
Abstract
The peroxidase-mediated deposition of hapten- and fluorochrome-labeled tyramides has recently been shown to increase the sensitivity of immunofluoresence and fluorescence in situ hybridization techniques. We have evaluated a number of red, green, and blue fluorescent tyramides for detection of antigens in tissue sections and cytospin preparations and for the detection of hapten- and horseradish peroxidase-labeled probes hybridized in situ to cells and chromosomes. With few exceptions, all fluorescent tyramide-based methods provided a considerable increase in sensitivity compared to conventional immunofluorescence and FISH methods.
Keywords
FOR LIGHT microscopic visualization of antigens and nucleic acids by immunohisto- and cytochemistry and in situ hybridization, both fluorescence and enzymatic procedures are commonly employed. Enzymatic procedures provide high sensitivity because many substrate molecules are converted to microscopically detectable final reaction products.
Fluorescence-based procedures derive their sensitivity from the fundamentally higher detection sensitivity of fluorescence compared to absorbance. Obviously, a combination of fluorescence- and enzyme-based procedures will profit from both sensitivity features. Most cytochemical enzyme staining procedures are chromogenic, resulting in a precipitated dye. Only a few alkaline phosphatase-based fluorescent substrates have been described (Speel et al. 1992,1995; Kagiyama et al. 1992; Raap 1986).
Horseradish peroxidase (HRP) is another frequently used enzyme, and a variety of chromogenic substrates are available (De Jong et al. 1985). To the best of our knowledge, only one fluorescent enzyme cytochemical reaction has been described for peroxidases. It is based on rhodamine co-precipitation of the oxidation product of homovanillic acid by lead ions and appears to be of historical interest only (Papadimitriou et al. 1976).
With the recent introduction of tyramide conjugates as substrates for HRP, new and potentially very sensitive combinations of enzyme- and fluorescence based cytochemical detection can be pursued. HRP reacts with hydrogen peroxide and the phenolic part of tyramide to produce a quinone-like structure bearing a radical on the C2 group. This “activated” tyramide then covalently binds to tyrosine in close vicinity to the HRP (Van Gijlswijk et al. 1996b; Bobrow and Litt 1993; Bobrow et al. 1989,1992). Therefore, when the tyramide is conjugated to a hapten or fluorochrome, indirect and direct fluorescence detection of enzymatically deposited tyramides is possible.
Although biotin- or hapten-labeled tyramides are valuable reagents in immunocytochemistry (ICC) immunohistochemistry (IHC), and fluorescence in situ hybridization (FISH), directly fluorochrome-labeled tyramides are of special interest because an additional immunological step, which potentially induces background, can be eliminated from the procedure.
In this study we have used green, red, and blue fluorescent tyramides and tested their performance in fluorescence IHC, ICC, and FISH applications. These applications include IHC staining of vimentin in acetone-fixed rat liver cryostat sections, ICC staining of different CD markers on formaldehyde-fixed human mononuclear white blood cells, and FISH for human metaphase chromosome preparations with hapten-labeled probes and 5'-HRP-labeled oligonucleotides.
Materials and Methods
Tyramide Conjugates
Table 1 shows the tyramide conjugates used in this study grouped as red, green, and blue fluorescent tyramides (generous gifts of NEN Life Science Products; Boston, MA). The stock solutions were stored in DMSO at −20C and small aliquots in use at 4C. Tyramides were stored for at least one year without loss of performance.
Immunohistochemistry
Detection of vimentin in 4-μm rat liver cryostat sections from rats, injected with polymeric rat IgA complexes (Bogers et al. 1990), was performed as follows. After acetone fixation for 10 min at room temperature (RT), sections were incubated for 1 hr at 37C with mouse IgG anti-vimentin (Amersham RPN 1102; Poole, UK) diluted 1:40 in TNB [50 mM Tris-HCl, pH 7.4, 150 mM NaCl, 0.5% blocking reagent (NEN Life Science Products)], washed three times for 5 min with TNT [50 mM Tris-HCl, pH 7.4, 150 mM NaCl, 0.05% (v/v) Tween-20], and incubated for 30 min at 37C with HRP-labeled anti-mouse antibody-avidin complexes (Van Gijlswijk et al. 1996a) at 5 μg/ml in TNB. After three 5-min washes in TNT, sections were rinsed in 0.15 M NaCl. Next, 0.5 ml tyramide solution [1 X TSA-buffer (NEN Life Science Products) with the tyramide in the appropriate concentration; see Table 1] was applied and incubated for 30 min at RT. Sections were then washed three times for 5 min in TNT, dehydrated through an ethanol series, and mounted in Vectashield (Vector; Burlingame, CA).
Summary of performance of different fluorescent tyramides in immunohisto/cytochemical and FISH applications a
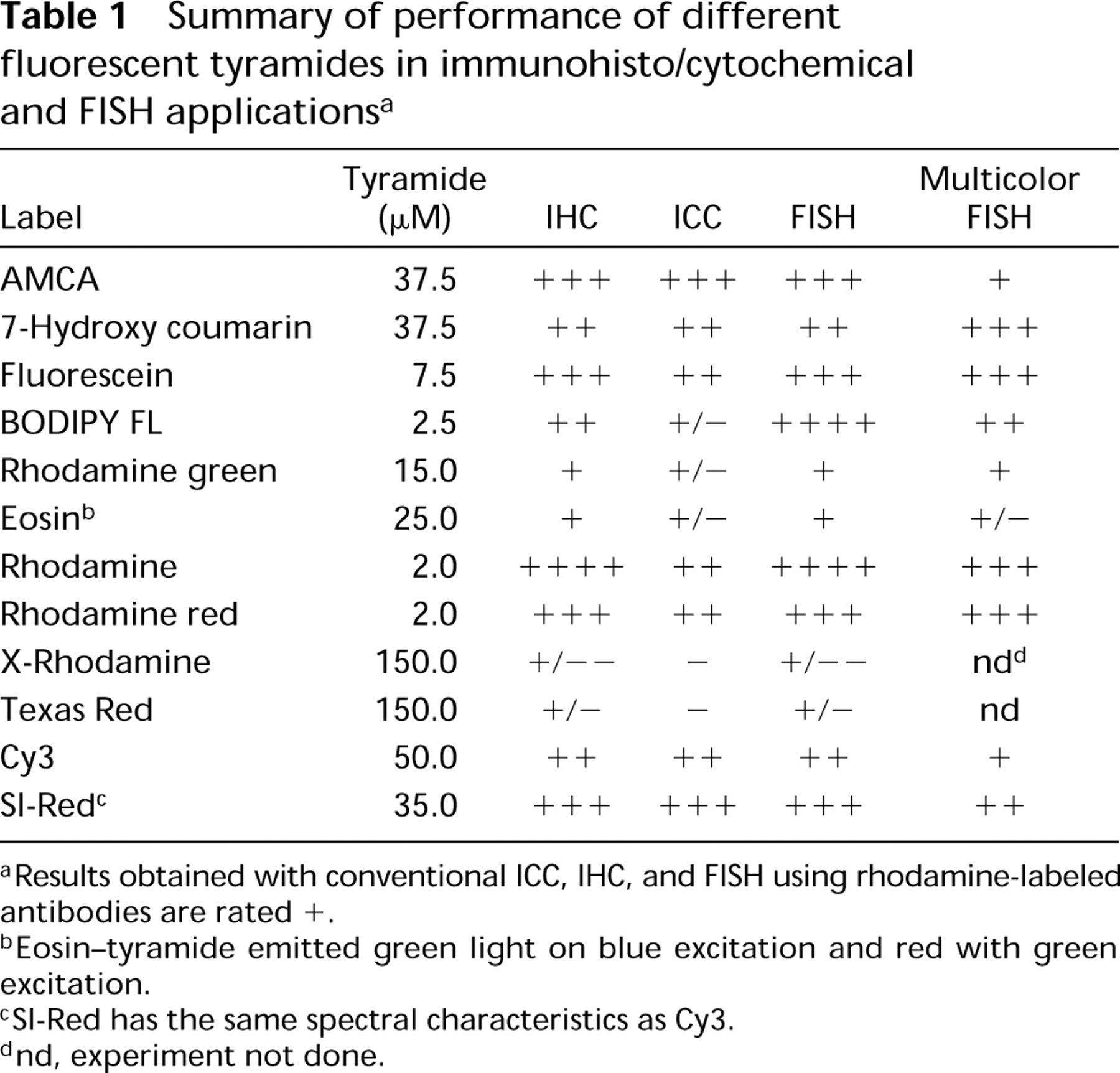
aResults obtained with conventional ICC, IHC, and FISH using rhodamine-labeled antibodies are rated +.
bEosin-tyramide emitted green light on blue excitation and red with green excitation.
cSI-Red has the same spectral characteristics as Cy3.
dnd, experiment not done.
The HRP-labeled goat anti-mouse IgG-avidin D complexes used to detect monoclonal antibodies were synthesized as described (Van Gijlswijk et al. 1996a). For immunohistochemical applications, an IgG-avidin D complex consisting of 1 IgG and 2–3 avidin D molecules proved to be optimal. The appropriate amount of biotin-HRP was added to the complex, mixed, and incubated for 15 min at RT. The complex was then diluted to 2 μg/ml (concentration of IgG) in TNB and incubated. HRP-labeled IgG-avidin D complexes were found to be more sensitive than directly HRP-labeled antibodies.
Immunocytochemistry
Different CD epitopes were detected on 1% formaldehyde-fixed, Ficoll-Isopaque-isolated human mononuclear white blood cells as follows. Mouse IgG monoclonal antibodies against CD4, −9, −13, −14, and −19 (Dakopatts; Glostrup, Denmark, and Becton Dickinson; Mountain View, CA) were incubated on cytospin preparations of the cells. Preparations were subsequently incubated with HRP-labeled goat antimouse IgG-avidin D complexes. For immunocytochemical applications, an IgG-avidin D complex consisting of 2 IgG and 5–6 avidin D molecules proved to be optimal. The appropriate amount of biotin-HRP was added to the complex and incubated for 15 min at RT. The complex was then diluted to 5 μg/ml (concentration of IgG) in TNB and incubated on the preparations. Between incubations they were washed three times for 5 min in TNT. Tyramide detection was performed as described above.
Conventional Immunohisto/Cytochemical Controls
After the monoclonal antibody incubations on tissue section and lymphocyte preparations described above, they were incubated with rhodamine-labeled rabbit IgG-anti-mouse IgG (Sigma, 2 μg/ml; St Louis, MO) and subsequentlly rhodamine-labeled goat anti-rabbit IgG (Sigma; 2 μg/ml). Incubations and washes were with TNB and TNT, respectively.
Fluorescent Tyramide-based FISH with Hapten-labeled Probes
Human metaphase chromosome preparations were used as a target. Methanol/acetic acid (3:1 v/v)-fixed lymphocyte preparations were dropped onto slides, dried for 2 min, dipped in 70% acetic acid, and dried overnight. They were then treated for 15 min at 37C with 0.1 mg/ml RNAse A (Boehringer; Mannheim, Germany) in 2 X SSC and washed twice for 5 min with PBS and once for 5 min at 37C with 0.01 M HCl. Next, they were incubated for 10 min at 37C in 0.05% (w/v) pepsin (Sigma) in 0.01 M HCl, washed twice in PBS, and postfixed for 10 min at RT in 1% formaldehyde/PBS. After two washes in PBS, preparations were dehydrated in ethanol and air-dried.
Both low-complexity probes (plasmids for alphoid DNA and rDNA repeat targets) and high-complexity probes (cosmids, chromosome libraries) were used. Plasmid probes specific for the 28S rRNA genes were denatured and hybridized in a mixture containing 50% formamide, 2 X SSPE (0.33 M NaCl, 20 mM Na-phosphate, pH 7.2, 4 mM EDTA), 10% dextran sulfate, 1 ng/μl probe, and 50 ng/μl herring sperm DNA. Chromosome libraries were denatured and hybridized in the same mixture containing 1 ng/μl labeled probe and 50 ng/μl human Cot-1 DNA. Hybridization of cosmid probes was similar to the libraries, but probe concentrations were 2 ng/μl and the mixture contained 200 ng/μl human Cot-1 DNA. Probe and target were denatured simultaneously by incubating the slides for 2 min at 80C, and hybridization was done for 16 hr at 37C. The slides were subsequently washed once at RT and twice for 5 min at 60C in 2 X SSPE/0.1% Tween-20, followed by two 5-min washes in 0.1 X SSPE at 60C.
Biotin- and digoxigenin-labeled probes were immunocytochemically detected with streptavidin-HRP (NEN Life Science Products; 5 μg/ ml in TNB) and sheep anti-digoxigenin-HRP (Boehringer; 0.5 U/ml in TNB), respectively. Incubations were for 30 min at 37C and washes three times for 5 min in TNT at RT. Tyramide detection was performed as described for tissue sections, except that 20% (w/v) dextran sulfate was added to the detection buffer. The dextran sulfate concentration was lowered to 5% when libraries were used.
Fluorescent Tyramide-based FISH with 5'-HRP-labeled Oligonucleotides
The 5'-HRP oligonucleotides were either generous gifts from NEN Life Science Products or were prepared according to Van Gijlswijk et al. (1997). Briefly, 5'-amino modified oligonucleotides (Eurogentec; Seraing, Belgium) were dissolved in double distilled water (ddH2O) to a final concentration of 2.5 mg/ml. One fifth volume of NHS buffer (0.25 M Na-phosphate, pH 8.0, 0.5 M NaCl, 25 mM EDTA) and one fourth volume of 5 mg/ml succinimidyl 4-[N-maleimidomethyl]cyclohexane-1-carboxylate (SMCC; Pierce; Rockford, IL) in DMSO were added and incubated for 1 hr at 30C. Unreacted SMCC was removed by gel filtration over Sephadex G25 Superfine (Pharmacia; Uppsala, Sweden) using 1 X TE (10 mM Tris-HCl pH 8.0, 1 mM EDTA) as eluent. After ethanol precipitation, the modified oligonucleotides were dissolved in 1 X MEI buffer (50 mM Na-phosphate, pH 6.8, 0.1 M NaCl, 5 mM EDTA) to obtain a 2.5 mg/ml solution. Thiolated HRP was then used to attach the HRP to the maleimide-activated oligonucleotide. To each mg of oligonucleotide, 5 mg of deprotected HRP-SATA (Van Gijlswijk et al. 1996a) was added. After 2 hr at 30 C, the HRP-labeled oligonucleotides were purified using a gradient from 0.2 to 0.7 M NaCl in 1 X TE on a MonoQ 6/1.5 column (Pharmacia).
HRP-labeled oligonucleotides specific for human α-satellite sequences on the centromeres of chromosomes 8, 9, and X, or specific for human satellite III DNA on chromosomes 1 (1q12), or Y (Yq12) were used (see also Van Gijlswijk et al. 1997). In addition, a 40-mer oligonucleotide specific for the 28S rRNA genes was used. Pretreatment of the human metaphase preparations was performed as described above. They were denatured either for 2 min at 80C in 70% formamide, 2 X SSC or for 5 min at 60C in 60% formamide, 1 M betaine-HCl (Janssen Chimica; Olen, Belgium). The preparations were then ethanol-dehydrated and air-dried. Hybridization was performed for 15 min in 3.5 M urea, 50 mM Tris-HCl, pH 7.4, 10 mM EDTA, 150 mM NaCl, 0.05% polyvinylpyrollidone, 0.05% Ficoll 400, 0.05% BSA, 1 mg/ml fish sperm DNA (Boehringer), and 0.1-2 nM probe. For the HRP-oligonucleotide probes specific for chromosomes 1, X, and Y, hybridization was performed at 37C, otherwise hybridization was done at 50C. After hybridization, preparations were washed four times for 3 min with 3 M urea, 50 mM Tris-HCl, pH 7.4, 5 mM EDTA, 150 mM NaCl, and 0.1% Tween-20 at either 37 C (chromosomes 1, X, and Y) or 50C (chromosomes 8 and 9). Optionally, the preparations were washed for 3 min with 40% formamide/1 X SSC at 37C to remove unspecific probe hybridization. After a 5-min wash in PBS at RT, the slides were incubated for 30 min at RT with 1.0 ml of the desired tyramide reaction mixture, washed three times for 3 min in TNT, dehydrated, and air-dried. The preparations were then either embedded in Vectashield or a second 15-min hybridization with a different HRP-labeled oligonucleotide probe was performed. These were then either washed and detected as described or a third hybridization/detection cycle followed. For triple hybridizations, the optimal sequence of fluorescent tyramides was (a) 7-hydroxy coumarin-tyramide, (b) fluorescein-tyramide, and (c) rhodamine- or rhodamine red-tyramide.
Photomicrographs were taken with a DM epifluorescence microscope (Leitz) on 640 ISO (3M) color slide film. A triple filter (Chroma Technology Corp. 61000-Triple: DAPI/FITC/TRITC excitation 385–405 nm, 480–500 nm, 550–580 nm; emission 455–475 nm, 510–525 nm, 590–620 nm) was used to visualize multicolored FISH results.
Results
Tyramide-based Immunohisto- and Cytochemistry
General. With the exception of Texas Red and X-rhodamine, all fluorescent tyramides tested improved the sensitivity of immunofluorescent assays compared to conventional detection with fluorescent antibodies (see Table 1). As an example, Figure 1 shows the results of the detection of vimentin on rat liver tissue using rhodamine-labeled antibodies (Figure 1 A) and rhodamine-tyramide (Figure 1 B). Specific detection of vimentin in Kupffer cells and endothelial cells of blood vessels and the bile duct was obtained. No effort was made to block endogeneous peroxidase on these tissue sections. Controls in which the primary monoclonal antibody was omitted showed faint staining of the Kupffer cells.
Sensitive results were also obtained when formaldehyde-fixed white blood cells were stained for CD markers with fluorescent tyramides (Table 1; Figure 3). The percentage of positive cells with the different monoclonal antibodies using fluorescent tyramide detection was equal to the percentages obtained with fluorochrome-labeled antibodies.
Red Tyramides. With the exception of Texas Red- and X-rhodamine, red tyramides performed well in the ICC and IHC assays, with a slight preference for rhodamine and rhodamine red-tyramide. Figure 2 shows that SI-Red-tyramide performed better than Cy3-tyramide (see also Chao et al. 1996). X-rhodamine- and Texas Red-labeled tyramides were inferior. The intensity of the fluorescence signals was generally less compared with those obtained using TRITC-labeled secondary antibodies.
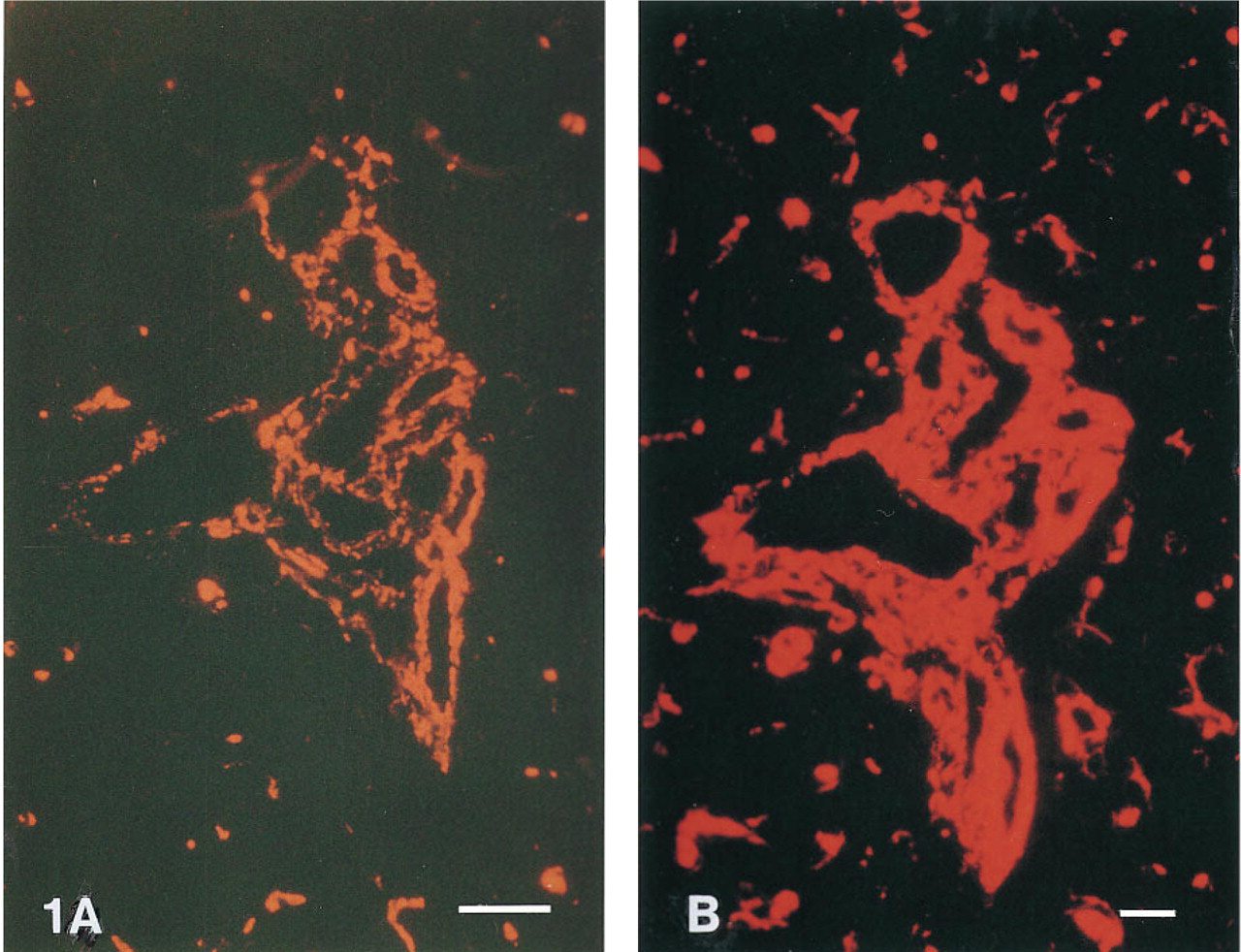
Immunohistochemical staining of vimentin in rat liver tissue. (
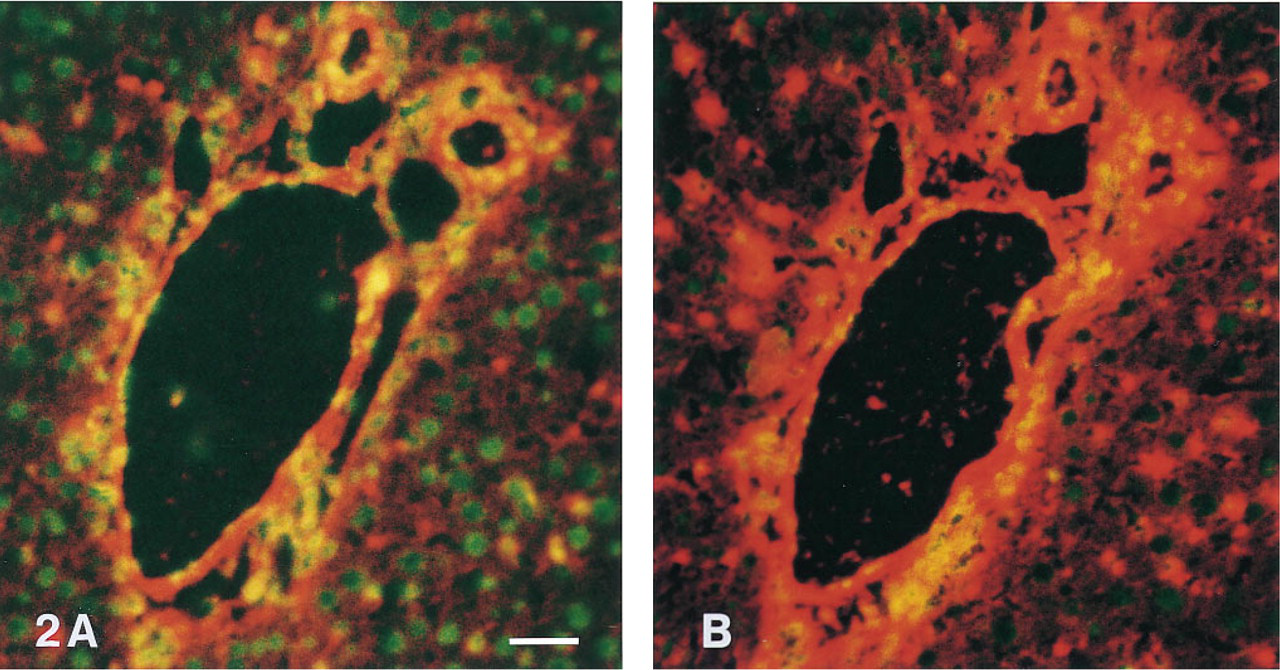
Tyramide-based immunohistochemical staining of vimentin on rat liver tissue using HRP-labeled antibody-avidin complexes and Cy3- and SI-Red-tyramides. Nuclei were counterstained in green with YOYO-1. Microphotography was through a double bandpass filter. With Cy3 tyramide (
Blue Tyramides. Both blue (AMCA and 7-hydroxy coumarin) tyramides gave good results. Care should be taken to select the optimal filter set because the spectra differ. For example, on using triple bandpass filters, 7-hydroxy coumarin-tyramide was found superior, whereas with the single bandpass filter AMCA-tyramide provided the best sensitivity. The coumarin-labeled tyramides can be used at high concentrations with little or no background staining.
Except for the two coumarin-, SI-Red-, and Cy3-tyramides, some background staining on the membranes of formaldehyde-fixed human white blood cells can occur with the fluorescent tyramides tested. However, even though the negative cells stain to some extent, specific staining is still easily observed. Background staining can be reduced by using more dilute tyramide solutions in combination with extended incubation times.
Green Tyramides. All green fluorescent tyramides tested (fluorescein, rhodamine green, BODIPY FL, and eosin tyramides) proved good substrates for HRP. However, fluorescein-tyramide is preferred because rhodamine green, eosine, and BODIPY FL tyramides may cause background problems on cells and tissue sections. In contrast to ICC and IHC, tyramide-related background problems are considerably less with chromosomal FISH, and both fluorescein- and BODIPY FL-tyramide gave good results. When BODIPY FL-tyramide was used at high concentrations, the BODIPY FL fluorescence emission shifted to the red (Figure 4 B). Eosin-tyramide performed well in FISH. Deposited eosin-tyramide emitted green light at blue excitation (Figure 2 B) and red light at green excitation.
Tyramide-based FISH
As reported earlier, tyramide-based FISH with hapten-labeled, cloned DNA probes is highly sensitive (De Haas et al. 1996; Van Gijlswijk et al. 1996b; Kerstens et al. 1995). The results shown in Table 1 were obtained with middle repetitive specific probes (28S rRNA genes) and single-copy gene-specific cosmid probes (target size 30–40
Figure 5 shows the result obtained with a chromosome library probe specific for chromosome 11. Library probe concentrations were decreased tenfold and the excess of human Cot 1 DNA was raised fivefold compared to the optimal protocol for conventional chromosome painting.
Figure 6 shows that fluorescent tyramides can be used to perform multicolor FISH staining. The triple FISH experiment with 5'-HRP-labeled oligonucleotides specific for (peri)centromeric regions of the human chromosomes 1, 8, and 9 illustrates that multiple targets can be visualized using sequential rounds of hybridization/tyramide detection. We found the sequence 7-hydroxy coumarin-, fluorescein-, and then rhodamine-tyramide optimal. Using this sequence with tyramide concentration as given in Table 1, intermittent HRP inactivation proved not to be necessary.
Multicolor tyramide FISH experiments were also conducted using digoxigenin- and biotin-labeled centromeric plasmid probes. These were visualized by sequential rounds of HRP-conjugate incubation/fluorescent tyramide staining (data not shown). Intermittent HRP inactivation (using PBS/1% hydrogen peroxide) now proved necessary.
We noted that bicolor tyramide FISH for co-localizing targets should be designed with care. For example, for combined cosmid and chromosome library FISH, the cosmid signal was not visible when the library was developed first. Apparently the tyramides immobilized in the first detection round occupied most if not all tyramide binding sites.
Discussion
A number of fluorescence-labeled tyramides were tested for their performance in immunohisto- and cytochemical and FISH applications.
With the exception of X-rhodamine- and Texas Red-tyramides, all tyramides were useful substrates. Chemically, X-rhodamine and rhodamine differ from the others in that X-rhodamine and Texas Red contain an additional sulfonic acid group. This adds negative charge to the labeled tyramide. Possibly, the different HRP-generated tyramide radicals differ in reactivity, dependent on the conjugated fluorescent group. This would alter the ability of the radical to bind to adjacent tyrosine moieties.
As with biotinylated tyramide (Adams 1992), the use of fluorescent tyramides can significantly improve the detection of antigens (Figures 1 and 3). Most fluorescent tyramides proved to be good substrates for HRP in the IHC/ICC applications tested. The two blue fluorescent coumarin-, SI-Red-, and Cy3-tyramides all gave strong signals and low background staining. However, rhodamine- and fluorescein-tyramide performed very well, too. The sensitivity increase obtained with fluorescent tyramides was not quantified. However, previous investigations indicated that it could be 10–100 times (Chao et al. 1996; Kerstens et al. 1995; Raap et al. 1995; Adams 1992).
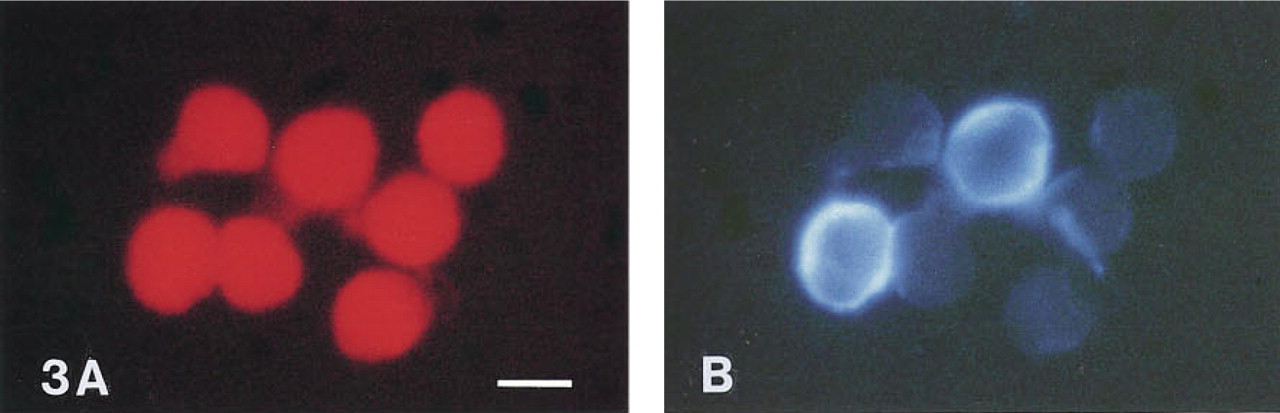
Immunocytochemical detection of CD14 epitopes on human mononuclear white blood cells with AMCA tyramide (
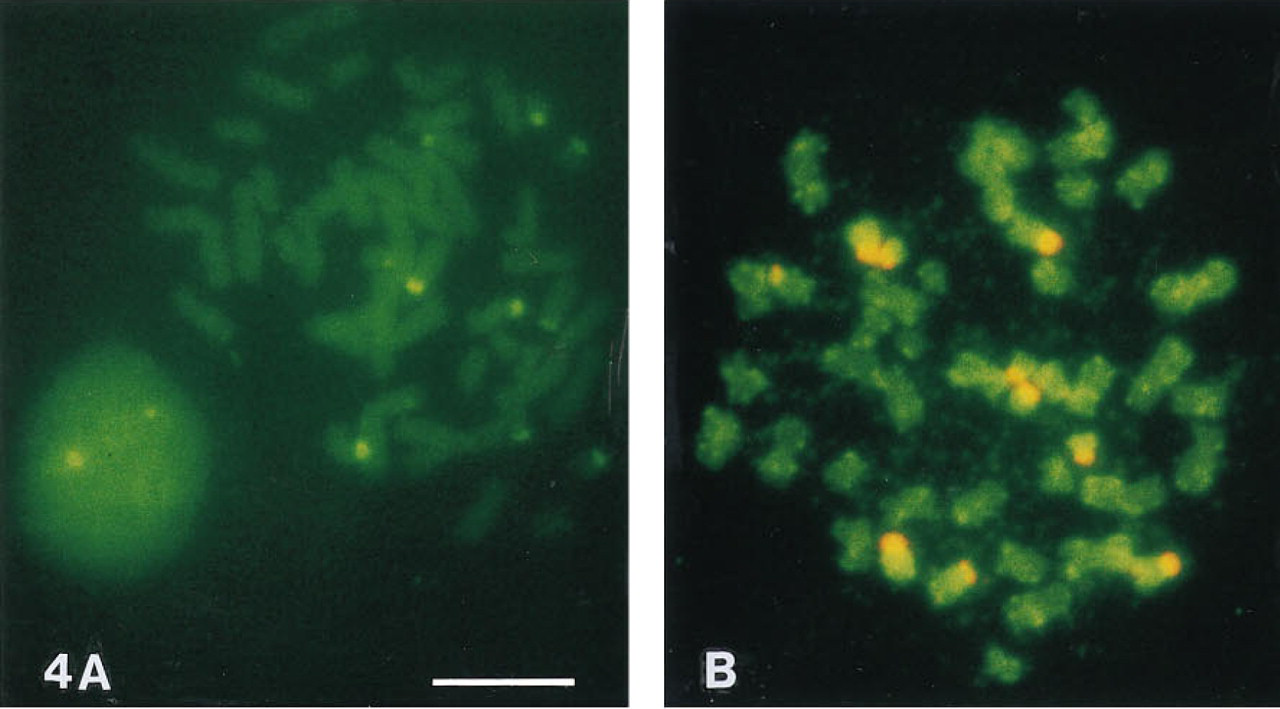
Tyramide-based FISH detection of the 28S ribosomal RNA genes in human metaphase chromosomes using eosin-tyramide (
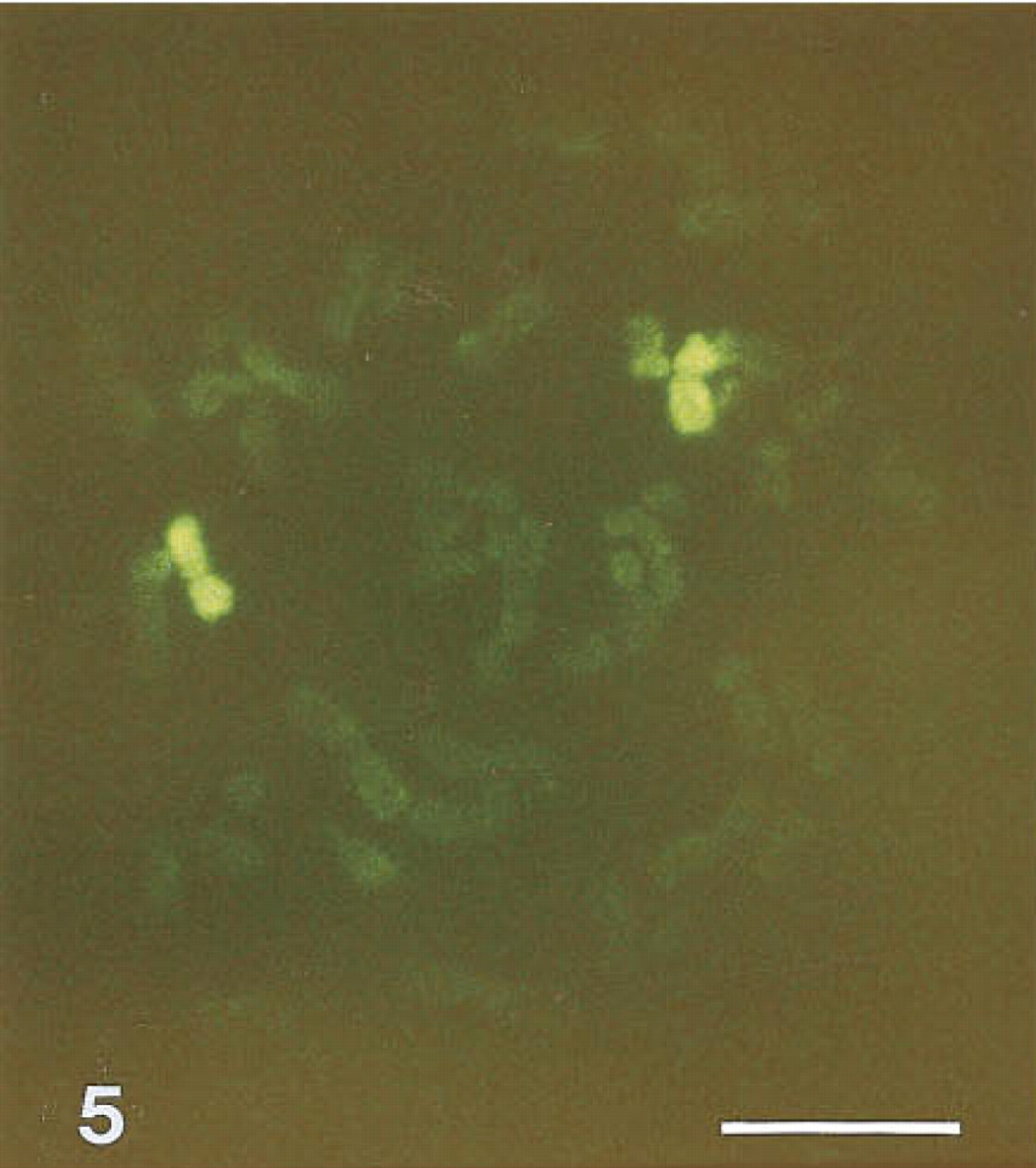
Fluorescein-tyramide-based FISH of human metaphase chromosomes with a chromosome 11 library. Bar = 5 μm.
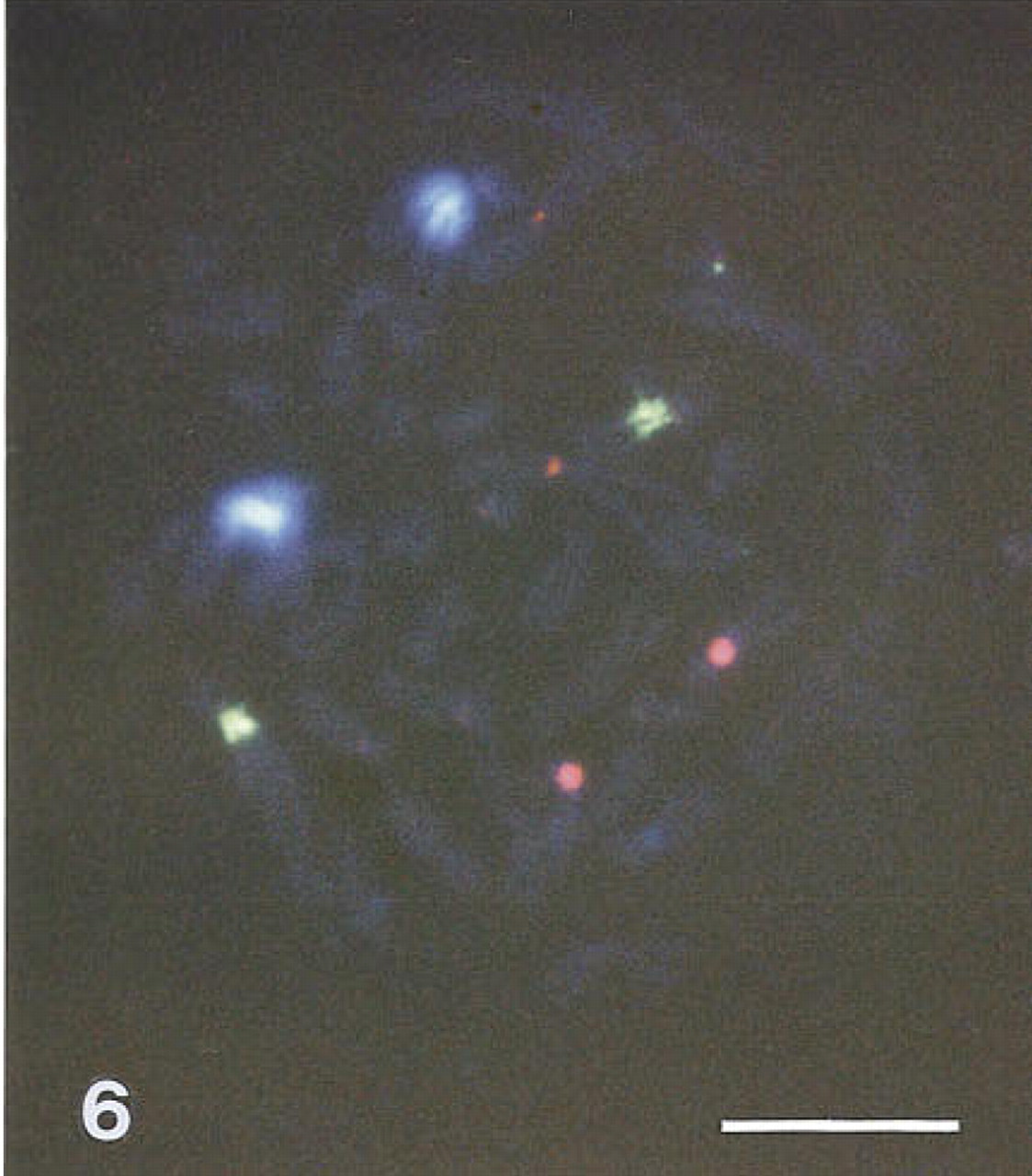
Tyramide-based triple FISH of human metaphase chromosomes with 5'-HRP-labeled oligonucleotides specific for the (peri)centomeric regions of chromosome 1 developed to 7-hydroxy coumarin-tyramide, chromosome 9 developed to fluorescein-tyramide, and chromosome 8 developed to rhodamine Red tyramide. Exposure was 90 sec through a triple bandpass filter. Bar = 5 μm.
The high intensity of the fluorescent signals obtainable with tyramide overcomes, at least in part, the problem of autofluorescence of the specimen. De Haas et al. (1996) showed that by using biotinylated tyramide and a time-resolved marker, autofluorescence can be further decreased.
As with ICC and IHC, in FISH a high signal intensity was obtained with most tyramides tested (Table 1). When FISH with plasmid or cosmid probes was performed, dextran sulfate was added to the tyramide detection buffer to improve the localization of the end product (Van Gijlswijk et al. 1996b). For some experiments, such as with chromosome painting, dextran sulfate concentration was lowered because better chromosomal paints were obtained. It should be realized, however, that part of the resulting paint coverage can in fact be artificial when the fluorescent tyramides are poorly localized. For detection of translocations involving small parts of chromosomes, the use of PVA or high dextran sulfate concentrations is recommended as otherwise the smaller translocation partner may become painted by the larger.
An advantage of the use of tyramides for FISH with high complexity probes such as whole chromosome libraries is that lower probe concentrations can be used. When the Cot-1 DNA concentration is maintained, the resulting higher ratio of supression/probe DNA leads to a more homogeneous staining of the target chromosome, and hybridization signals to other parts of the genome are suppressed better than in conventional techniques. The low probe concentration also allows more tests per labeled probe batch.
Multicolored tyramide FISH is quite feasible, but because tyramide detection is strictly peroxidase-dependent, it relies on sequential rounds of incubations with HRP-labeled oligonucleotides or, in the case of hapten-labeled probes, HRP-labeled antibodies and a tyramide staining reaction. This method is useful for non-colocalizing targets, but when the different targets co-localize one should be aware that the first round of tyramide detection may prevent deposition of tyramides in the second. In this respect, combined conventional immunofluorescence and tyramide-based detection is an obvious option.
Interestingly, BODIPY FL-tyramide gave red fluorescent signals on blue excitation, especially when relatively high intensities were present (Figure 4 B). When similar experiments were performed with relatively high concentrations of fluorescein- and rhodamine-tyramide, complete quenching of the FISH signals was observed. This clearly indicates that large amounts of tyramides can be deposited on or close to the target.
The high signal generation capacity of the HRP- fluorescent tyramide reaction allowed the use of direct HRP-labeled oligonucleotides for rapid chromosome detection and enumeration (Figure 6; see also Van Gijlswijk et al. 1997). An advantage of this approach is that vast amounts of probe can be produced merely in a chemical way, eliminating repeated plasmid labeling and quality controls to a great extent. Furthermore, because the complexity of the probe is lessened it allows discrimination of targets with little difference in base composition.
In conclusion, the detection of antigens and DNA sequences with fluorescent tyramides using immunoaffinity or hybridization methods is highly sensitive compared to conventional methods. Many different fluorochromes can be conjugated to tyramines, and most of the resulting tyramides proved to be good substrates for tyramide signal amplification.
Footnotes
Acknowledgements
We thank Danielle van Gijlswijk-Janssen for providing us with the tissue sections and Leica for the DM microscope.