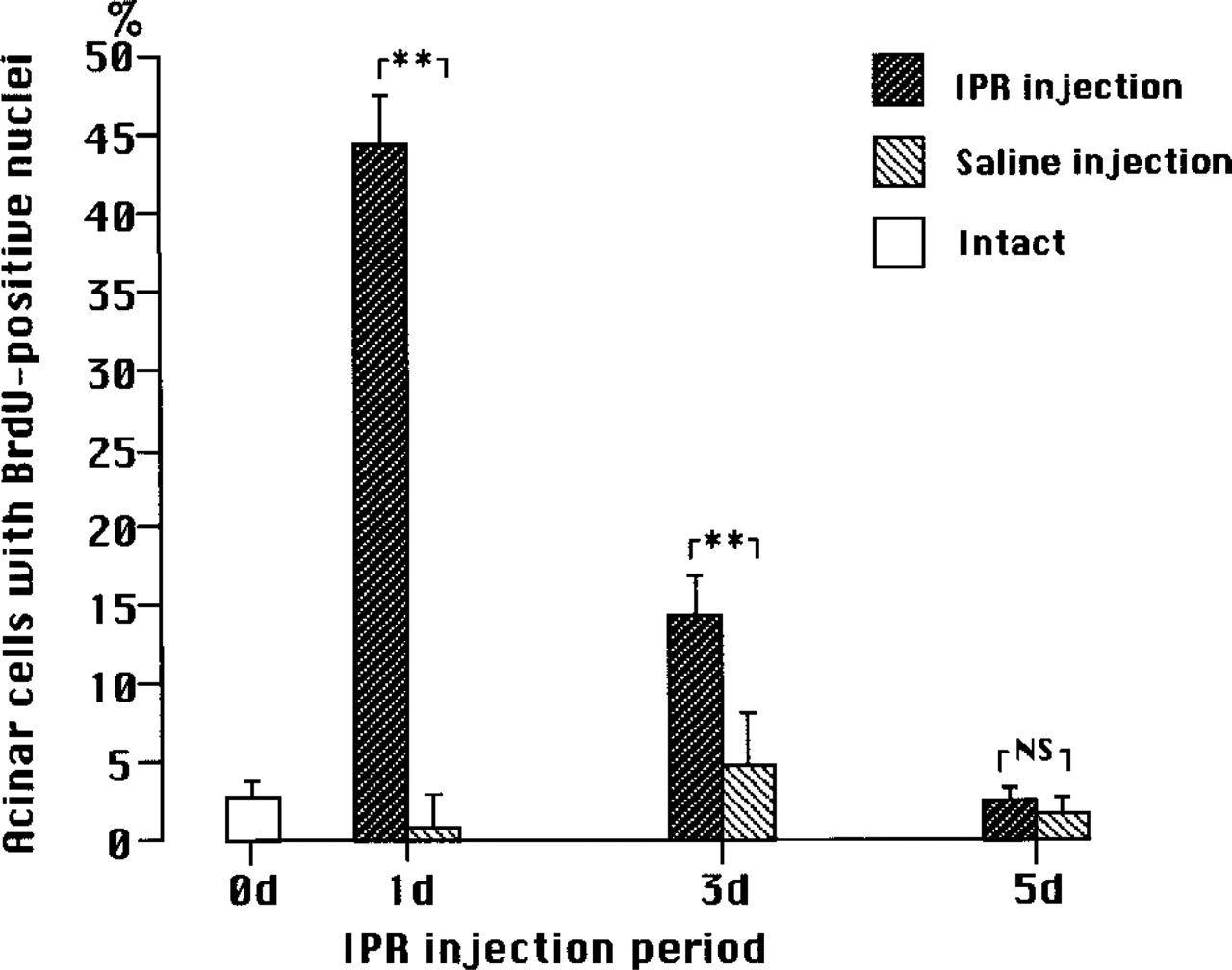
Changes in percentages of BrdU-incorporated nuclei in acini of SMG of 4-week-old mice during chronic administration of IPR or saline injections. Mean values ± SD. Less than 50% of the acinar cells have nuclei that have incorporated BrdU following one injection of IPR, and the percentage declines with increasing number of IPR injections. Significant differences were found between IPR and saline injection for 1 and 3 days (∗∗, p > 0.001) using Student's t-test, but the difference for 5 days was statistically not significant (NS).
Immunostaining for BrdU in the SMG of 4-week-old control mice that received saline injections showed only a few labeled acinar cells. However, some duct cells were stained, indicating a continued proliferative growth of the duct system (Figures 5B, 5D, and 5F). A single injection of IPR increased the number of nuclei immunoreactive for anti-BrdU (Figure 5A). IPR administration for 3 days resulted in fewer immunolabeled cells than were observed after one injection of the drug (Figure 5C). After IPR injections for 5 days there were even fewer labeled nuclei, almost the same as in the controls (Figure 5E).
The change in the percentage of BrdU-positive acinar nuclei in the glands of immature mice during chronic IPR stimulation was comparable to that found in the SMGs of adults (Figure 6). Contrary to our expectations, however, the percentage of BrdU-positive acinar nuclei induced by one injection of IPR in immature mice was less (44.5 ± 3.9%) than that in adults (52.5 ± 3.0%).
In a third study, we gave adult mice IPR every 48 hr for up to seven injections. We surmised that this regimen allows the cells to move into a new S-phase. Comparative data of gland weights and the percentages of BrdU-positive acinar cells between mice that received IPR injections at 24-hr intervals and those that were injected at 48-hr intervals are shown in Table 1. The relative weight of the SMGs of mice that received IPR at 48-hr intervals increased, depending on the number of IPR injections, up to five injections, but the rate of increase was moderate in comparison with that for mice injected every 24 hr. The percentage of BrdU-positive acinar cell nuclei in the gland of mice injected every 48 hr was higher than that of mice that received daily injections over the same period of time. However, this value was still less than half of that found after one injection of IPR, and the transient pattern was basically the same in the two groups. Thus, we confirmed that an increase in the interval between subsequent IPR injections did not lead to a continuous increase in DNA synthesis during chronic stimulation by the drug.
Changes in weight and percentage of BrdU-positive nuclei of SMGs treated with IPR at 24-hr and 48-hr intervals

bStatistical significance of difference (Student's t-test) between values of 24-hr- and 48-hr-injected mice; p > 0.005.
Discussion
The weight of the SMGs of adult female mice increased steadily with the number of daily IPR injections, which is consistent with the expected hyperplastic/hypertrophic effect of chronic IPR administration and which confirms the results of former studies (Brown–Grant 1961; Selye et al. 1961b; Schneyer 1962; Barka 1965b). In addition, the immunohistochemical studies with anti-BrdU antibodies on plastic-embedded sections confirmed that the IPR treatment induced the G0-phase acinar cells in the SMGs of mature mice to move into the S-phase and that such an induction was restricted to acinar cells during chronic stimulation. This suggests that duct cells, such as the intercalated duct (ICD) cells, which have been regarded to serve as stem cells for acinar cells, are not involved in the hyperplasia induced by IPR. It has been reported that in rats the ICD cells of the SMG and PAG incorporate [3H]-thymidine (Barka 1965b; van den Brenk et al. 1970) and undergo mitosis (Hand and Ho 1985) during chronic administration of IPR. Therefore, the lack of proliferation of ICD cells shown in this study may be specific to the mouse SMG. In the second study using 4-week-old mice, DNA synthesis was also noted in duct cells, including cells in the ICD and GCT, of both control and IPR-injected mice, without any difference in frequency of labeled nuclei. Differentiation of the duct system is taking place during this developmental stage in the mouse SMG, which is more protracted compared with that of acini (Gresik 1980). Therefore, DNA synthesis in the duct cells in the SMGs of immature mice is a physiological phenomenon.
We also observed that nuclei of acinar cells in the glands of mice treated with IPR, regardless of whether or not they incorporated BrdU, were less well-preserved after hydrolysis with HCl and digestion by protease, which are essential treatments before immunostaining with anti-BrdU, than those of duct cells and acinar cells of untreated animals. Stein and Baserga (1970) have reported that the acidic nuclear proteins may be involved in the control of cell proliferation in the IPR-stimulated salivary gland. It is likely that the nuclear protein components in the acinar cells changed during β-adrenergic stimulation, resulting in an altered chemical property of the nuclei.
During chronic IPR treatment the induction of DNA synthesis was transient. More than 50% of the acinar cells incorporated BrdU after one injection of IPR, and then the percentage rapidly decreased by 5 days of treatment and was the same as the control value after seven injections. There are some differences between the results in this study and that of Barka (1965b) for the induction pattern of DNA synthesis during chronic IPR stimulation of the rat SMG. His autoradiographic analysis of the rat SMG has shown that the peak of the incorporation of [3H]-thymidine into the acinar cells occurred after 2 days of IPR injection. His data, however, also have shown a lack of induction of DNA synthesis after five injections of IPR, as described in the present report.
It has been reported that the percentage of acinar cells engaged in DNA synthesis reaches its peak around 24 hr after a single injection of IPR (Barka 1965a; Radley et al. 1968). In addition, Radley et al. (1968) have shown that the cells in the rat PAG and SMG receiving multiple IPR injections enter S-phase more quickly than after a single injection. It is possible that we did not compare the percentages of labeled nuclei at the peak of response for each animal receiving IPR more than twice, when the frequency of cells in S-phase rapidly declines. However, Radley et al. (1968) have also shown that the peak value was significantly reduced after seven IPR injections, indicating that continuous stimulation of DNA synthesis was not maintained by repeated treatment with IPR, as was also observed in this study. Even so, it will be necessary to determine the peak time of DNA synthesis of acinar cells in the glands of mice receiving multiple injections of IPR.
Whereas the induction of DNA synthesis by chronic administration of IPR was transient, the gland weight increased in proportion to the number of IPR injections. The increased volume of acinar cells, which was noted after more than two injections of IPR, may reflect the increased number and size of secretory granules, as shown in a previous study of the rat SMG (Matsuura and Hand 1991). Chronic IPR administration causes an altered expression of secretory proteins. Proline-rich proteins (PRPs) (Ann and Carlson 1985) and cystatins (Barka et al. 1986b) are typical proteins induced in the SMG and parotid glands of rats and mice undergoing chronic IPR treatment, resulting in altered protein composition of the secretory granules of acinar cells (Matsuura and Hand 1991). Such an excessive protein synthesis and oversecretion could cause trophic changes during chronic stimulation, resulting in the enlargement of acinar cells.
The enlargement of the adult SMG by chronic IPR injection is mainly due to the enlargement (or hypertrophy) of acinar cells rather than to proliferation (or hyperplasia). The reason that the induction of DNA synthesis does not continue during chronic stimulation remains unknown. One possibility is that the secretory cells in the mature gland are postmitotic and therefore cannot sustain mitotic activity. Therefore, we have conducted a study with immature 4-week-old mice. At this developmental stage, the gland is still increasing in size and it was anticipated that the cells still have the potential to proliferate more than those in the mature SMG. However, the patterns of response to IPR administration were similar in immature and adult mice. In addition, the induction of DNA synthesis by IPR in 4-week-old mice was less than that in adults. It has been reported that in rats IPR had a greater effect on the salivary glands of 5-week-old animals than on those of 4-week-old ones (Klein 1984) and that IPR stimulation of DNA synthesis is age dependent (Barka et al. 1973). Malamud and Baserga (1967) have described that local injection of IPR into the rat SMG is more effective in stimulating DNA synthesis than systemic administration, suggesting that IPR acts directly through receptors on the salivary gland cells in inducing DNA synthesis. Little is known about the development of the β-adrenergic receptors in the salivary gland cells, either in terms of the number or binding affinity for the ligand. The rate of increase in SMG weight in immature mice during chronic IPR treatment was similar to that in the adults, indicating that β-adrenergic receptors are mature enough to mediate a continuous hypertrophy, as in adults. Therefore, the lesser effect of IPR on the immature SMG compared to that observed in adult appears to be specific for the induction of DNA synthesis, suggesting that the pathway that leads to the induction of DNA synthesis after the binding of the ligand to the receptor may still be immature at this developmental stage.
It is possible that the 24-hr interval between injections of IPR, as in the first study, did not provide sufficient recovery time to allow the cells to reenter a new S-phase. The rate of increase in SMG weight of mice treated at 48-hr intervals was moderate in comparison with that for those injected every 24 hr, but the percentages of DNA-synthesizing cells were about twice those of mice receiving daily injections for the same total number of injections. However, these percentage values for BrdU-positive cells were still less than half of those found after one injection of IPR, and the transient stimulation of DNA synthesis was basically the same. This indicates that the extended interval between IPR injections contributes only in a minor way to continued proliferation with repeated IPR treatment.
Tsang et al. (1980) have proposed a role for an elevated level of cAMP, the intracellular messenger of IPR, in the induction of DNA synthesis by IPR in the rat PAG. They reported two peaks of elevated cAMP after a single injection of IPR and claimed that the second elevation, detected 12 hr after injection, was essential to the initiation of DNA synthesis. There is therefore the possibility that the pattern of cAMP level was changed after multiple IPR injections. In a preliminary experiment, the cAMP levels after one or two injections of IPR were measured by the courtesy of Dr. Sugiya (Nihon Univ. Matsudo Dental School, Japan). However, we did not find a second elevation of cAMP after the first surge for mice injected either once or twice with IPR, and found no pattern of change in cAMP level that would account for the fact that the DNA synthesis did not occur after multiple IPR injections.
On the basis of our results, we propose that the regulatory systems in the acinar cells of the mouse SMG for hypertrophy and hyperplasia are different. The transient induction of DNA synthesis by IPR is an essentially physiological response unrelated to the proliferation seen in malignant transformation. A future study will focus on the branching point in the intracellular transduction pathways that leads to different responses.
Footnotes
Acknowledgements
Supported in part by Grant-in-Aid 04671112 (SM) from the Ministry of Education, Science and Culture of Japan.
We thank Dr Hiroshi Sugiya (Nihon Univ. Matsudo Dental School) for his work on the measurement of cAMP levels.