
Abstract
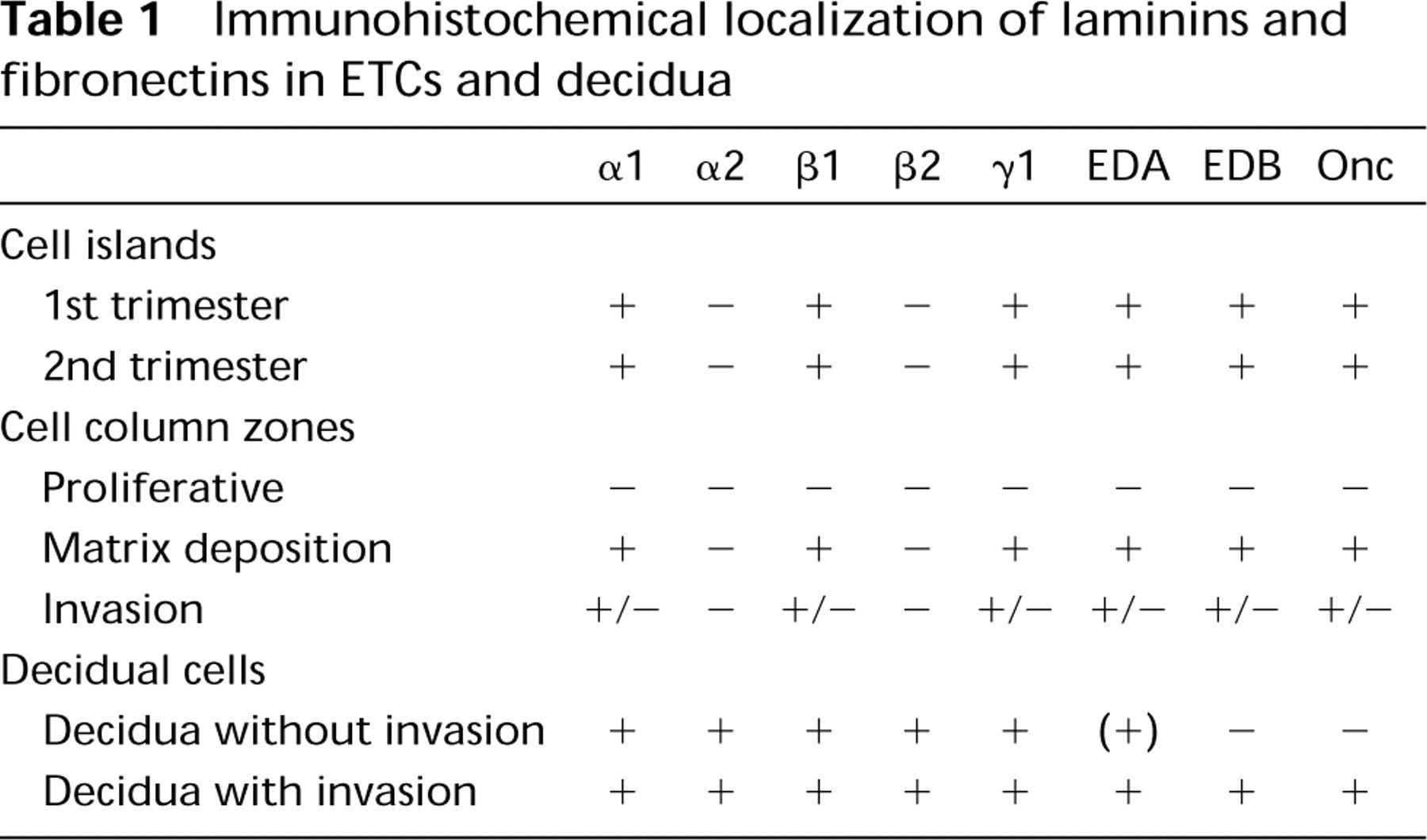
We studied the distribution of laminin (Ln) α1–α3, β1–β3, and γ1 chains, and of the extradomain-A (EDA) and EDB and the oncofetal epitope of fibronectin (Onc-Fn) in extravillous trophoblastic cells and decidua in the human placenta by immunohistochemistry. We found that the transition from villous to extravillous trophoblast was accompanied by emergence of immunoreactivity for EDA-, EDB-, and Onc-Fn among the cells. Furthermore, whereas the villous trophoblastic basement membrane (BM) contains Ln α1, α2, β1, β2, and γ1 chains, immunoreactivity for Ln α1, β1, and γ1, but not for Ln α2 and β2 chains, was detected in association with extravillous trophoblastic cells. Interestingly, although immunoreactivity for the Ln α1, α2, β1, β2, and γ1 chains was detected in all decidual cell BMs, EDB-Fn and Onc-Fn were detected only in decidua that had been invaded by the trophoblast. In summary, our results describe distinct changes in the distribution of Ln and Fn isoforms during the differentiation of villous trophoblast into extravillous trophoblastic cells. Furthermore, EDB- and Onc-Fn are preferentially found in decidua that has been invaded by the trophoblast, indicating that the deposition of these Fn isoforms reflects a decidual cell response to invasion.
Keywords
The Cyto- and syncytiotrophoblast form the epithelial covering of fetal chorionic villi. The villous cytotrophoblastic cells are capable of fusing to form syncytiotrophoblasts that comprise the outer cellular layer covering the villi. The extravillous trophoblastic cells (ETCs) represent another unique direction of cytotrophoblastic cell differentiation (Benirschke and Kaufmann 1990; Yeh and Kurman 1989). During placentation, the ETCs leave the villous surface and can be found as cell islands in intervillous spaces, in anchoring cell columns, or as single cells that have invaded into the decidua, decidual septae, or chorionic plate. The molecular events guiding the differentiation of ETCs are largely unknown.
Laminins (Lns) are a family of extracellular matrix glycoproteins that are important basement membrane (BM) constituents. They are heterotrimers, consisting of an α-, a β-, and a γ-type chain. Several isoforms have been described for each chain type (Timpl and Brown 1994). Fibronectins (Fns) are ubiquitous components of extracellular matrices (ECMs). Several variants are produced by alternative splicing of exons encoding for the extradomain A (EDA), EDB, and type III connecting segment (IIICS) domains (Ffrench-Constant 1995). Another source of immunological heterogeneity of Fns is glycosylation. The MAb FDC-6 specifically recognizes an alternatively glycosylated epitope, called the oncofetal epitope, within the IIICS domain (Matsuura et al. 1988; Matsuura and Hakomori 1985).
Immunolocalization studies have revealed that Ln is found in villous BMs (Charpin et al. 1985; Earl et al. 1990). However, as the trophoblast differentiates into ETCs in trophoblastic columns, there are changes in the distribution of Ln and Fn isoforms. Three immunohistochemically defined zones within the anchoring cell columns represent different stages of ETC differentiation (Frank et al. 1994; Damsky et al. 1992). The most proximal cell layers of the column (the proliferative zone) contain proliferating cytotrophoblastic cells, which synthesize few or no ECM molecules. The more distal parts of the column contain few proliferative cells, but there is abundant deposition of Fn, Ln and collagen Type IV (matrix deposition zone). The most distal cells of the column invade the decidua (invasion zone).
Recently, we found that the invasion of the uterine wall by the ETCs is accompanied by a change in integrin expression of the invading cells. When villous trophoblastic cells differentiate into ETCs in anchoring columns, the cells cease to express the integrin α6β4, which is a Ln receptor, and begin to express the α1β1 and α5β1 integrins, which bind Ln and Fn, respectively (Damsky et al 1992,1994; Aplin 1993; Korhonen et al. 1991). We have also studied earlier the distribution of EDA-Fn in the developing and term placenta (Virtanen et al. 1988). Here we have extended these immunohistochemical studies to the distribution of the various Ln and Fn isoforms in human ETCs and decidua using monoclonal antibodies (MAbs).
Materials and Methods
Tissues
A total of 16 human placentas (seven 8–10 weeks, five 12–20 weeks, and four 38–40 weeks of gestation) were acquired from spontaneous abortions due to rupture of fetal membranes, from legal abortions performed for psychosocial indications, or from normal deliveries (Jorvi Hospital; Espoo, Finland). The tissues were frozen in liquid nitrogen and stored at −74C until use. Routine hematoxylin-eosin staining was used to establish the normal histology of the tissue samples.
Immunoprecipitation and Western Blotting
For SDS-PAGE and Western blotting, whole placental tissue was extracted in electrophoresis sample buffer (Laemmli 1970). For enrichment of placental ECM, placental tissue was extracted with 0.5% sodium desoxycholate (DOC; Sigma, St Louis, MO) in 50 mM Tris-HCl (pH 7.4) at 0C for 30 min, followed by washes in 10 mM Tris-HCl (pH 7.4) at 0C three times in the presence of protease inhibitors (Hedman et al. 1979). The samples were run in a 6.5% polyacrylamide electrophoresis gel in a reducing buffer (Laemmli 1970), and transferred onto nitrocellulose membranes (Towbin et al. 1979). The membranes were blocked with 3% bovine serum albumin-PBS overnight, incubated with the MAbs for 60 min, and exposed to peroxidase-coupled rabbit anti-mouse IgG antiserum (Dako; Glostrup, Denmark). The bound antibodies were visualized by the 3,3-diaminobenzidine tetrahydrochloride reaction (DAB; Sigma).
Antibodies
MAbs and antisera to Lns and Fns were as follows: α1 (clone 4C7; Engvall et al. 1986, 1990); α2 (clone 5H2; Leivo and Engvall 1988); α3 (clone BM-2; Rousselle et al. 1991); β1 (clone 4E10; Engvall et al. 1990; Wewer et al., 1983); β2 (clone C4; Hunter et al. 1989); β3 (clone 6F12; Marinkovich et al. 1992); γ1 (clone 2E8; Engvall et al. 1986,1990); 425, a rabbit antiserum to Ln (Liesi et al. 1983); all Fn isoforms (clone 52BF12; Vartio et al. 1987); EDA-Fn (clone 52DH1; Vartio et al. 1987); EDB-Fn (clone BC-1; Carnemolla et al. 1989) and Onc-Fn (clone FDC-6; Matsuura and Hakomori 1985). All of these have been characterized earlier. We have recently raised and characterized a mouse MAb that detects the human Ln β1 chain in cryostat tissue sections as well as in Western blots (LnB1; Virtanen et al., submitted for publication). Anti-cytokeratin 19 serum (Leoncini et al. 1990) and the MAb 2A4 recognizing cytokeratins 8, 18, and 19 (Virtanen et al. 1985) were used to identify trophoblastic cells in double immunostainings and serial sections, respectively. The MAb Ki-67 was used to identify proliferating cells (Gerdes et al. 1983). Polyclonal antisera to desmin (Virtanen et al 1986; Badley et al. 1978) and vimentin (Virtanen et al. 1981) were used to identify cells in the villous stroma. The optimal dilutions of the antibodies were determined by using a panel of human normal tissues and cultured cells.
The designation of the epitopes recognized by the various Fn antibodies requires comment. The MAbs 52DH1 and BC-1 used in this study recognize EDA- and EDB-Fns, respectively (Carnemolla et al. 1989; Vartio et al. 1987). The MAb FDC-6 reacts with an alternatively glycosylated epitope within the IIICS segment, which is frequently found in fetal and malignant tissues and was designated the “oncofetal Fn epitope” (Matsuura et al. 1988; Matsuura and Hakomori, 1985). Subsequently EDB-Fn has also been called “oncofetal Fn” in some studies (Frank et al. 1994). In this study, however, Fn containing the FDC-6 epitope is called “oncofetal Fn (Onc-Fn),” and the epitope recognized by the MAb BC-1 is called the EDB domain.
Immunohistochemistry
Frozen sections were cut at 5 μm and fixed in acetone at −20C for 10 min. For indirect immunofluorescence microscopy, the sections were exposed to the primary and subsequently to the secondary antibodies (fluorescein isothiocyanate-coupled goat anti-mouse IgG and tetramethylrhodamine-coupled goat anti-rabbit IgG sera; Jackson Immunoresearch, West Grove, PA) at room temperature (RT) for 30 min. For negative controls, sections were exposed only to the secondary antisera. The specimens were embedded in sodium veronal/glycerol buffer (1:1; pH 8.4) and examined with a Leitz Aristoplan microscope equipped with appropriate filters. For light microscopy, sections were exposed to the primary antibodies at RT for 30 min. The bound antibodies were visualized with the APAAP (alkaline phosphatase-anti-alkaline phosphatase) method (Dako). Endogenous alkaline phosphatase was blocked with 1 M levamisole (Sigma). Negative controls were carried out for each set of serial sections by omitting the primary antibodies. The controls revealed minor unspecific staining in certain areas of the tissue sections; these were excluded from the study. The sections were counterstained with Mayer's hematoxylin (Merck; Darmstadt, Germany) and mounted in GVA mounting medium (Zymed; San Francisco, CA).
Immunohistochemical localization of laminins and fibronectins in ETCs and decidua
Placental Histology and Nomenclature
The distribution of the various Ln and Fn isoforms was evaluated by indirect immunofluorescence as well as light microscopy from sets of serial sections of the placental samples. Histology of the immunofluorescence sections was evaluated by light microscopy of adjacent sections. We have used descriptive terms such as “weak” and “strong” to denote the intensity of the immunoreaction compared to other areas within the same microscopic section. Other terms such as “sparse” or “specks” vs “rich” or “extensive” immunoreactivity were used to describe the distribution of the immunoreactivity. Nomenclature for placental structures has been adopted from Benirschke and Kaufmann (1990) and Castellucci et al. (1990). The zones of anchoring cell columns (proliferative, matrix deposition, and invasion zones; see above) are adapted from Damsky et al. (1992) and Frank et al. (1994).
Results
The immunolocalization results are summarized in Table 1.
Specificity of the β2 MAb in Human Placental Tissue
MAb C4 recognizes the Ln β2 chain in several species, including human. However, weak crossreactivity of the MAb with the Ln β1 chain and an unidentified 400-kD protein has been reported in rat protein extracts. We used Western blotting of whole human second-trimester placental lysates and desoxycholate extracts of human placentas to define the specificity MAb C4 (Figure 1). These results demonstrate that it recognizes a single 190-kD band corresponding to the Ln β2 chain but does not react with the β1 chain or a higher molecular weight band in the human placenta.
ETCs in Cell Islands
Trophoblastic cell islands of the first trimester placenta are frequently cellular, with little extracellular material. Antibodies to Ln α1 (Figure 2c), β1 (Figure 2a), and γ1 chains revealed small specks of immunoreactivity in the midst of the trophoblastic cells, whereas no immunoreactivity for the Ln α2 (Figure 2e) or β2
Western blotting of whole human second trimester placental lysates (Lanes 1–3) and preparation from the same tissue enriched for extracellular matrix material by desoxycholate treatment (Lanes 4–6). Lanes 1 and 4, total protein profile of the samples as revealed by Amido black staining. Lanes 2 and 5, immunoblotting with LnB1, an Ln β1 chain MAb. Lanes 3 and 6, the corresponding immunoreactions with MAb C4 to the Ln β2 chain. Note that there is no reaction at the level of the Ln β1 chain band with C4 and that there are no reactive higher molecular weight bands with either of the antibodies. The faint smaller molecular weight bands most probably correspond to degradation products of Ln.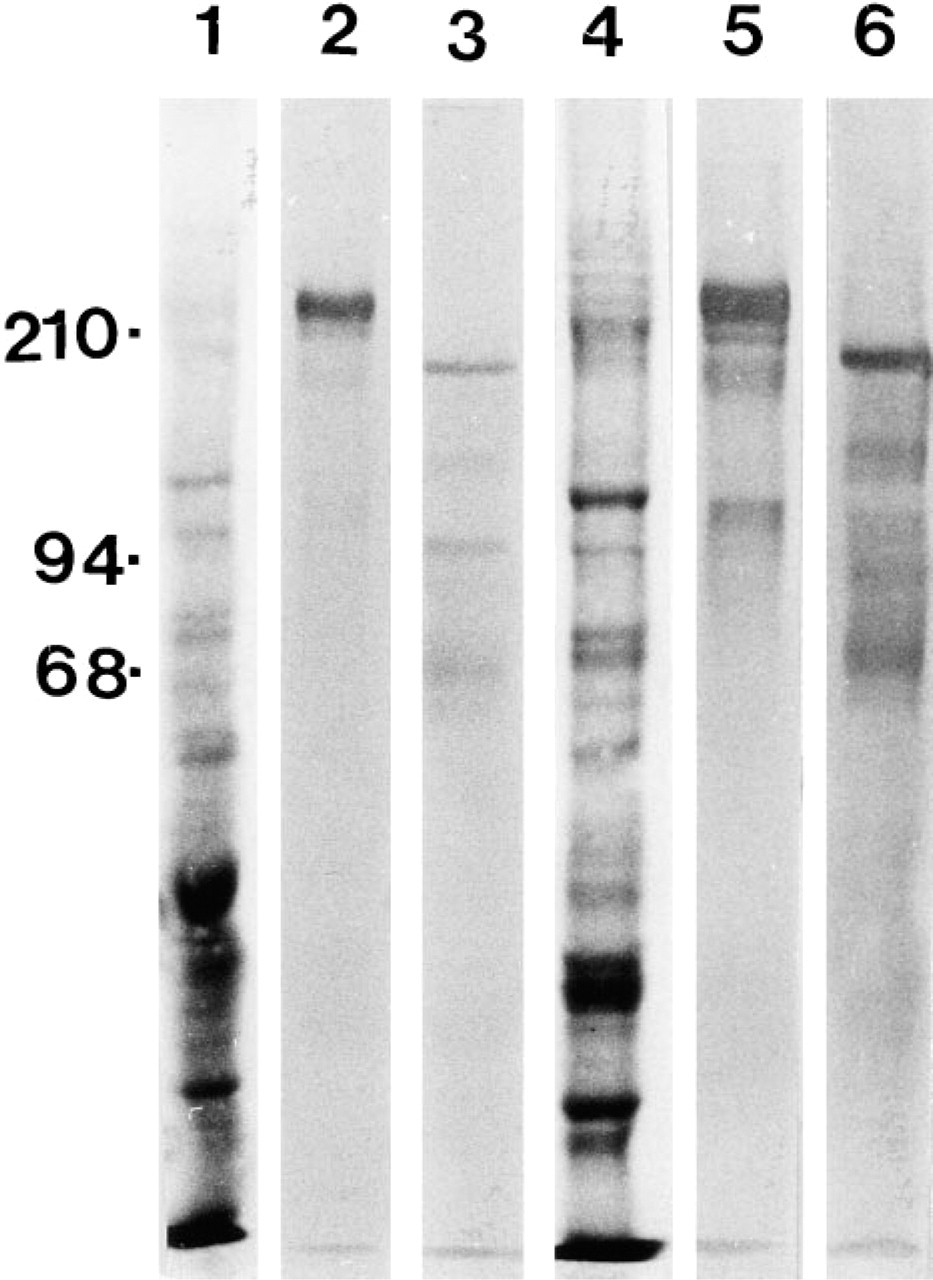
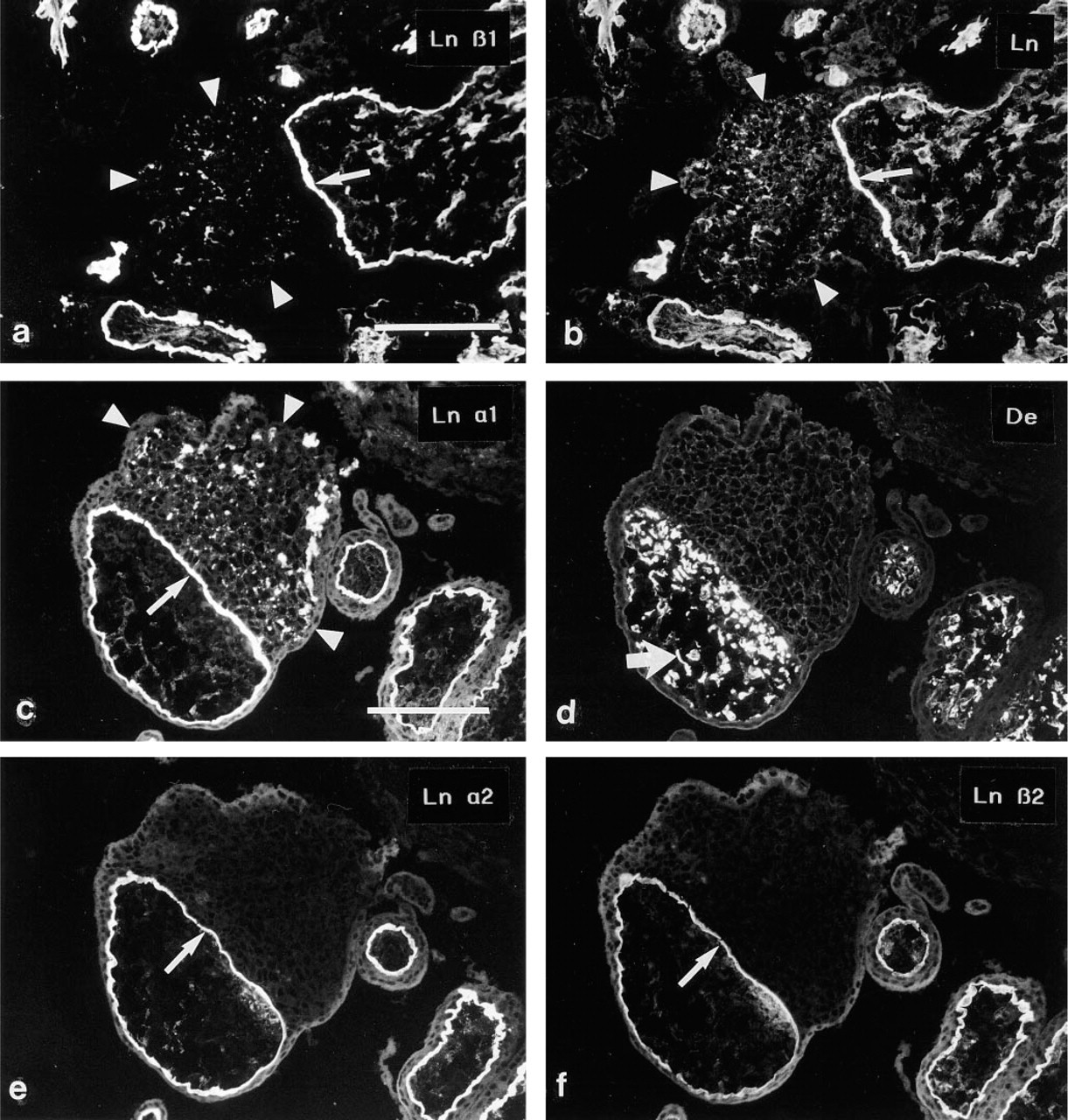
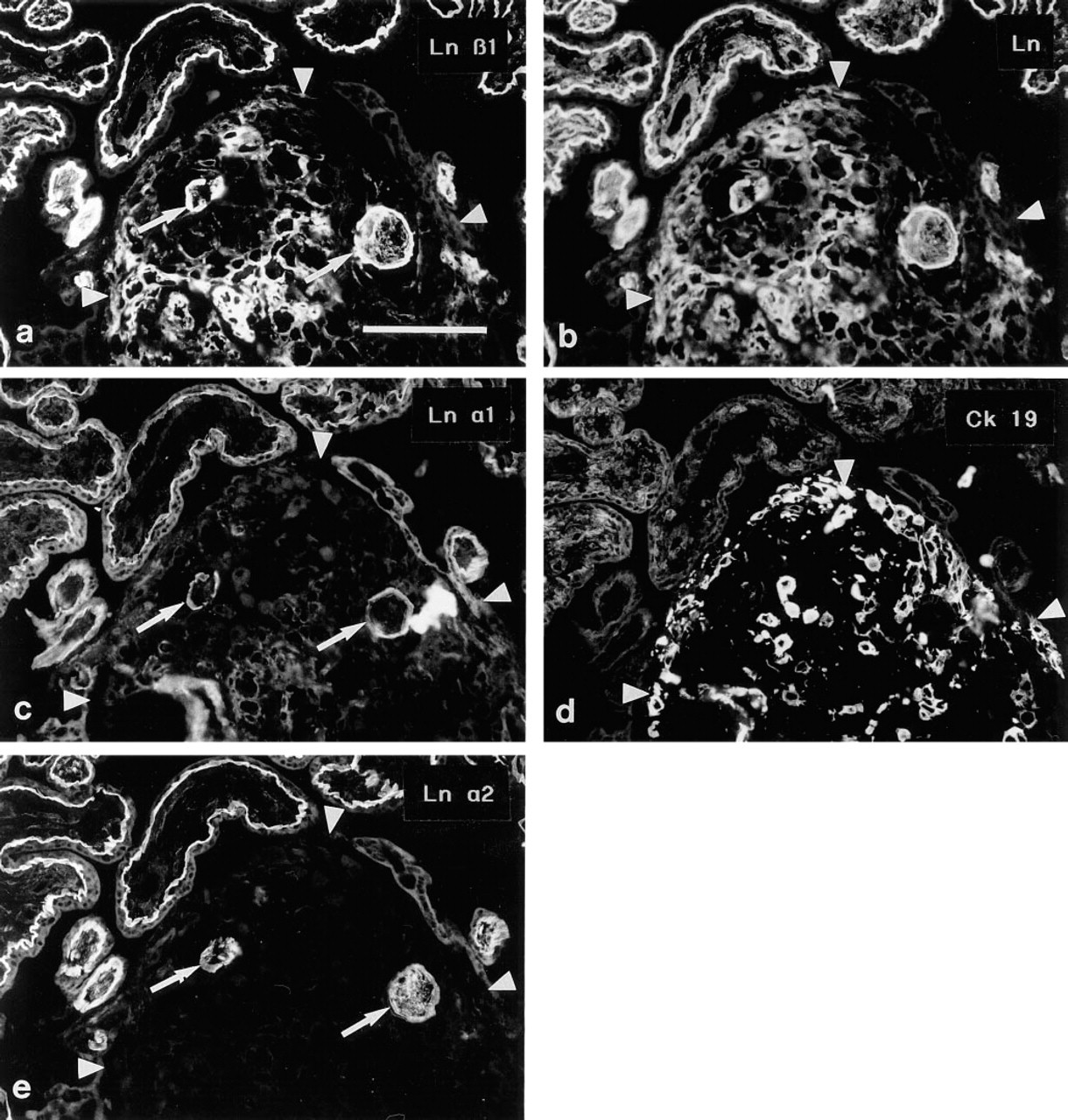
(a,b) Double immunostaining of a cell island from a 10-week placenta. The Ln β1 chain MAb (
During the second and third trimesters of placental development, cell islands characteristically consist of ETCs embedded within fibrinoid material. Distinct Ln β1 chain immunoreactivity, co-extensive with that seen with the polyclonal Ln antiserum, was detected among the fibrinoid material within the cell island (Figures 4a and 4b). A similar, albeit less intensive, immunoreactivity was detected for the Ln α1 (Figures 4c and 4d) and γ1 chains, whereas the Ln α2 (Figures 4e and 4f) and β2 antibodies did not react with the fibrinoid material. As in cell islands without fibrinoid, abundant immunoreactivity for EDA-, EDB-, and Onc-Fns was detected within the fibrinoid (not shown).
Serial sections of a cell island from a 10-week placenta immunostained for the Ln β1 chain (
ETCs in Anchoring Cell Columns
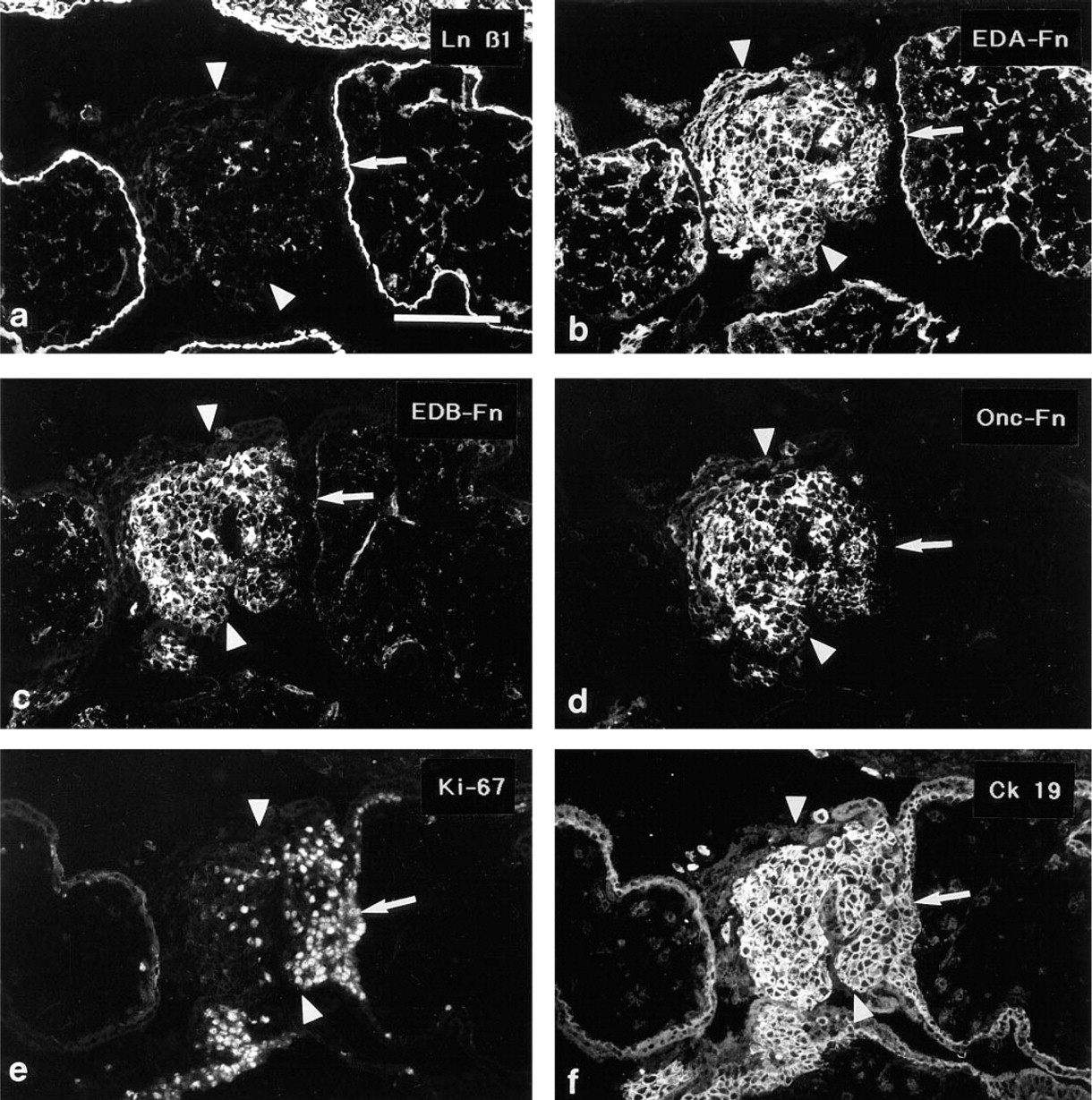
The Ln α1, α2, β1, β2 and γ1 MAbs all reacted with the villous trophoblastic basement membranes at the villus-column interphase. The nuclei of the basal first to fifth rows of cells adjacent to the villous BM reacted acted intensively with the proliferation marker Ki-67 (proliferative zone; not shown). The Ln and Fn antibodies did not react with these most basal cell rows in the columns. In the proximal part of the matrix deposition zone, small specks of Ln α1, β1 (Figures 5a and b) and γ1 chain immunoreactivity were seen. ETCs that were located more distally in the columns (matrix deposition zone) were surrounded by distinct linear Ln α1, β1 (Figures 5a and 5b) and γ1 chain immunoreactivities, and extensive immunoreactivity for EDA-, EDB-, and Onc-Fns (Figures 5e and 5f). The Ln α2 (Figures 5c and 5d) and β2 chains were not detected among ETCs in anchoring cell columns.
Serial sections of a trophoblastic cell island in which the ETCs are enmeshed within fibrinoid from a 16-week placenta. Cross-sections of freely floating villi are seen in the upper part of the micrographs. Arrowheads delineate the cell island; arrows identify two villi that are enveloped within the fibrinoid material of the island. Double immunostaining with the Ln β1 chain MAb (
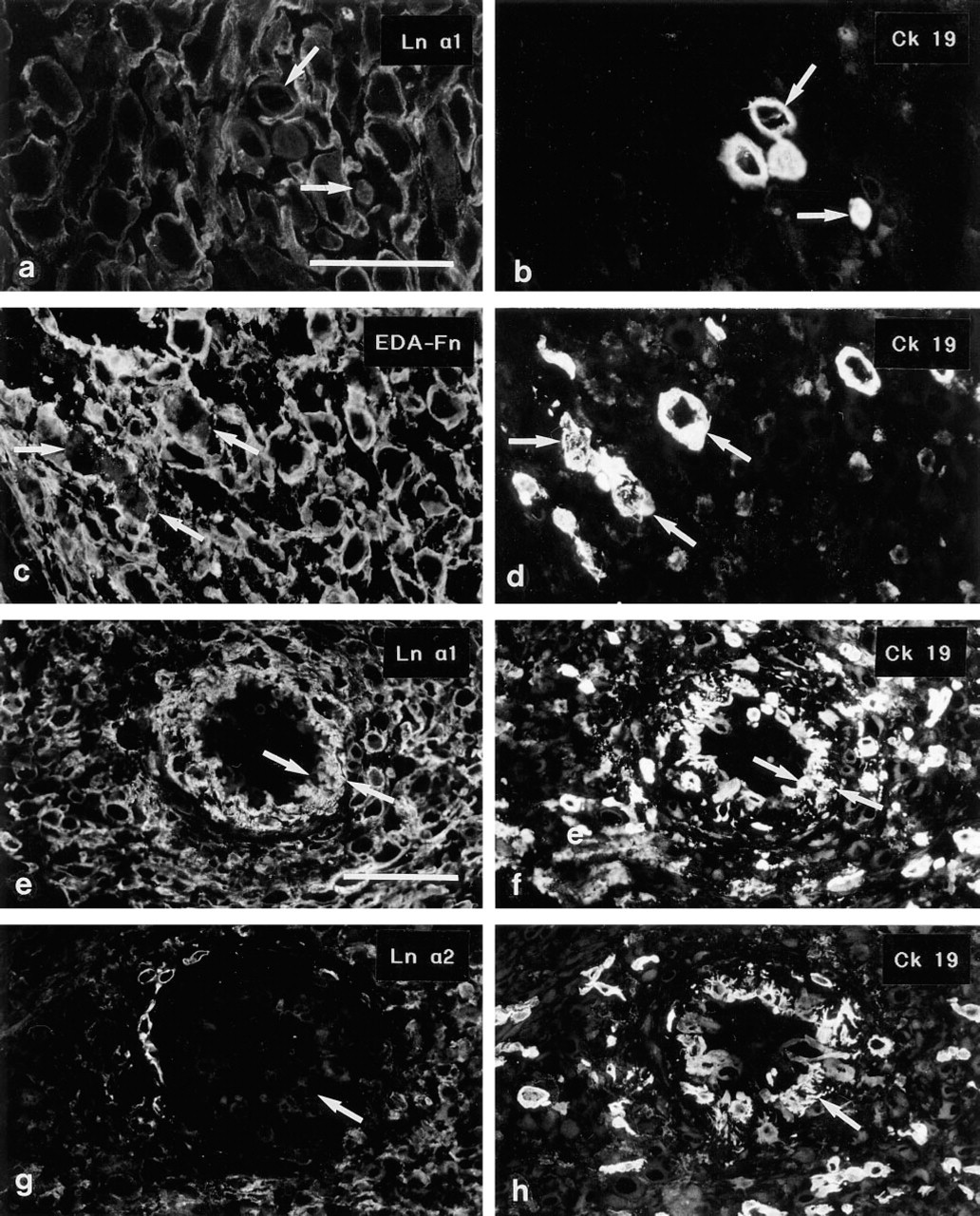
Double immunostainings using the polyclonal anti-cytokeratin serum enabled us to identify individual ETCs that had invaded further into the placental plate (invasion zone). These cells reacted heterogeneously with the Ln antibodies: a part of the cells were devoid of cell-associated Ln chain immunoreactivity, whereas some cells presented weak reactivity for the Ln α1 (Figures 6a and 6b) β1 and γ1 chains. MAbs to the Ln α2 and β2 chains reacted weakly at the surface of a small minority of the ETCs that were found among the decidual cells. Similarly, immunoreactivity for the Fn isoforms was variable, with some cells displaying distinct pericellular immunoreactivity whereas other cells appeared negative (Figures 6c and 6d, displaying EDA-Fn). Only occasionally did these cells display nuclear positivity for MAb Ki-67 (not shown).
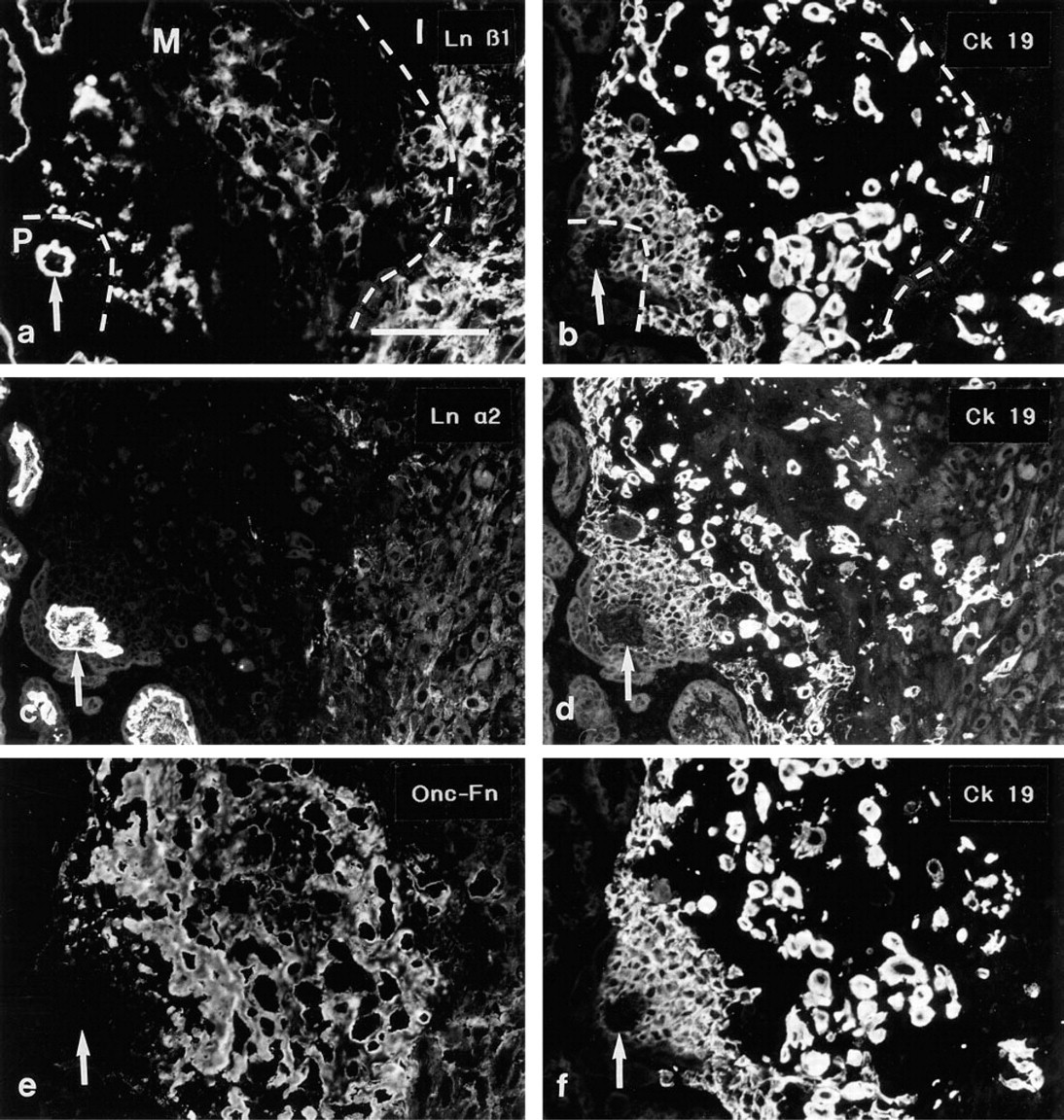
Serial sections of an anchoring cell column from a 16-week placenta immunostained for the Ln β1 (
(
(
ETCs within the walls of spiral arteries reacted with MAbs to Ln α1 (Figures 6e and 6f), β1 and γ1 chains, but not with MAbs to α2 (Figures 6g and 6h) or β2. MAbs against the Fn isoforms reacted distinctly with these ETCs (not shown).
Decidual Cells
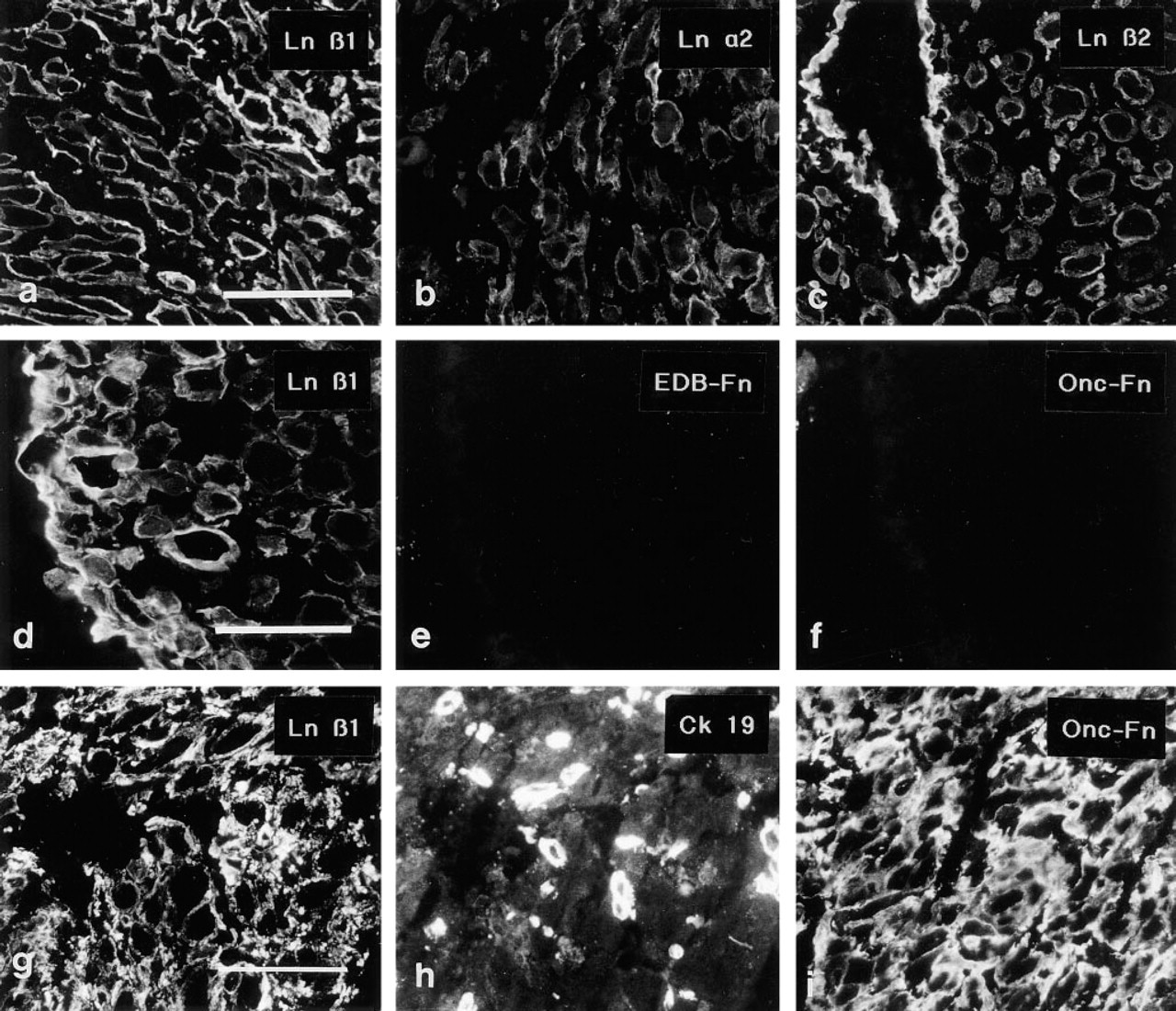
MAbs to the Ln α1, α2 (Figure 7b), β1 (Figure 7a), β2 (Figure 7c) and γ1 chains revealed linear pericellular immunoreactivity around the decidual cells. The immunoreactivity co-aligned with that of a polyclonal anti-Ln serum not shown), surrounding the individual cells in a basement membrane-like fashion.
Interesting differences regarding the distribution of Fns were noted in decidual tissue. In decidual tissue that was not invaded by ETCs, the MAb recognizing the EDA-Fn reacted only weakly and the MAbs to EDB-Fn (Figure 7e) and ONC-Fn (Figure 7f) did not react with decidual cell basement membranes. In samples in which invasion by ETCs was detected by double immunostaining with cytokeratin 19 antiserum, intensive immunoreactivity for EDA-, EDB-, and ONC-Fns (Figure 7i) was seen in decidual cell BMs and the intercellular matrix. One of the tissue samples from a 10-week placenta contained both areas with and areas without ETC invasion; the same differential distribution of Fn epitopes was detected in the two areas. In contrast to the differences in Fn distribution, the anti-Ln 2α, β1 (Figures 6d and 6g), and β2 MAbs reacted with decidual cell BMs in all decidual samples. The MAb recognizing Ln α1 reacted weakly but detectably with decidual tissue without ETC invasion, whereas more distinct (Figures 6a and 6b) immunoreactivity was detected for this Ln chain in samples with invasion.
The Ln α3 and β3 chains, components of Ln-5, Ln-6 and Ln-7, were not detected in ETCs or the decidua (not shown).
Discussion
The distribution of Ln and Fn isoforms in the extravillous trophoblast is significant in two respects. Altered expression of receptors for extracellular matrix components, including receptors for Lns and Fns, has been shown to accompany trophoblast detachment from the villous BM and subsequent invasion into the placental bed (Aplin 1993; Damsky et al. 1992; Korhonen et al. 1991). Second, it is becoming apparent that the ETC represents a distinct direction of trophoblast differentiation (Benirschke and Kaufmann 1990; Yeh and Kurman 1989). It is therefore of interest to determine whether the expression of Lns or Fns is altered on differentiation in the extravillous direction.
Although MAb C4 has been widely used to detect the Ln β2 chain in tissue sections, Sanes et al. (1990) reported weak crossreactivity with the β1 chain in rat protein extracts, and also reactivity with an undefined 400-kD protein (Hunter et al. 1989). In this work we showed by immunoblotting that C4 recognizes only the Ln β2 chain in the human placenta.
The MAb 4C7 does not recognize denatured laminin in Western blots, and its specificity has been shown indirectly. It binds to the G-domain of a Ln α-type chain (Engvall et al. 1986); it immunoprecipitates a trimeric laminin molecule from JAR cells known to produce the α1 chain (Haaparanta et al. 1991). The epitope is immunologically distinct from Ln α2 (Engvall et al. 1990), and the molecular weights of the Ln α3-α5 chains are distinct from the heavy chain recognized by the MAb 4C7 (Engvall and Wewer 1996). The MAb 4C7 has been used in this and a number of other studies to localize the α1 chain, but confirmation of its epitope will require the characterization of other α1-specific antibodies. To our knowledge, no such MAbs are available at present.
Decidual Response to Invasion
Decidualization is accompanied by the deposition of a BM around each individual decidual cell. This BM has been shown to consist of Ln, Fn, Type IV collagen, and heparan sulfate proteoglycan (Charpin et al. 1985; Wewer et al. 1985). Our results show that the decidual cell BM contains the Ln α1, α2, β1, β2, and γ1 chains. Interestingly, increased immunoreactivity for EDA-Fn and emergence of the EDB- and Onc-Fn epitopes were detected only in decidual tissue that was invaded by ETCs. These results suggest that deposition of these Fn epitopes in decidual cell BMs is a response to invasion by the ETC and is distinct from decidualization itself. Deposition of alternatively spliced Fn isoforms is likewise found in the stroma of malignant tumors (Hauptmann et al. 1995; Iozzo 1995).
Church et al. (1996) reported that Ln α2, β1, β2, and γ1 chains are found in decidual cell BMs, but that only minor amounts of the α1 chain are present. In our samples, the immunoreaction for the α1 chain was indeed weak in decidual tissue without ETC invasion but was distinct around decidual cells of invaded tissue. Church et al. (1996) did not address the issue of invasion, which may explain the partial discrepancy of our results with theirs.
The Extracellular Matrix of Cell Islands and Columns
Trophoblastic cell islands of the first trimester (8–10 weeks in our material) were often cellular, with only a limited amount of extracellular material. On the other hand, some first-trimester and most of the second- and third-trimester cell islands consisted of ECTs enmeshed in fibrinoid material. The extracellular matrix in these structures has been termed “matrix-type fibrinoid” (Frank et al. 1994). It contains Fns (including the EDA and EDB domains), Ln, collagen Type IV, and tenascin (Frank et al. 1994; Nanaev et al. 1993). The results of this study show that the matrix of both types of cell islands contains Ln α1, β1, and γ1 chains (components of Ln-1), whereas the Ln α2 and β2 chains were not detected. The matrix was also strongly immunoreactive for EDA-, EDB-, and Onc-Fn.
Recently, a gradient of Ln and Fn chain distribution along the proximal-distal axis of cell columns has been described (Frank et al 1994; Damsky et al 1992). Our results are in agreement with and further define this model. The basal cell layer adjacent to the villous basement membrane was in contact with Ln α1, β1, γ1, α2, and β2 chains found in the basement membrane. The first three to five cell layers were completely devoid of Ln and Fn chain immunoreactivity. Distinct nuclear reactivity for MAb Ki-67 identified these cells as a proliferating cell population (the proliferative zone). The cells located more distally in the columns were surrounded by distinct immunoreactivity for Ln α1, β1, and γ1 chains and for EDA-, EDB-, and Onc-Fns (matrix deposition zone).
In addition to the sites at which villi were anchored to the basal plate or placental septa, trophoblastic cell columns were also found at sites where a villus was located immediately adjacent to a cell island. In all these locations, the matrix molecule gradient described above was likewise found, and the most proximal cell rows of the columns contained Ki-67 antigen-positive nuclei as a sign of active cell proliferation. It has been proposed that cell islands and columns may be homologous structures (Castellucci et al. 1991; Benirschke and Kaufmann 1990). Our results showing a similar distribution of Ln and Fn chains, as well as localization of the proliferative zone of trophoblastic cells within the two structures, support this hypothesis and suggest that islands indeed may be formed via trophoblastic cell column formation.
Changes in the Extracellular Matrix Characterize ETC Differentiation
Our results suggest that all ETCs, found in diverse locations in the placenta such as in cell islands, cell columns of anchoring villi, the trophoblastic shell, as individual trophoblastic cells among the decidual cells within the basal plate, and as intra-arterial trophoblasts, are frequently associated with immunoreactivity for the Ln α1, β1, and γ1 chains, whereas the α2 and β2 chains are not found. This is interesting in light of the finding that all of these chains are present in the villous trophoblastic BM. It would be interesting to determine whether the Ln α2 and β2 chains are actually expressed in situ by villous trophoblastic cells and downregulated on ETC differentiation. Alternatively, these chains could be deposited into the villous BM solely by the villous stromal cells, as has been suggested for some BMs (Simon-Assmann et al. 1994; Vuolteenaho et al. 1994). In situ hybridization experiments have established that indeed both the villous trophoblast and stromal cells express β1 Ln chain mRNA (Autio-Harmainen et al. 1991). The small amounts of Ln α2 and β2 chains that were sporadically found associated with ETCs found within the decidua could represent Lns that have been secreted by the neighboring decidual cells and assembled at the ETC surface. In partial contrast to our results, Leivo et al. (1989) detected cytoplasmic immunoreactivity for the Ln α2 chain in extravillous trophoblasts within the basal plate, using pepsin-digested paraffin sections. It should be emphasized that they reported cytoplasmic and not pericellular BM-like immunoreactivity for the Ln α2 chain. Our results, however, are supported by those of Damsky et al. (1992,1994), who did not detect the α2 chain in cell columns.
The distribution of Fn isoforms in ETCs is worth noting. Fn per se is found in the villous trophoblastic BM in the early developing placenta but disappears during the late first trimester (Virtanen et al 1988; Yamada et al 1987; Yamaguchi et al., 1985). The abundant immunoreactivity among ETCs in cell islands and columns suggests that the expression of the EDA- and EDB-Fns and the Onc-Fn is another characteristic of ETC differentiation.
Role of the Extracellular Matrix in ETC Invasion
Antibody inhibition experiments have suggested that interaction of α1β1 integrin with Ln and collagen Type IV enhances trophoblast invasion and, conversely, that binding of α5β1 integrin to Fn restrains invasion (Damsky et al. 1994). There are few Ln polypeptides in the proliferative parts of the anchoring cytotrophoblastic cell columns, whereas the distal parts (matrix deposition zone) contain Lns that may be utilized by the invading ETCs for adhesion and for subsequent induction to secrete ECM-degrading proteases (Yudoh et al. 1995; Turpeenniemi-Hujanen et al. 1986). Relatively small amounts of pericellular Lns and Fns are associated with ETCs that are deeper within the placental bed. It is possible that these cells deposit only small amounts of their own matrix and interact largely with the decidual matrix during invasion. It can also be hypothesized that the increased deposition of Fn isoforms in the decidual cell pericellular matrix in response to invasion by ETCs is a mechanism by which the invasion process may be controlled.
In conclusion, the differentiation of extravillous from villous trophoblast is accompanied by distinct modulation in the distribution of Ln and Fn isoforms. ETCs are characteristically associated with Ln α1, β1, and γ1, but not α2 and β2 Ln chains. Furthermore, EDA-, EDB-, and Onc-Fns are abundantly found in the pericellular matrix of differentiating ETCs. Our results also imply that EDB- and Onc-Fns are deposited in decidual cell BMs in response to ETC invasion, and that this modification of the decidual cell basement membrane is distinct from that seen during decidualization.
Footnotes
Acknowledgements
The skillful technical assistance of Ms Marja-Leena Pi-ironen, Mr Reijo Karppinen, and Mr Hannu Kamppinen is gratefully acknowledged.